TRIM44沉默对肝癌细胞增殖的影响及分子机制研究
2016-12-26张蕴蕴吴永忠
张蕴蕴, 田 艺, 周 伟, 孙 卯, 吴永忠
(重庆市肿瘤研究所,重庆 400030)
TRIM44沉默对肝癌细胞增殖的影响及分子机制研究
张蕴蕴, 田 艺, 周 伟, 孙 卯, 吴永忠△
(重庆市肿瘤研究所,重庆 400030)
目的: 研究三结构域蛋白44(tripartite motif-containing protein 44, TRIM44)对肝癌细胞增殖的影响并探讨其分子机制。方法:RT-qPCR和Western blot法分别检测TRIM44在正常肝组织、肝癌组织和癌旁组织,以及永生化肝细胞和肝癌细胞系中的mRNA和蛋白表达水平;在肝癌细胞中转染靶向沉默TRIM44的shRNA,Wes-tern blot实验检测对TRIM44的沉默效果;MTS实验分析TRIM44沉默对肝癌细胞活力的影响,EdU标记实验检测TRIM44沉默对肝癌细胞DNA合成能力的影响;软琼脂集落形成实验检测TRIM44沉默对肝癌细胞锚定非依赖生长能力的影响;Western blot法检测TRIM44沉默对哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)总蛋白及磷酸化水平的影响;用mTOR激活剂MHY1485处理TRIM44沉默的肝癌细胞,并通过MTS实验分析肝癌细胞的活力。结果:TRIM44的mRNA和蛋白水平在肝癌组织中高于癌旁组织和正常肝组织,TRIM44的mRNA和蛋白水平在肝癌细胞系中高于永生化肝细胞;TRIM44沉默可抑制肝癌细胞的活力、DNA合成能力及锚定非依赖生长能力;TRIM44沉默可降低mTOR磷酸化水平,MHY1485拮抗TRIM44沉默对肝癌细胞活力的抑制作用。结论:TRIM44沉默可能通过下调mTOR活性抑制肝癌细胞增殖。
三结构域蛋白44; 肝细胞癌; 哺乳动物雷帕霉素靶蛋白
三结构域蛋白(tripartite motif-containing protein,TRIM)家族是细胞生物学功能的关键调节因子,TRIM家族成员含有多域的E3泛素连接酶,并以N端存在三结构域为特征[7]。研究表明TRIM家族与细胞增殖、分化、凋亡、迁移和初始免疫等有关[7-10]。目前,TRIM蛋白家族成员TRIM16、TRIM3、TRIM28、TRIM40和TRIM26在恶性肿瘤中的作用也逐渐被证实[11]。而TRIM44的功能研究较少,已知TRIM44含有B box、coiled coil和zinc finger 结构域。由于zinc finger结构域在泛素特异性蛋白酶(ubiquitin-specific protease,USP)家族也有发现,因此TRIM44可能发挥“USP样TRIM”功能抑制泛素化过程。据文献报道TRIM44有助于多种病理学状态,如癌症、发育障碍、神经退行性变和病毒感染[12]。也有研究证实了TRIM44在非小细胞肺癌、食管腺癌和胃癌发展中的作用[12-14]。然而,目前尚无TRIM44在HCC发生发展中的作用研究报道。本研究将检测TRIM44在肝癌细胞、永生化肝细胞及正常肝组织中的表达差异,并在肝癌细胞中用shRNA沉默TRIM44的表达,进而观察肝癌细胞增殖的变化及探讨其分子机制,以分析TRIM44对HCC进展的影响,为鉴定HCC药物治疗靶点提供新的依据。
材 料 和 方 法
1 材料

2 方法
2.1 细胞培养 SK-Hep-1、PLC/PRF/5、Huh-7和MIHA细胞均培养于含10% 胎牛血清、1% 青-链酶素的DMEM培养基中;HepG2细胞培养于含10%胎牛血清、1% 青-链酶素的MEM培养基中。所有细胞均在5% CO2、37 ℃培养箱中常规培养。
2.2 质粒转染 接种8×104SK-Hep-1细胞或PLC/PRF/5细胞于6孔板中,24 h后更换为无抗生素的DMEM培养基。将2 μg质粒与6 μL质粒转染试剂混匀后,静置15 min,均匀滴加于6孔板中。24 h后更换为常规培养基。
