小麦专用复方抗旱型浸种剂和包衣剂对小麦幼苗SOD、POD和CAT同工酶表达的影响
2016-11-08贾洪涛胡晓君邱奉同
贾洪涛,胡晓君,邱奉同,张 莉
(临沂大学生命科学学院,山东临沂 276005)
小麦专用复方抗旱型浸种剂和包衣剂对小麦幼苗SOD、POD和CAT同工酶表达的影响
贾洪涛,胡晓君,邱奉同,张 莉
(临沂大学生命科学学院,山东临沂 276005)
为从抗氧化酶同工酶表达方面了解小麦专用复方抗旱型浸种剂和包衣剂对小麦抗旱性的调节作用,以小麦品种临麦2号为材料,采用盆栽试验法,研究了小麦专用复方抗旱型浸种剂和包衣剂处理种子对干旱胁迫下小麦幼苗超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)同工酶表达的影响。结果表明,干旱胁迫下,小麦幼苗Mn-SOD和Fe-SOD表达量(活性)高于对照,而Cu/Zn-SOD同功酶表达量(活性)降低。与对照相比,干旱胁迫下,POD同工酶中的Rf 0.05、Rf 0.13、Rf 0.22和Rf 0.61条带表达量降低,抗旱型包衣剂和浸种剂处理种子均提高干旱胁迫下Rf 0.05、Rf 0.13和Rf 0.61条带的表达量,而普通包衣剂处理增加了Rf 0.18、Rf 0.22条带的表达量。干旱胁迫降低了CAT同功酶Rf 0.04、Rf 0.76条带的表达量,其中Rf 0.04表达量降幅较大,而抗旱型包衣剂和浸种剂处理种子均提高干旱胁迫下这两个条带的表达量,尤其是Rf 0.04的表达量显著超过对照。说明抗旱型包衣剂和浸种剂能够调控小麦幼苗抗氧化酶同工酶的表达,进而增强植株的抗氧化能力,提高小麦的抗旱性。
小麦;干旱胁迫;抗旱型浸种剂;抗旱型包衣剂;同工酶电泳
植物细胞内存在产生活性氧自由基的多种代谢途径和高效率清除这些自由基的抗氧化系统[1-2]。在逆境胁迫下,植株体内活性氧自由基产生与清除动态平衡被打破,并导致自由基离子积累,进而导致膜脂过氧化或脱脂化,破坏膜结构及其完整性,引起膜透性增大,引起代谢紊乱[2-3]。
超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)是三种重要的抗氧化酶。其中,SOD以Cu/Zn-SOD、Mn-SOD和Fe-SOD三种金属蛋白的形式存在。Cu/Zn-SOD被氰化钾和过氧化氢所抑制,Fe-SOD被过氧化氢所抑制,Mn-SOD对氰化钾和过氧化氢不敏感,且已证明这三种SOD均存在于植物体中[4-6]。干旱会引起小麦幼苗SOD活性增高[9]。在干旱胁迫下,抗旱的小麦品种SOD活性升高且具持续性[8]。5%和15% PEG6000胁迫后,小麦种子萌发期POD、CAT活性均呈上升趋势,但高浓度处理先升高而后降低[9]。