2.3 RT-qPCR 收集适量细胞,用1 mL TRIczol裂解细胞,提取RNA。取米粒大小正常肝组织、肝癌组织或癌旁组织于流式管中,加入1 mL TRIzol,用组织匀浆机破碎组织,并提取组织RNA。取1 μg RNA逆转录成cDNA,随后使用FastStart Universal SYBR Green Master Mix进行PCR,以β-actin为内参照。TRIM44的上游引物5’-GCGAGGCCGAAGAAGACAAC-3’,下游引物为5’-TCCGGACACTTCCTCTTGGC;β-actin的上游引物为5’-CTCTTCCAGCCTTCCTTCCT,下游引物为5’-AGCACTGTGTTGGCGTACAG-3’。PCR反应条件为 95 ℃ 2 min; 94 ℃ 20 s,60 ℃20 s,72 ℃ 20 s,75 ℃ 1 s,循环34次。每组实验重复3次。以2-ΔΔCt计算目的基因的相对表达水平。
表1为主成分的特征值和方差贡献率。累积方差贡献率是确定需要采用的主成分个数的主要依据,其次才是特征值。表中第一主成分(PC1)的特征值λ1=0.047,第二主成分(PC2)的特征值λ2=0.016,所有主成分的特征值均小于1,均不能用以确定主成分的个数。PC1、PC2的方差贡献率分别为70.259%、23.212%,前2个主成分解释了总方差的近93.47%,故可以选择前2个主成分来取代45个原始变量。
2.4 Western blot实验 收集细胞,加入相应体积含蛋白酶抑制剂的RIPA裂解缓冲液,置于冰中摇床上裂解15 min,16 000×g离心5 min,吸取上清,用BCA法测定蛋白浓度。取米粒大小正常肝组织、肝癌组织或癌旁组织于流式管中,加入1 mL 含蛋白酶抑制剂的RIPA裂解缓冲液,用组织匀浆机破碎组织,破碎后将流式管置于冰中15 min,转移样本至EP管中,16 000×g离心5 min,吸取上清,用BCA法测定蛋白浓度。取30 μg蛋白上样,SDS-PAGE分离蛋白后,转移蛋白至NC膜,5%脱脂牛奶封闭1 h,稀释 I 抗(TRIM抗体、mTOR抗体和p-mTOR抗体均1∶1 000稀释,GAPDH抗体以1∶5 000稀释),4 ℃摇床孵育过夜,II 抗室温下摇床孵育2 h,ECL显影。以GAPDH为内参照。
2.5TRIM44沉默效果检测 shControl和shTRIM44转染细胞72 h后提取细胞总蛋白,测定浓度后,用Western blot法检测样本中的TRIM44蛋白及内参照GAPDH表达水平,并比较shControl和shTRIM44转染后,TRIM44蛋白表达差异。
2.6 MTS实验 细胞转染质粒24 h后,消化细胞计数,取8 000个细胞接种于96孔板。分别在转染后 2 d、3 d、4 d、5 d加入20 μL MTS试剂,5% CO2、37 ℃培养箱中静置2 h,于490 nm处读取样品吸光度。每个样品重复3次。
2.7 EdU标记实验 细胞接种于含无菌盖玻片的6孔板中,24 h后转染质粒,3 d后运用Click-iT EdU Imaging Kits进行EdU标记实验,实验操作严格按试剂盒说明书进行。
2.8 软琼脂集落形成实验 将1.2%的琼脂糖凝胶与含有20%血清的2×DMEM培养基等体积充分混匀,加入6孔板中,作为底层琼脂糖凝胶,室温凝固。细胞转染质粒24 h后,消化细胞并计数,取1 000个细胞加入含有20%血清的2×DMEM培养基中,混匀后再与0.7%的琼脂糖凝胶等体积充分混匀作为上层胶加入铺有底层胶的6孔板中,置于5% CO2、37 ℃培养箱中培养。每3 d向6孔板中加入若干滴培养基,3 周后观察集落形成,0.05%结晶紫染色。
3 统计学处理
采用SPSS 19.0软件进行统计学分析,计量资料用均数±标准差(mean±SD)表示。对两样本均数间比较采用配对t检验。多样本均数间比较采用单因素方差分析,各组均数的两两比较采用SNK-q检验。以P<0.05为差异有统计学意义。
结 果
1 TRIM44在肝癌组织和癌旁组织中的表达差异
RT-qPCR分析结果显示TRIM44的mRNA水平在肝癌组织中显著高于癌旁组织和正常肝组织,见图1;Western blot法检测结果也显示TRIM44的蛋白水平在肝癌组织中也显著高于癌旁组织和正常肝组织,见图2。以上结果说明TRIM44在肝癌组织和肝癌细胞中高表达,提示TRIM44可能有利于肝癌细胞生长。

Figure 1. The mRNA level of TRIM44 in the normal liver tissues (2 cases), HCC (4 cases) and adjacent nontumor liver tissues (4 cases) determined by RT-qPCR. N: adjacent nontumor liver tissue; T: HCC tissue. Mean±SD.**P<0.01vsN.
图1 RT-qPCR 检测正常肝组织、肝癌组织和癌旁组织中TRIM44的mRNA水平
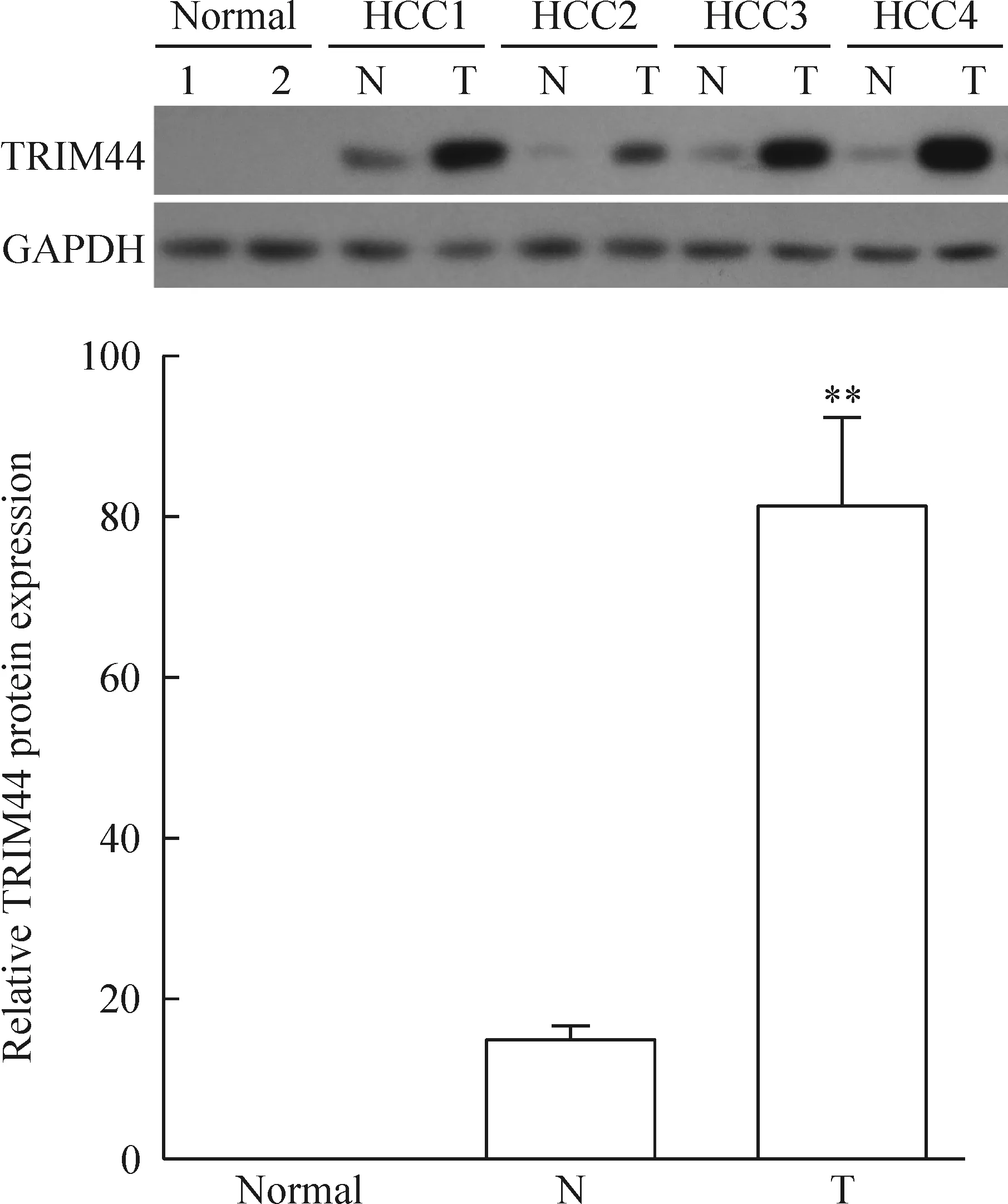
Figure 2. The protein level of TRIM44 in the normal liver tissues (2 cases), HCC (4 cases) and adjacent nontumor liver tissues (4 cases) determined by Western blot. N: adjacent nontumor liver tissue; T: HCC tissue. Mean±SD.**P<0.01vsN.
图2 Western blot法检测正常肝组织、肝癌组织和癌旁组织中TRIM44的蛋白水平
2 TRIM44在不同细胞系中的表达差异
我们通过RT-qPCR分析了TRIM44 的mRNA水平在不同细胞中的表达差异,结果显示TRIM44的mRNA水平在肝癌细胞系中显著高于永生化肝细胞,见图3。另外,我们也用Western blot法检测了TRIM44蛋白在不同细胞中的表达差异,结果显示TRIM44 蛋白水平在肝癌细胞系中也显著高于永生化肝细胞,见图4。

Figure 3.The mRNA level of TRIM44 in immortalized hepatocytes and hepatoma cell lines determined by RT-qPCR. Mean±SD.n=3.*P<0.05vsMIHA.
图3 RT-qPCR 检测永生化肝细胞和肝癌细胞中TRIM44的mRNA水平

Figure 4. The protein level of TRIM44 in immortalized hepatocytes and hepatoma cell lines determined by Western blot. Mean±SD.n=3.*P<0.05vsMIHA.