在干旱和盐胁迫下,小麦根部POD同工酶活性升高,但没有酶带变化,叶片POD同工酶活性降低且在盐胁迫下少了一条谱带[10]。但有研究表明,干旱胁迫下小麦幼苗根和叶POD同工酶A、B、C区的酶带数目增加,且酶带颜色均有不同程度的加深,根和叶中POD活性显著增强[11];抗旱性不同的小麦品种功能叶POD同工酶活性和酶谱在受到干旱胁迫后均会发生变化,某些同工酶活性的增加与小麦的抗旱性呈正相关[12]。上述研究结果说明, 抗氧化酶活性与小麦抗旱性密切相关,因此可通过栽培等措施对其进行调节,以增强植株抗旱能力。
化学调控是提高小麦抗旱能力的一种重要方法。在干旱条件下施FA后小麦幼苗的SOD和POD活性增高,植株抗旱能力增强[13]。使用种衣剂可使SOD活性升高,起到延缓小麦植株衰老和诱导产生抗逆性的作用[14]。外源精胺能够提高水分胁迫下小麦叶片SOD、CAT 和 POD 活性,且抗旱性弱的品种的保护酶活性增幅高于抗旱性强的品种[15]。在干旱胁迫下,低浓度外源NO供体硝普钠处理可显著提高小麦幼苗叶片 SOD、POD和CAT 活性,缓解膜脂过氧化[16]。喷洒壳聚糖提高了干旱胁迫下小麦幼苗SOD、 POD 及 CAT 活性[17]。虽然单一成分的化控物质可提高小麦抗旱能力,但效果往往有限,因此开发有更好效果的复合型化控试剂或产品十分必要。
我们自主研发的小麦专用复方抗旱型浸种剂和包衣剂含有植物所需的大量和微量元素、植物生理活性物质、植物抗逆调控因子等成分,使用其处理小麦种子后,幼苗抗旱能力显著提高[18]。目前,有关小麦专用复方抗旱型浸种剂和包衣剂作用机理的研究很少。本研究试图从抗氧化酶同工酶表达方面探讨抗旱型包衣剂和浸种剂对小麦幼苗抗旱性的调节机制,以期为此种浸种剂和包衣剂的推广应用及新产品的开发提供参考。
1 材料与方法
1.1材 料
供试小麦为临麦2号,种子购于山东省临沂市种子公司。小麦专用复方抗旱型浸种剂和包衣剂均由本课题组成员贾洪涛主持研制。
1.2培养条件
试验采用盆栽方式,用土采自大田耕层,土壤先过35目筛,风干后混合均匀后装入直径30 cm、高35 cm的盆中,每盆6 kg。供试土壤pH 7.4,有机质含量281 mg·kg-1,速效氮含量126.8 mg·kg-1,速效磷含量38.4 mg·kg-1,速效钾含量118 mg·kg-1。
1.3材料处理
Yangzhou summit of 2018 World Green Design Forum held 7 60