图4 Western blot法检测永生化肝细胞和肝癌细胞中TRIM44的蛋白水平
3 TRIM44沉默对肝癌细胞活力的影响
我们首先选取了TRIM44表达较高的2种肝癌细胞SK-Hep-1和PLC/PRF/5,并在细胞中转染了靶向沉默TRIM44表达的shRNA质粒shTRIM44及对照质粒shControl,用Western blot法检测沉默效率,结果显示shTRIM44明显沉默了TRIM44在这2种细胞中的表达,见图5。在TRIM44有效沉默的基础上,我们进行了MTS实验,结果发现沉默TRIM44表达明显抑制了SK-Hep-1和PLC/PRF/5细胞的活力,且随着天数的增加,抑制趋势更加明显;在第5 天时,TRIM44沉默对SK-Hep-1和PLC/PRF/5细胞活力的抑制率分别达到50.8%和47.5%,差异具有统计学显著性(P<0.01),见图6。这说明TRIM44沉默可显著抑制肝癌细胞的活力。

Figure 5.The silencing efficiency of shTRIM44 in SK-Hep-1 and PLC/PRF/5 cells analyzed by Western blot. Mean±SD.n=3.*P<0.05vsshControl.
图5 Western blot法检测SK-Hep-1 和 PLC/PRF/5细胞中shTRIM44的沉默效率
4 TRIM44对肝癌细胞DNA合成能力的影响
我们在SK-Hep-1细胞中沉默了TRIM44的表达并进行了EdU标记实验。结果显示TRIM44沉默组的EdU阳性细胞数比对照组减少约46.4%,说明TRIM44沉默可显著抑制肝癌细胞的DNA合成能力,见图7。
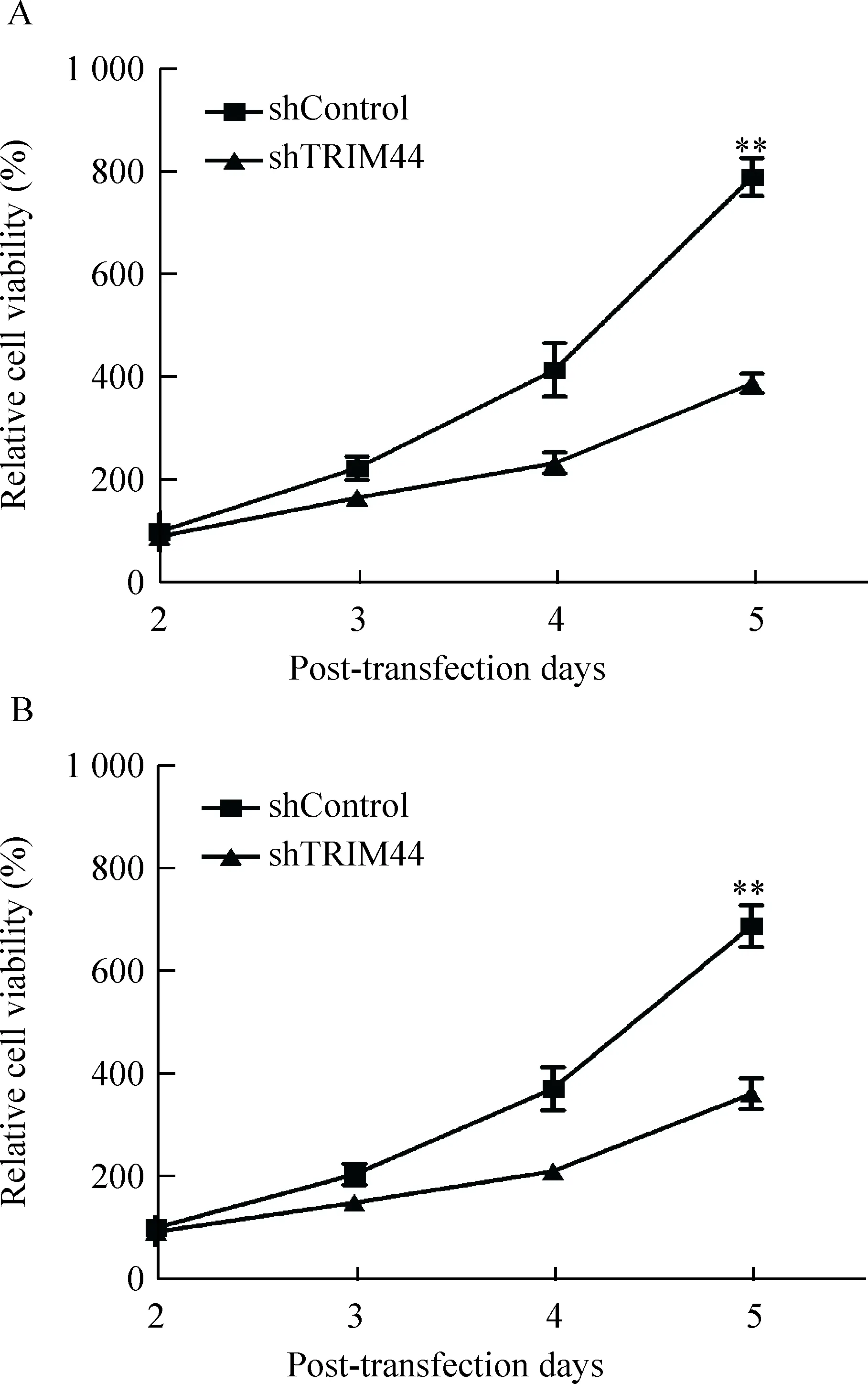
Figure 6. The effects ofTRIM44 knockdown on the viability of SK-Hep-1 (A) and PLC/PRF/5 (B) cells measured by MTS assay. Mean±SD.n=3.**P<0.01vsshControl.