选取大小一致、无病虫害的小麦种子用于试验。试验设对照(CK)、干旱(D)、普通包衣剂+干旱(PD)、复方抗旱型包衣剂+干旱(FBD)和复方抗旱型浸种剂+干旱(FJD)5种处理。CK和D处理的种子均不进行任何处理;PD处理使用江苏铜山农药总厂生产的铜农牌15%甲拌悬浮种衣剂对种子进行人工包衣,包衣剂与种子比例为1∶80;FBD处理使用小麦专用复方抗旱型包衣剂进行人工包衣,包衣剂与种子比例为1∶80;FJD处理使用小麦专用复方抗旱型浸种剂25 ℃浸泡种子12 h,中间搅拌三次,晾12 h播种,浸种与种子比例为1∶1.5。
1.4试验方法
将未处理及经过包衣或浸种处理过的种子分别均匀播种于相应的花盆中,每盆50粒,小麦幼苗长到3片真叶时每盆定苗40株,CK处理土壤含水量维持在田间持水量的70%(通过称重法控制),其他处理同时停止浇水进行干旱处理。待干旱处理的小麦幼苗叶片开始出现萎蔫、第一片叶下垂时,所有处理取样幼苗地上部分,进行同工酶样品制备。使用GE公司Image Scanner 扫描记录同工酶电泳图像,用Gel-Pro Analyzer分析记录条带数,测量每条电泳条带的累积光密度(IOD,亦称积分光密度),用每条条带的累积光密度值表示酶活性大小,并绘成Excel折线图。
1.4.1同工酶样品的制备
通过预试验比较了不同研究者常用的四种提取液电泳效果。四种提取液分别为提取液Ⅰ[0.10 mol·L-1Tris-HCl(pH 8.0)][19]、提取液Ⅱ[0.05 mol·L-1磷酸缓冲液(PBS,pH 6.6)][20]、提取液Ⅲ[0.10 mol·L-1Tris-HCl缓冲液(pH 7.2),含5% 蔗糖、0.5% (w/v) PVP、10 mmol·L-1β-巯基乙醇][21]和提取液Ⅳ[0.10 mol·L-1磷酸缓冲液(pH 7.8),含0.000 1 mol·L-1EDTA、1.0% (w/v) PVP和 0.5 % (v/v) Triton X-100]。提取液体积(mL)和新鲜材料重量(g)比例为 3∶1;研钵-20 ℃ 预冷,提取液4 ℃ 预冷;10 000 r·min-14 ℃ 离心30 min;Brandford法测定可溶性蛋白含量,小牛血清蛋白(BSA)作标准蛋白;-20 ℃保存提取液。
四种提取液中提取液Ⅲ和提取液Ⅳ电泳结果条带较清晰,数量较多,因此本研究选用这两种提取液作为同工酶提取液成分的依据,并对其作了改良,改良后的配方为0.10 mol·L-1Tris-HCl缓冲液(pH 7.2)中含10% 甘油、0.002 mol·L-1EDTANa2、0.10% (v/v) Triton X-100、4.0% (w/v)PVP、1.0% β-巯基乙醇,提取液分装后-20 ℃保存备用。改良后提取液电泳图像条带更清晰,条带更多,酶样品-20 ℃储藏时间更久。
1.4.2非变性聚丙烯酰胺凝胶电泳
采用非变性聚丙烯酰胺凝胶电泳;分离胶SOD浓度为10%,POD和CAT均为7.5%,浓缩胶浓度均为3%;上样量统一为15 μg可溶性蛋白;稳流,浓缩胶电流为10 mA·胶-1,样品进入分离胶后,加大电流至20 mA·胶-1[22]。电泳仪为美国通用电气公司(GE)EPS301电源和Hoefer○RminiVE电泳槽,每个样品分别点两个泳道。
SOD同工酶电泳胶染色采用氯化硝基四氮唑蓝(NBT)负染色法,不同SOD同工酶亚型的鉴定采用氯仿-乙醇抑制法、氰化钾(KCN)抑制法和过氧化氢(H2O2)抑制法[6,23]。
POD同工酶电泳胶染色参照吴少伯的方法[24]并进行了改良。改良后的染色液为每100 mL 0.05 mol·L-1pH 4.6乙酸缓冲液中包含0.1 g少量丙酮溶解的联苯胺、70 mg抗坏血酸,染色前加入0.75 mL 3%H2O2。剥下凝胶,放入染色液中30 ℃暗处孵育,直至褐色条带出现,去离子水冲洗后扫描。
CAT同工酶染色采用改良淀粉-碘反应方法[25]。具体步骤:灌制含0.1%可溶性淀粉的聚丙烯酰胺分离胶凝胶,电泳结束后,将胶剥离并冲洗两遍,放入含100 mL含0.15 g H2O2、0.14 g Na2S2O3·H2O 的pH 7.0 PBS缓冲液(现配现用)中,室温振荡15 min,冲洗两次,再放入去离子水配制的100 mL含0.0 747 g KI和1 mL冰乙酸(新鲜加入)的染色液中,室温下振荡至胶出现浅蓝色时用扫描仪扫描记录,连续扫描几次,透明的条带因蓝色渐浓而呈现时间较短暂。
2 结果与分析
2.1小麦幼苗叶片SOD同工酶亚型鉴定结果
SOD同工酶电泳结果共显示了5个条带,分别是Rf 0.25、Rf 0.35、Rf 0.37、Rf 0.55、Rf 0.58(图1)。H2O2抑制结果仅显示Rf 0.25条带,因为Cu/Zn-SOD在H2O2作用5 min内会急剧失活,Fe-SOD活性也受到较大影响,只有Mn-SOD活性比较稳定,故Rf 0.25条带应为Mn-SOD;氯仿-乙醇抑制结果仅显示Rf 0.55、Rf 0.58条带,Mn-SOD和Fe-SOD在氯仿-乙醇溶液里会迅速失活,而Cu/Zn-SOD能保持24 h内活性稳定,故条带Rf 0.55、Rf 0.58应为Cu/Zn-SOD;氰化钾抑制结果仅显示Rf 0.25条带,由于Mn-SOD在70 min内稳定,而Fe-SOD和Cu/Zn-SOD活性均不同程度下降,也说明Rf 0.25条带应为Mn-SOD;因三种抑制物质均对Fe-SOD活性有影响,所以三种抑制物质染色结果均看不到Rf 0.35和Rf 0.37,故这两个条带应为Fe-SOD。

1:SOD亚型;2:CK;3:加入H2O2;4:加入氯仿-乙醇;5:加入KCN