图6 MTS检测TRIM44沉默对SK-Hep-1 和 PLC/PRF/5细胞活力的影响

Figure 7.The effect ofTRIM44 knockdown on the DNA synthesis in SK-Hep-1 cells detected by EdU incorporation assay. Mean±SD.n=3.*P<0.05vsshControl.
图7 EdU标记实验检测TRIM44沉默对SK-Hep-1细胞DNA合成能力的影响
5 TRIM44对肝癌细胞锚定非依赖生长能力的影响
我们在SK-Hep-1细胞中沉默了TRIM44的表达并进行了软琼脂集落形成实验,结果显示TRIM44沉默的SK-Hep-1细胞形成集落的数目比对照细胞减少55%,说明TRIM44沉默可显著抑制肝癌细胞的锚定非依赖生长能力,见图8。以上结果揭示TRIM44沉默明显抑制了肝癌细胞增殖。
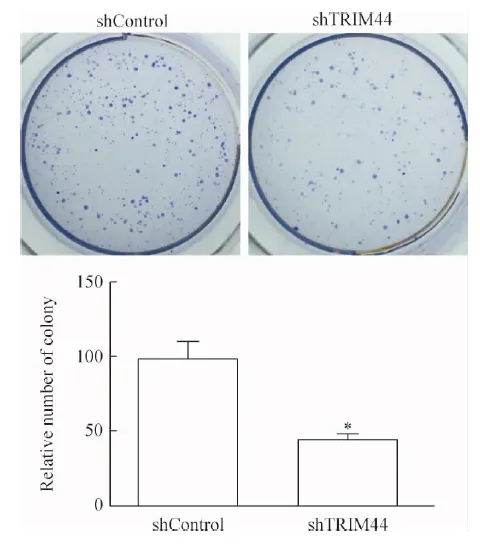
Figure 8.The effect ofTRIM44 knockdown on the ability of anchorage-independent growth in SK-Hep-1 cells observed by the method of colony formation on the soft agar. Mean±SD.n=3.*P<0.05vsshControl.
图8 软琼脂集落形成实验检测TRIM44沉默对SK-Hep-1细胞锚定非依赖生长能力的影响
6 mTOR在TRIM44沉默抑制肝癌细胞增殖中的作用
我们通过Western blot法检测了TRIM44沉默的SK-Hep-1细胞中的mTOR及mTOR活性形式p-mTOR的蛋白水平,结果显示TRIM44对mTOR表达未见显著影响,而能明显抑制p-mTOR的蛋白水平,提示TRIM44沉默降低了mTOR的活性,见图9。为进一步分析mTOR在TRIM44沉默抑制肝癌细胞增殖中的作用,我们在沉默TRIM44的同时用2 μmol/L的mTOR激活剂MHY1485处理细胞,Western blot法检测发现p-mTOR的蛋白水平得到明显恢复,见图10。而与此同时,MTS实验分析结果显示MHY1485也明显拮抗了TRIM44沉默对肝癌细胞活力的抑制作用,见图11。这些发现揭示TRIM44可能通过下调mTOR信号通路进而抑制肝癌细胞增殖。
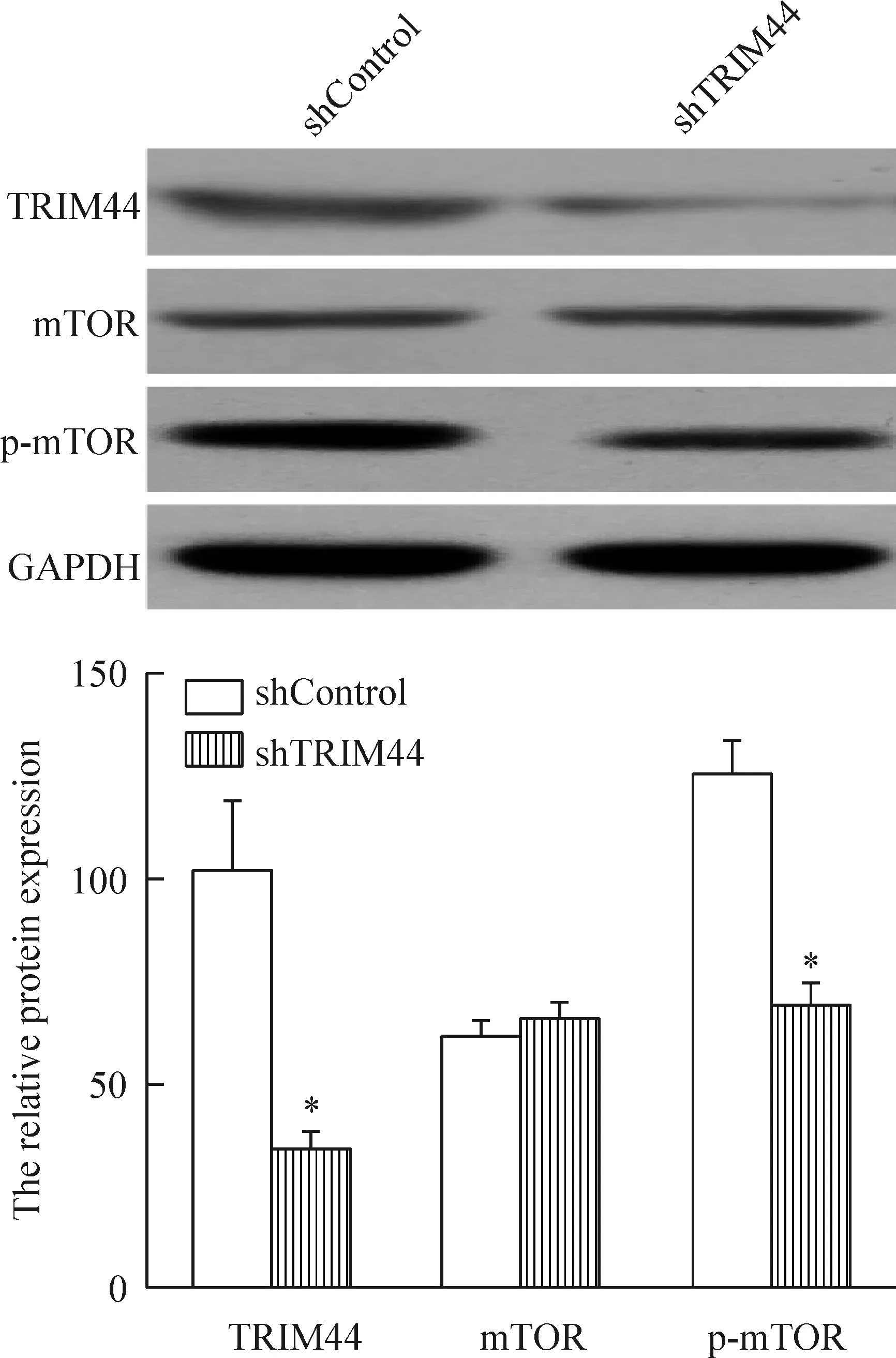
Figure 9.The effect ofTRIM44 knockdown on the protein levels of mTOR and p-mTOR measured by Western blot. Mean±SD.n=3.*P<0.05vsshControl.
图9 Western blot法检测TRIM44沉默对mTOR和p-mTOR蛋白水平的影响
讨 论
近年来,许多文献报道了TRIM44与肿瘤的关系。研究发现TRIM44在胃癌组织中的高表达与淋巴结转移和复发有关,也可作为患者不良预后的预测因子[14],并且TRIM44的表达可增加巴雷特食管病发展为高度异型增生,最后进展为食管癌[13, 15]。在对非小细胞肺癌的研究中也发现TRIM44的高表达与肿瘤的低分化、TMM分期、淋巴结转移、恶性腺瘤亚型和不良预后有关,TRIM44过表达能通过mTOR信号通路促进上皮间质转化过程,进而促进肿瘤的侵袭和转移,并且加快了G1/S期转变,促进了肿瘤细胞增殖[12]。这些研究证实TRIM44是肿瘤形成和进展的一个重要因素。本研究主要关注于TRIM44对肝癌细胞增殖的影响,我们发现TRIM44在肝癌细胞及肝癌组织中是高表达的,TRIM44沉默能抑制肝癌细胞增殖。有文献报道在肿瘤进展中TRIM44与mTOR信号通路密切相关[12, 15],并且mTOR活化有利于肝癌细胞增殖[16]。我们的进一步研究也发现TRIM44沉默可降低mTOR的活性,并且mTOR激活剂可拮抗TRIM44沉默对肝癌细胞增殖的抑制作用,这些发现揭示TRIM44沉默可能通过下调mTOR的活性抑制肝癌细胞生长。
Figure 10.The effect ofTRIM44 knockdown with or without mTOR agonist MHY1485 on the protein levels of p-mTOR determined by Western blot. Mean±SD.n=3.*P<0.05vsshControl;#P<0.05vsshTRIM44.