1:SOD isozyme isoforms;2:CK;3:Adding H2O2;4:Adding choroform-ethanol;5:Adding KCN
图1小麦叶片SOD亚型的鉴定结果
Fig.1Identification of SOD isozyme isoforms in the leaves of wheat seedlings
2.2小麦专用复方抗旱型浸种剂和包衣剂对小麦叶片SOD同工酶表达的影响
从图2和图3可以看出,干旱胁迫下,Mn-SOD活性变化幅度较小,其中FBD和FJD处理的Mn-SOD活性较CK略有升高;干旱胁迫下,各处理的Fe-SOD活性均较高,其中FBD和FJD处理较D处理下降,但略高于CK;Cu/Zn-SOD活性表现为FBD和FJD处理低于其他处理。
2.3小麦专用复方抗旱型浸种剂和包衣剂对小麦苗期叶片POD同工酶表达的影响
从图4和图5可以看出,干旱胁迫对小麦幼苗POD同工酶表达影响较大,使其条带数量、宽度和颜色深浅均发生了变化。相对CK处理,D处理中Rf 0.05、Rf 0.13、Rf 0.22和Rf 0.61条带表达量降低,而FBD和FJD处理表达量均提高,以Rf 0.61变化最显著,且D和FBD处理增加了Rf 0.33、Rf 0.39条带表达量;D和PD处理增加了Rf 0.18、Rf 0.22两个条带的表达量。
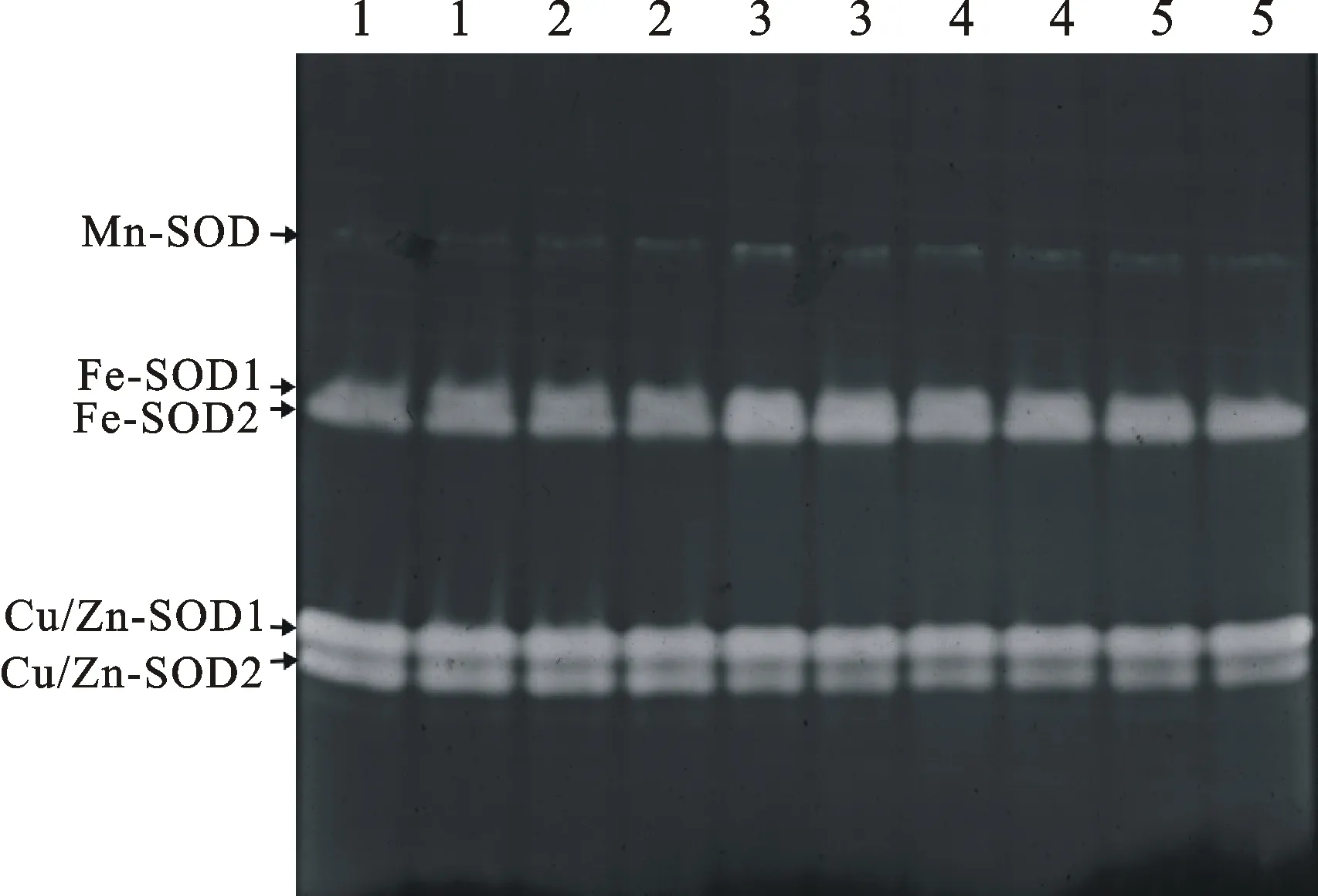
图2 不同处理下小麦苗期叶片SOD同工酶的表达Fig.2 Expression of SOD isozyme in the leaves of wheat seedlings under different treatments
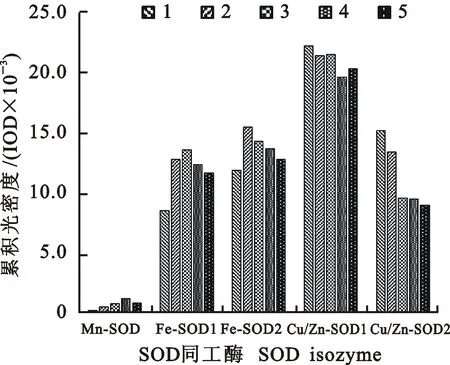
1:CK;2:D;3:PD;4:FBD; 5:FJD。下图同The same as in the following figures
图3不同处理下小麦苗期叶片SOD同工酶活性的差异
Fig.3Activity variation of SOD isozyme isoforms in the leaves of wheat seedlings under different treatments
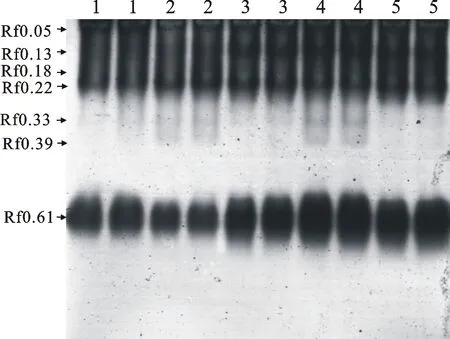
图4 不同处理下小麦苗期叶片POD同工酶的表达Fig.4 Expression of POD isozyme in the leaves of wheat seedlings under different treatments
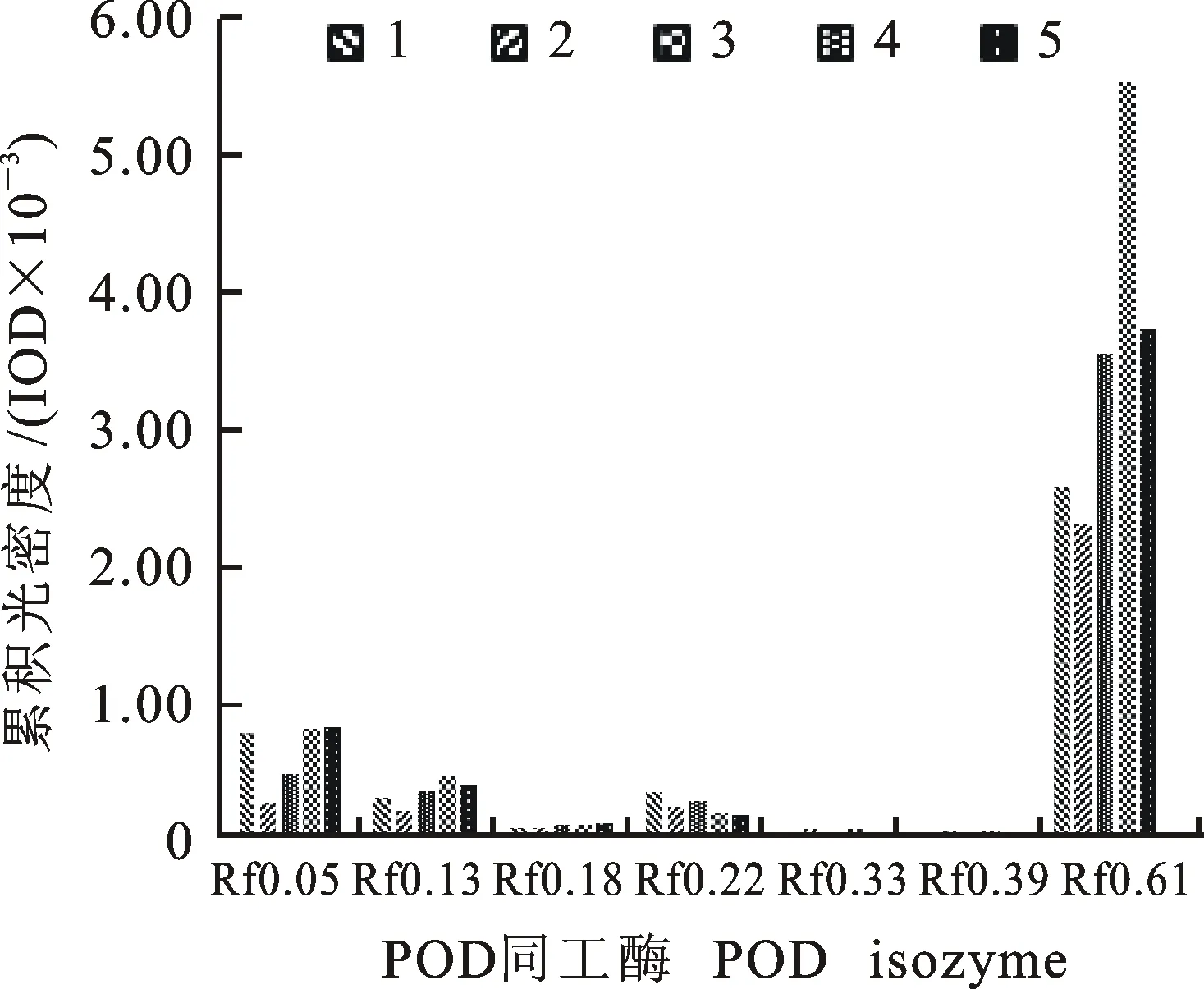
图5 不同处理下小麦苗期叶片POD同工酶活性的差异Fig.5 Activity variation of POD isozyme in the leaves of wheat seedlings under different treatments
2.4小麦专用复方抗旱型浸种剂和包衣剂对小麦苗期叶片CAT同工酶表达的影响
从图6和图7可以看出,小麦幼苗地上部分CAT同工酶电泳显示Rf 0.04、Rf 0.76两个条带。D处理明显降低了这两个条带的表达量,其中Rf 0.04变化幅度较大;FBD和FJD处理显著增加了Rf 0.04、Rf 0.76的表达量,尤其是Rf 0.04变化较明显。

图6 不同处理下小麦苗期叶片CAT同工酶的表达Fig.6 Expression of CAT isozyme in the leaves of wheat seedlings under different treatments
3 讨 论
干旱胁迫下,小麦幼苗SOD、POD和CAT同功酶活性和/或酶带发生了变化。其中, Fe-SOD活性升高,Cu/Zn-SOD活性降低,Mn-SOD活性变化较小。一些研究结果显示,干旱胁迫使SOD活性升高[7-10],这应该是Fe-SOD活性变化的结果。本研究中,干旱胁迫导致POD同工酶活性降低,这与部分研究者的结果不一致。抗旱型包衣剂和抗旱型浸种剂处理显著增加了POD同工酶Rf 0.05、Rf 0.13和Rf 0.61条带的表达量,尤其是Rf 0.61条带变化最显著,这与一些研究者的结果相一致[13-18]。干旱胁迫下,CAT同功酶Rf 0.04、Rf 0.76条带表达量均降低,抗旱型包衣剂和抗旱型浸种剂处理显著增加了这两个条带的表达量,这一结果尚未见文献报道。用抗旱型包衣剂和抗旱型浸种剂处理能够显著增加小麦幼苗的抗旱能力[18]。冯彩平[12]认为,POD同工酶酶带活性的增加与小麦的抗旱性存在正相关关系。本研究结果显示,抗旱型包衣剂包衣和抗旱型浸种剂浸种处理种子增加了Fe-SOD活性、POD同工酶活性和CAT同工酶活性,有助于增强小麦幼苗抗旱性。因此推测抗旱型包衣剂和抗旱型浸种剂对小麦幼苗抗氧化酶同工酶表达的影响可能与相关基因调控有关,但还需要进一步研究证实。
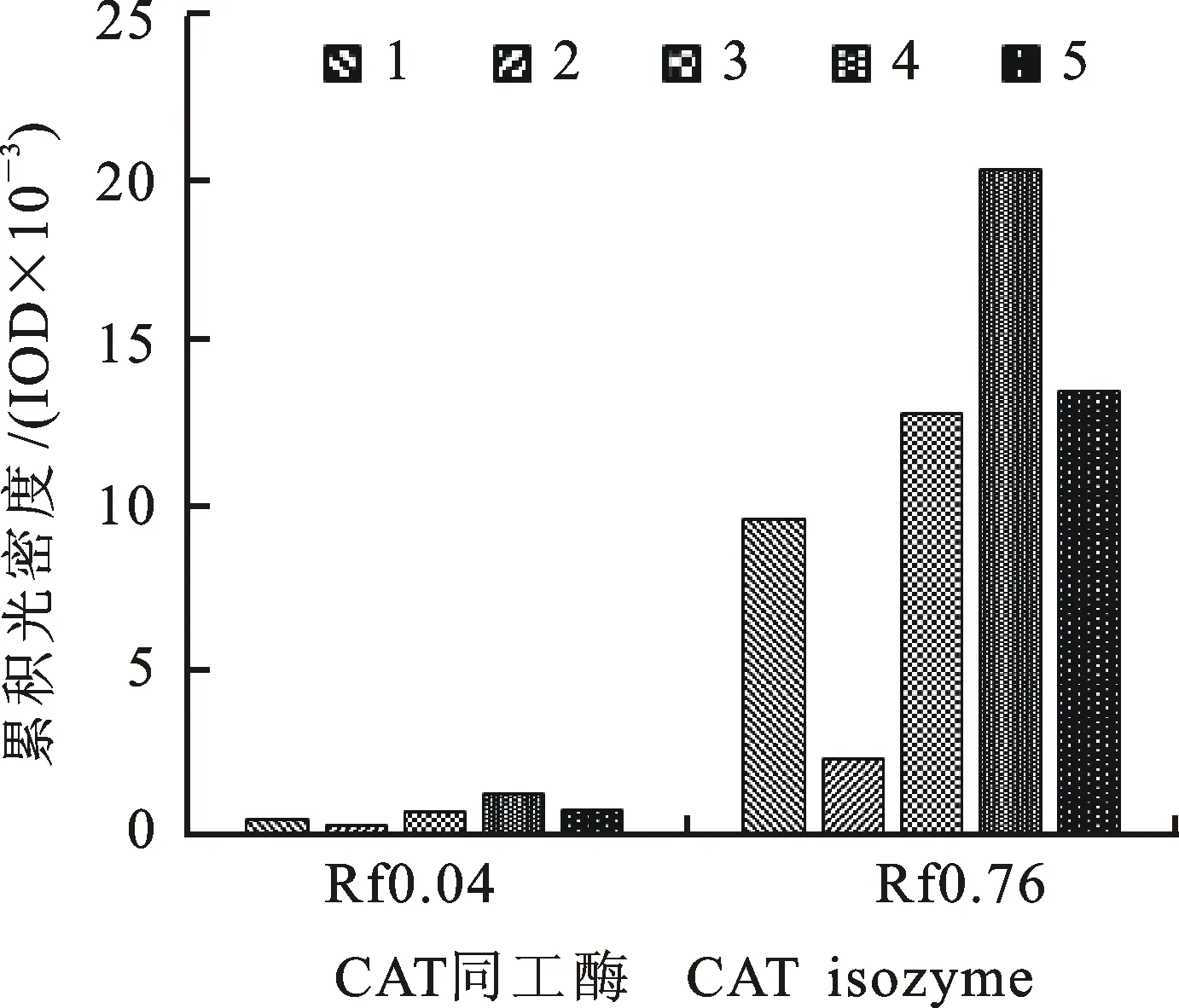
图7 不同处理下小麦苗期叶片CAT同工酶活性的差异Fig.7 Activity variation of CAT isozyme in the leaves of wheat seedlings under different treatments
[1]曹锡清.脂质过氧化对细胞与机体的作用[J].生物化学与生物物理进展,1986(2):17-21.