图10 Western blot法检测mTOR激动剂MHY1485对 p-mTOR蛋白水平的影响

Figure 11.The effect of mTOR agonist MHY1485 on the viability ofTRIM44-silenceing SK-Hep-1 cells analyzed by MTS assay. Mean±SD.n=3.**P<0.01vsshTRIM44.
图11 MTS实验分析MHY1485对TRIM44沉默的SK-Hep-1细胞活力的影响
mTOR是一种高度保守的蛋白激酶,属于磷脂酰肌醇3-激酶相关激酶[phosphoinositide 3-kinase (PI3K)-related kinase,PIKK]家族[17]。mTOR信号通路是细胞多种功能的关键调节因子,如细胞生长、增殖和代谢[18-19]。mTOR下游的靶点有4E-BP1、S6K、SREBP、ATG13、Akt、SGK等,通过这些靶点mTOR可调节蛋白和脂类合成、溶酶体生物合成、自噬、能量代谢、细胞生长、凋亡和血管生成等生理过程。其上游的调节因子有TSC1/2、Rheb、RAG、AMPK等[18]。目前,已发现mTOR信号通路与HCC的发生发展的关系十分密切,研究证实mTOR信号通路活化与HCC的低分化、恶性增殖、预后不良、早期复发高度相关[20]。并且我们的研究也发现TRIM44沉默下调了mTOR的磷酸化水平,而mTOR激活剂明显减弱了TRIM44沉默对肝癌细胞增殖的抑制作用。这结果提示TRIM44位于mTOR信号通路的上游。据研究显示TRIM家族中多个成员具有E3泛素连接酶活性,并且能泛素化和降解AMPK[21-22]。AMPK位于mTOR上游,是其活性抑制因子[23]。而TRIM44具有稳定TRIM家族成员的功能[21],因此TRIM44可能通过维持TRIM家族成员的E3泛素连接酶活性促进AMPK的泛素化降解,最终上调mTOR的活性,反之,TRIM44沉默则抑制mTOR的活性。这一可能的TRIM44沉默下调mTOR信号通路从而抑制肝癌细胞增殖的分子机制需要进一步的验证。
[1] DeSantis CE, Lin CC, Mariotto AB, et al. Cancer treatment and survivorship statistics[J]. CA Cancer J Clin, 2014, 64(4):252-271.
[2] Siegel R, Ma J, Zou Z, et al. Cancer statistics[J]. CA Cancer J Clin, 2014, 64(1):9-29.
[3] Forner A, Gilabert M, Bruix J, et al. Reply: heterogeneity of intermediate-stage HCC necessitates personalized management including surgery[J]. Nat Rev Clin Oncol, 2015, 12(1):10.
[4] Bosch FX, Ribes J, Díaz M, et al. Primary liver cancer: worldwide incidence and trends[J]. Gastroenterology, 2004, 127(5 Suppl 1):S5-S16.
[5] Kanwal F, El-Serag HB, Ross D. Surveillance for hepatocellular carcinoma: can we focus on the mission?[J]. Clin Gastroenterol Hepatol, 2015, 13(4):805-807.
[6] Psyrri A, Arkadopoulos N, Vassilakopoulou M, et al. Pathways and targets in hepatocellular carcinoma[J]. Expert Rev Anticancer Ther, 2012, 12(10):1347-1357.
[7] Ikeda K, Inoue S. TRIM proteins as RING finger E3 ubiquitin ligases[J]. Adv Exp Med Biol, 2012, 770:27-37.
[8] Zhang J, Fang L, Zhu X, et al. Ro52/SSA sensitizes cells to death receptor induced apoptosis by down-regulating c-FLIP(L)[J]. Cell Biol Int, 2012, 36(5):463-468.
[9] Schwamborn JC, Berezikov E, Knoblich JA. The TRIM-NHL protein TRIM32 activates microRNAs and prevents self-renewal in mouse neural progenitors[J].Cell, 2009, 136 (5):913-925.
[10]Uchil PD, Pawliczek T, Reynolds TD, et al. TRIM15 is a focal adhesion protein that regulates focal adhesion disassembly[J]. J Cell Sci, 2014, 127(pt 18):3928-3942.
[11]Wang Y, He D, Yang L, et al. TRIM26 functions as a novel tumor suppressor of hepatocellular carcinoma and its downregulation contributes to worse prognosis[J]. Biochem Biophys Res Commun, 2015, 463(3):458-465.
[12]Xing Y, Meng Q, Chen X, et al. TRIM44 promotes proliferation and metastasis in non-small cell lung cancer via mTOR signaling pathway[J]. Oncotarget, 2016,7(21):30479-30491.
[13]Ong CA, Shapiro J, Nason KS, et al. Three-gene immunohistochemical panel adds to clinical staging algorithms to predict prognosis for patients with esophageal adenocarcinoma[J]. J Clin Oncol, 2013, 31(12):1576-1582.