Cao X Q.The effects of lipid peroxidation on the cell and organism [J].ProgressinBiochemistryandBiophysics,1986(2):17-21.
[2]Elster F E.Oxygen activation and oxygen toxicity [J].AnnualReviewPlantPhysiology,1982,33:73-96.
[3]王宝山.生物自由基与植物膜伤害[J].植物生理学通讯,1988(2):12-16.
Wang B S.Biological free radicals and membrane damage of plants [J].PlantPhysiologyComunications,1988(2):12-16.
[4]Foster J G,Edwards G E.Localization of superoxide dismutase in leaves of C3and C4plants [J].PlantCellPhysiology,1980,21(5):895-906.
[5]Fridovich I.Superoxide dismutases [J].AnnualReviewofBiochemistry,1975,44:147-159.
[6]Lee D H,Kim Y S,Lee C B.The inductive responses of the antioxidant enzymes by salt stress in the rice (OryzasativaL.) [J].JournalofPlantPhysiology,2001,158(6):737-745
[7]黄国存,崔四平,马春红,等.干旱对小麦幼苗SOD活性和CaM水平的影响[J].华北农学报,1995,10(1):40-44.
Huang G C,Cui S P,Ma C H,etal.Influence of water stress on sod activities and calmodulin levels [J].ActaAgriculturaeBoreali-Sinica,1995,10(1):40-44.
[8]于同泉,刘宗萍,路 萍,等.水分胁迫小麦SOD、MDA动态变化与抗旱性的关系[J].北京农学院学报,1995,10(6):22-25.
Yu T Q,Liu Z P,Lu P,etal.The relations between dynamie state change of SOD,MDA in water-stressed wheat seeding and wheat drought resistance [J].JournalofBeijingAgriculturalCollege,1995,10(6):22-25.
[9]陈 军,高贵珍,方雪梅,等.干旱胁迫对小麦萌发期POD、CAT活性的影响[J].安徽农业科学,2014,42(17):5360-5361,5389.
Chen J,Gao G Z,Fang X M,etal.Effects of drought stress on activity of POD and CAT in wheat during generation period [J].JournalofAnhuiAgriculturalScience,2014,42(17) :5360-5361,5389.
[10]时丽冉,张文文.干旱和盐胁迫对小麦过氧化物同工酶的影响[J].衡水学院学报,2008,10(4):50-52.
Shi L R,Zhang W W.Effect of drought stress and salt stress on isoperoxidases of wheat [J].JournalofHengshuiUniversity,2008,10(4):50-52.
[11]裴冬丽.干旱胁迫对小麦幼苗过氧化物酶同工酶的影响[J].湖北农业科学,2011,50(8):1530-1532.
Pei D L.Effect of drought stress on peroxidase isoenzyme of wheat seedlings [J].HubeiAgriculturalScience,2011,50(8):1530-1532 .
[12]冯彩平.土壤干旱对冬小麦功能叶过氧化物酶同工酶及抗旱性的影响[J].水土保持通报,1996,16(4):46-50.
Feng C P.Effects of soil drought on peroxidase isoenzyme in winter wheat leaves and drought resistance [J].BulletinofSoilandWaterConservation,1996,16(4):46-50.
[13]陈玉玲,曹 敏,李云荫,等.干旱条件下黄腐酸对冬小麦幼苗中内源 ABA 和 IAA 水平以及SOD和POD 活性的影响[J].植物生理学通讯,2000,36(4):11-14.
Chen Y L,Cao M,Li Y Y,etal.Effect of fulvic acid on ABA,IAA and activities of superoxide dismutase and peroxydase in winter wheat seedling under drought condition [J].PlantPhysiologyComunications,2000,36(4):11-14.
[14]史国安,付国占,刘丰明.种衣剂对小麦幼苗膜脂过氧化及某些酶活性的影响[J].麦类作物学报,1997,17(4):62-64.
Shi G A,Fu G Z,Liu F M.Affects of seed-coating agent on membrane lipid peroxidation and some enzymes activities in wheat seedlings [J].JournalofTriticeaeCrops,1997,17(4):62-64.
[15]覃凤云,吕金印,陆 璃,等.外源精胺对水分胁迫下小麦幼苗保护酶活性的影响[J].西北植物学报,2006,26(1):86-91.
Qin F Y,Lu J Y,Lu L,etal.Effects of exogenous spermine on protect enzyme activities in wheat seedling under water stress [J].ActaBotanicaBoreali-OccidentaliaSinica,2006,26(1):86-91.
[16]李 慧,张 倩,王金科,等.外源 NO对干旱胁迫下小麦幼苗抗氧化酶活性的影响[J].江西农业学报,2010,22(12):1-3.
Li H,Zhang Q,Wang J K,etal.Effects of exogenous nitric oxide on activity of anti-oxidative enzymes in wheat seedling under drought stress [J].ActaAgriculturaeJiangxi,2010,22(12):1-3.
[17]朱启忠,姜 山,张真豪.干旱胁迫下壳聚糖对小麦幼苗保护酶活性的影响[J].干旱区资源与环境,2012,26(5):145-147.
Zhu Q Z,Jiang S,Zhang Z H.Effects of chitosan on protective enzyme activities in seedlings of wheat under drought stress [J].JournalofAridLandResourcesandEnvironment,2012,26(5):145-147.
[18]贾洪涛,曹善东,杜立树.小麦专用复方抗旱型浸种剂和包衣剂抗旱机理的初步研究[J].麦类作物学报,2010,30(3):496-499.
Jia H T,Cao S D,Du L S.Study of drought resistance effectiveness of wheat compound anti-drought seed soaking agent and seed coating agent on wheat seedlings under drought stress [J].JournalofTriticeaeCrops,2010,30(3):496-499.
[19]孙 静,王宪泽.盐胁迫对小麦过氧化物酶同工酶基因表达的影响[J].麦类作物学报,2006,26(1):42-44.
Sun J,Wang X Z.Effects of salt stress on gene expression of peroxidase isozyme in wheat [J].JournalofTriticeaeCrops,2006,26(1):42-44.
[20]Tanwja S R,Abrol Y P ,Sachar R C.Modulation of O-diphenolase and monophenolase enzymes during wheat grain development [J].CerealChemistry,1974,51:457-464.
[21]Siva R,Krishnamurthy K V.Isozyme diversity inCassiaauriculataL.[J].AfricanJournalofBiotechnology,2005,4 (8):772-775.
[22]赵世杰,史国安,董新纯.植物生理学实验指导[M].北京:中国农业科学技术出版社,2002:136-139,142-143.
Zhao S J,Shi G A,Dong X C.Experiment Guide of Plant Physiology [M].Beijing:Chinese Agricultural Science and Technology Press,2002:136-139,142-143.
[23]罗广华,王爱国,付爱根.鉴别超氧化物歧化酶类型的定位染色法[J].生物化学与生物物理进展,1996,23(4):356-359.
Luo G H,Wang A G,Fu A G.The location staining method for distinguishing diferent types of SOD [J].ProgressinBiochemistryandBiophysics,1996,23(4):356-359.
[24]吴少伯.植物组织中蛋白质及同功酶的聚丙烯酚胺凝胶盘状电泳[J].植物生理学通讯,1979(1):30-33.
Wu S B.The polyacrylamide disk gel electrophoresis of protein and isozyme from plant tissue [J].PlantPhysiologyComunications,1979(1):30-33.
[25]Manchenko G P.Handbook of Detection of Enzymes on Electrophoretic Gels(Second Edition) [M].Boca Raton London New York Washington,D C:CRC Press,2003:158-159.
The Effects of Compound Anti-drought Seed Soaking Agent and Seed Coating Agent on SOD, POD and CAT Isozyme Expression in Wheat Seedlings
JIA Hongtao,HU Xiaojun,QIU Fengtong,ZHANG Li
(Institute of Life Science,Linyi University,Linyi,Shandong 276001,China)
The effects of anti-drought seed soaking agent and seed coating agent on the oxidation reduction system isozyme expression were explored with wheat variety Linmai 2 under drought stress by controlling soil moisture in plant pots. The results showed that the Mn-SOD and Fe-SOD isozyme amount (activity) were higher than control; Cu/Zn-SOD isozyme amount (activity) was lower than control under drought stress; The POD isozyme expression results showed that the amount of band Rf 0.05, Rf 0.13, Rf 0.22 and Rf 0.61 was less than contrast under drought treatment; The amount of band Rf 0.05, Rf 0.13 and Rf 0.61 were exceeded over control by the treatments of “ anti-drought seed soaking agent” and “anti-drought seed coating agent” under drought stress; The new bands of Rf 0.18 and Rf 0.22 appeared under “drought stress” and “anti-drought seed soaking with drought stress” .The CAT expression showed that the amounts of band Rf 0.04 and Rf 0.76 were lower than control under “drought stress” treatment;The amounts of band Rf 0.04 and Rf 0.76,especially Rf 0.04 were significantly higher than control under the “anti-drought seed soaking with drought stress” and “anti-drought seed coating with drought stress” treatments. In summary, the seeds treated by drought-resistant seed soaking agent or seed coating agent could enhance isozyme expression by upgrading oxidation reduction system under drought stress, which increased drought-resistant and antioxidant ability of wheat seedling.
Wheat; Drought stress; Drought-resistant seed soaking agent; Drought-resistant seed coating agent; Isozyme electrophoresis
时间:2016-05-10
2015-11-15
2015-12-21
山东省科技攻关项目(2009GG10009040); 国家自然科学基金项目(No.31000744);山东省中青年科学家奖励项目(BS2010SW004)
E-mail:ht_jia@163.com
S512.1;S311
A
1009-1041(2016)05-0647-06
网络出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20160510.1625.032.html