[14]Kashimoto K, Komatsu S, Ichikawa D, et al. Overexpression of TRIM44 contributes to malignant outcome in gastric carcinoma[J]. Cancer Sci, 2012, 103(11):2021-2026.
[15]Ong CA, Shannon NB, Ross-Innes CS, et al. Amplification of TRIM44: pairing a prognostic target with potential therapeutic strategy[J]. J Natl Cancer Inst, 2014, 106(5):dju050.
[16]Choi KJ, Baik IH, Ye SK, et al. Molecular targeted therapy for hepatocellular carcinoma: present status and future directions[J]. Biol Pharm Bull, 2015, 38(7): 986-991.
[17]Cargnello M, Tcherkezian J, Roux PP. The expanding role of mTOR in cancer cell growth and proliferation[J]. Mutagenesis, 2015, 30(2):169-176.
[18]Xu K, Liu P, Wei W. mTOR signaling in tumorigenesis[J]. Biochim Biophys Acta, 2014, 1846(2):638-654.
[19]费洪荣, 赵 莹, 王桂玲, 等. PI3K/mTOR双重抑制剂PF-04691502 诱导人胃癌SGC-7901 细胞凋亡[J]. 中国病理生理杂志, 2013, 29( 11):1962-1965.
[20]Matter MS, Decaens T, Andersen JB, et al. Targeting the mTOR pathway in hepatocellular carcinoma: current state and future trends[J]. J Hepatol, 2014, 60(4):855-865.
[21]Urano T, Usui T, Takeda S, et al. TRIM44 interacts with and stabilizes terf, a TRIM ubiquitin E3 ligase[J]. Biochem Biophys Res Commun, 2009, 383(2):263-268.
[22]Pineda CT, Potts PR. Oncogenic MAGEA-TRIM28 ubi-quitin ligase downregulates autophagy by ubiquitinating and degrading AMPK in cancer[J]. Autophagy, 2015, 11(5):844-846.
[23]Dong LX, Sun LL, Zhang X, et al. Negative regulation of mTOR activity by LKB1-AMPK signaling in non-small cell lung cancer cells[J]. Acta Pharmacol Sin, 2013, 34(2):314-318.
(责任编辑: 陈妙玲, 罗 森)
Effect of TRIM44 silencing on proliferation of hepatocellular carcinoma cells and its molecular mechanism
ZHANG Yun-yun, TIAN Yi, ZHOU Wei, SUN Mao, WU Yong-zhong
(ChongqingCancerInstitute,Chongqing400030China.E-mail:cqmdwyz@yahoo.com.cn)
AIM: To investigate the effect of tripartite motif-containing protein 44 (TRIM44) on the proliferation of hepatocellular carcinoma (HCC) cells and to study the molecular mechanism. METHODS: The expression of TRIM44 at mRNA and protein levels in normal liver tissues, HCC tissues, adjacent nontumor liver tissues, immortalized hepatocytes and hepatoma cell lines was determined by RT-qPCR and Western blot, respectively. The silencing ofTRIM44 was conducted by transfection of vector expressing shRNA targetingTRIM44 (shTRIM44) in the HCC cells, and the protein level of TRIM44 was measured by Western blot. The viability of the HCC cells was analyzed by MTS assay. The DNA synthesis of HCC cells was detected by Click-iT EdU Imaging Kit. The ability of anchorage-independent growth was determined by the method of colony formation on the soft agar. The effects of TRIM44 on the total protein and phosphorylation of mammalian target of rapamycin (mTOR) levels were measured by Western blot. The HCC cells were transfected with shTRIM44 and treated with mTOR agonist MHY1485, and the cell viability was analyzed by MTS assay. RESULTS: The mRNA and protein levels of TRIM44 in the HCC tissues were significantly higher than those in the adjacent nontumor liver tissues and normal liver tissues. In addition, the mRNA and protein levels of TRIM44 in the hepatoma cell lines were significantly higher than those in the immortalized hepatocytes.TRIM44 silencing significantly inhibited the viability of HCC cells and reduced the abilities of DNA synthesis and anchorage-independent growth of the HCC cells.TRIM44 silencing decreased the phosphorylation level of mTOR protein. MHY1485 significantly antagonized the inhibitory effect ofTRIM44 silence to the viability of HCC cells. CONCLUSION:TRIM44 silencing inhibits the proliferation of HCC cells possibly through down-regulating the activity of mTOR.
Tripartite motif-containing protein 44; Hepatocellular carcinoma; Mammalian target of rapamycin
1000- 4718(2016)11- 1972- 07
2016- 05- 30
2016- 09- 23
R735.7; R730.23
A
10.3969/j.issn.1000- 4718.2016.11.009
杂志网址: http://www.cjpp.net
△通讯作者 Tel: 023-65358274; E-mail: cqmdwyz@yahoo.com.cn
