二穗短柄草成熟胚再生体系建立及农杆菌介导转化研究
2016-11-08胡文祥杨帼一赵惠贤刘香利
胡文祥,杨帼一, 刘 童, 赵惠贤,2, 刘香利,2
(1.西北农林科技大学生命科学学院,陕西杨陵 712100; 2.旱区作物逆境生物学国家重点实验室,陕西杨陵 712100)
二穗短柄草成熟胚再生体系建立及农杆菌介导转化研究
胡文祥1,杨帼一1, 刘 童1, 赵惠贤1,2, 刘香利1,2
(1.西北农林科技大学生命科学学院,陕西杨陵 712100; 2.旱区作物逆境生物学国家重点实验室,陕西杨陵 712100)
为建立二穗短柄草组织培养及遗传转化体系,以二穗短柄草BD21-3成熟胚为外植体,对成熟胚愈伤诱导、分化以及农杆菌侵染条件进行了研究。结果表明,在含有2.5 mg·L-12,4-D的培养基上,愈伤组织出愈率最高为93.83%;在含有0.2 mg·L-1KT的分化培养基上,分化率最高为38.18%;对二穗短柄草胚性愈伤组织农杆菌转化和GUS染色结果表明,侵染的过程中农杆菌菌液浓度OD600为0.6、侵染时间为5 min时转化率最高。
二穗短柄草;成熟胚;愈伤组织;农杆菌
拟南芥作为重要的模式植物在植物基因功能研究中起着非常重要的作用,但拟南芥属于双子叶植物,其与单子叶植物在进化上关系较远,而许多禾谷类植物的性状,如抗病性、穗与颖壳发育相关基因分析、籽粒产量和品质相关基因功能验证等研究,以拟南芥为模拟植物就有一定的限制和不足。水稻(Oryzasativa)作为第一个完成全基因组测序的禾本科植物,具有基因组小、功能基因组资源广泛等多方面的优势[1-2]。但水稻由于生长周期长,对生长环境要求高,尤其是在高产基因试验方面要求更高,且它很早就从冷季型禾本科中分离出来,其作为禾本科其他亚科的模式植物就有一定的局限性[3]。因此,需要寻找一种新型的、更合适的禾草模式植物。
二穗短柄草(Brachypodiumdistachyon)属于禾本科早熟禾亚科短柄草族短柄草属,一年生温带植物,与模式植物拟南芥有很多共同的特点:基因组小、植株矮小、自花授粉、繁殖力强、生长周期短、遗传资源丰富等[4-9]。拥有模式植物所必备的生物学特性,且与很多重要的谷类作物如小麦、大麦、燕麦、玉米、水稻、高粱等以及许多牧草与草坪草有较近的亲缘关系,其基因与小麦有很高的相似性和共线性[10-11]。二穗短柄草的这些特性使其成为重要的禾谷类模式植物。
作为功能基因组研究的模式植物,其组织培养再生及遗传转化体系的建立至关重要。目前,幼胚是二穗短柄草胚性愈伤组织在组织培养和遗传转化中使用最多的外植体。Draper等[10]研究发现,在加有2.5 mg·L-12,4-D的LS培养基上,二穗短柄草ABR1的胚性愈伤组织诱导率达到了45%;并初步证明了基因枪法转化二穗短柄草的可行性。Christiansen等[12]用基因枪法获得了二倍体二穗短柄草和多倍体二穗短柄草的转基因植株,但其得到的转化效率均在5%左右或更低。之后,更多学者尝试用基因枪法转化二穗短柄草,但基因枪转化法最显著的缺点是多拷贝插入,从而造成不规则的分离,稳定性差,试验成本较高。Vogel等[9]对来源于美国国家植物种质系统(NPGS)的二穗短柄草二倍体品系进行了农杆菌介导的转化研究,发现3个二倍体品系的再生率为4%~11%,多倍体品系的再生率为0.4%~15%。与基因枪转化相比,农杆菌介导的转化中,T-DNA插入常常是一个或最多几个位点的插入。用农杆菌转化拟南芥和水稻的转基因植物中,平均仅有1.5和1.4个拷贝T-DNA的插入[13-14]。农杆菌已被用来转化多种禾本科植物,成为某些禾本科植物转化的常规方法[15]。
成熟胚因取材方便,生理状态较为一致,是禾谷类作物如小麦、水稻等转基因研究的理想外植体。目前国内外对二穗短柄草成熟胚的再生与遗传转化研究报道很少[16]。因此,本试验以二穗短柄草品系BD21-3的成熟胚为外植体材料,通过对成熟胚愈伤组织诱导、分化、生根以及农杆菌侵染条件的研究,建立二穗短柄草成熟胚组织培养再生及转化体系,以期为小麦等禾谷类作物的品质改良、遗传转化以及基因功能验证奠定理论基础。
1 材料与方法
1.1材 料
二穗短柄草品系BD21-3温室种植后的成熟种子用于成熟胚培养;农杆菌菌株GV3101由本研究室保存;载体选用pCAMBIA3301[内含bar基因(膦丝菌素乙酰转移酶基因)、gus报告基因(β-葡糖苷酸酶基因)],其T-DNA区如图1。

35S promoter:花椰菜花叶病毒启动子 35S promoter:Cauliflower mosaic virus图1 pCAMBIA3301质粒T-DNA区Fig.1 T-DNA region of pCAMBIA3301
愈伤组织诱导及继代培养基:MS基本培养基 + 2.5 mg·L-12,4-D + 0.6 mg·L-1CuSO4+ 0.5 mg·L-1酸水解酪蛋白 + 30 g·L-1蔗糖 + 7 g·L-1琼脂;共培养培养基:MS基本培养基 + 2.5 mg·L-12,4-D + 0.2 mmol·L-1乙酰丁香酮 + 10 g·L-1葡萄糖(固体培养基另加7 g·L-1琼脂);愈伤组织分化培养基:MS基本培养基 + 500 mg·L-1酸水解酪蛋白 + 30 g·L-1蔗糖 + 7 g·L-1琼脂,添加不同浓度的KT、6-BA;生根培养基:1/2 MS基本培养基 + 0.6 mg·L-1IAA + 15 g·L-1蔗糖 + 7 g·L-1琼脂。以上培养基pH均为5.8,并在121 ℃下高压灭菌20 min后备用。
1.2方 法
1.2.1种子预处理
二穗短柄草BD21-3种子于超净工作台中用75%乙醇消毒45 s,无菌水冲洗3~5次,然后用0.2%的次氯酸钠消毒3~5 min,无菌水冲洗3~5次,于无菌水中浸泡过夜后在超净工作台中剥取成熟胚。
1.2.2愈伤组织诱导及继代
将剥取的二穗短柄草BD21-3的成熟胚置于愈伤组织诱导培养基中,24±2 ℃暗培养诱导愈伤组织,每2周继代一次,2周后统计愈伤组织出愈率,7~8周后统计胚性愈伤率。
出愈率= 愈伤组织数/接种的胚数×100%
胚性愈伤率=胚性愈伤组织数/接种的胚数×100%
1.2.3愈伤组织分化及植株再生
将生长7~8周的胚性愈伤组织转至生根培养基中,24±2 ℃、光暗比16/8 条件下培养2~3周后观察分化情况,统计分化植株。将分化后高于3 cm的幼苗转入生根培养基进行生根。再生率=再生绿苗数/胚性愈伤组织数×100%
1.2.4成熟胚愈伤组织的农杆菌转化及瞬时表达检测
胚性愈伤组织农杆菌转化:挑取含有质粒pCAMBIA3301的农杆菌GV3101单菌落,接种于含有50 mg·L-1卡那霉素和30 mg·L-1利福平的YEB液体培养基;28 ℃培养48 h后,按1%接种扩大培养;28 ℃培养4~6 h后至OD600值约为0.6和0.8, 5 000 r·min-1离心5 min收集菌体;用液体的共培养培养基悬浮,分别侵染生长状态良好的胚性愈伤组织,分别侵染5 min 和15 min。侵染结束后用无菌滤纸吸干成熟胚胚性愈伤组织表面液体;将胚性愈伤组织接种到固体的共培养培养基中,24±2 ℃黑暗共培养3 d 后进行GUS组织化学染色。
GUS组织化学染色:GUS组织化学检测参照Jefferson等[17]的方法进行。共培养3 d后的胚性愈伤组织用无菌水洗去表面的农杆菌,浸入X-Gluc溶液,37 ℃保温染色24~48 h,95%乙醇脱色,观察蓝色反应,统计蓝色愈伤组织数。
GUS阳性率=出现蓝斑愈伤数/转化愈伤数×100%
2 结果与分析
2.1愈伤发生与胚性愈伤组织的形成
二穗短柄草BD21-3成熟胚接到继代培养基上7 d左右就会形成乳白色的愈伤组织(图2A),愈伤组织生长状况良好。随着不断继代,愈伤组织进一步生长膨大,5~7周后开始形成淡黄色、颗粒状的胚性愈伤组织(图2B)。从表1可以看出,在诱导培养基上,两批材料的出愈率均较高,分别为92.5%和93.8%,胚性愈伤组织形成率分别为62.3%和55.2%。
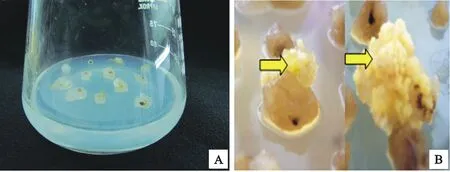
A:二穗短柄草成熟胚诱导的愈伤组织;B:继代后形成的胚性愈伤组织
A:Callus induced by the mature embryo ofBrachypodiumdistachyon;B:Embryonic callus after several rounds

图2 成熟胚愈伤组织诱导Fig.2 Callus induction of mature embryo表1 成熟胚的诱导率Table 1 Frequency of callus induction from mature embryo
2.2不同激素配比对二穗短柄草成熟胚胚性愈伤组织再生的影响
二穗短柄草成熟胚愈伤组织经过继代7~8周后开始形成致密的胚性愈伤组织,将胚性愈伤组织转至加有不同浓度KT的分化培养基中,生长2周后愈伤组织逐渐分化出绿芽(图3A),进一步分化形成植株(图3B),分化结果统计如表2。由表2可以看出,随着KT浓度的增加,愈伤组织分化率成先上升后降低的趋势,在KT浓度为0.2 mg·L-1时,其再生率达到最高,为38.18%。曾尝试在含有0.2 mg·L-1KT的分化培养基中加入不同浓度的6-BA,结果发现加入6-BA后,胚性愈伤组织并没有正常分化和再生,且随着6-BA浓度的增加,褐化逐渐加深并且最终导致愈伤死亡。
2.3IAA对植株生根的影响
分化小苗转至生根培养基后逐渐分化形成根(图4),观察10株分化小苗的生根状况发现,10 d后,加入IAA的生根培养基中分化小苗可形成根,而不加IAA的培养基中的分化小苗则未出现根;20 d后两种处理的分化小苗均能形成根,但加IAA的培养基中小苗生根数目较多(图5)。

A:胚性愈伤组织分化出绿芽;B:胚性愈伤组织分化
A:Green buds on embryonic callus;B:Differentiation of embryonic callus
图3胚性愈伤组织分化
Fig.3Differentiation of embryonic callus

图4 二穗短柄草再生植株及其生根Fig.4 Regeneration and rooting of Brachypodium distachyon

图5 不同时间下两种培养基中分化小苗的生根数Fig.5 Number of roots differentialion seeding with different duration in two types medium
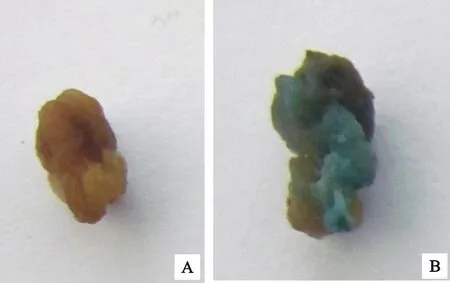
A:对照;B:侵染后愈伤组织
A:Control;B:Callus after infection
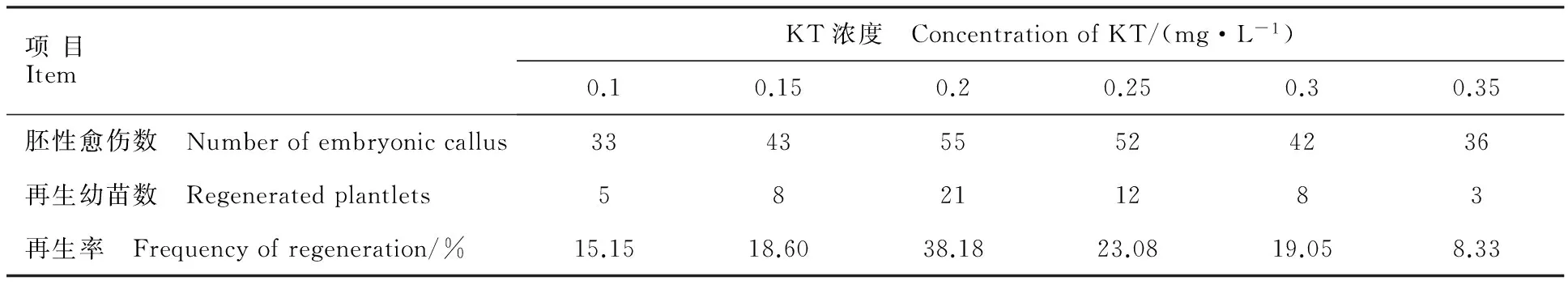
图6 GUS染色Fig.6 GUS staining表2 不同KT浓度对二穗短柄草再生的影响Table 2 Effects of different concentration of KT on regeneration of Brachypodium distachyon
2.4成熟胚胚性愈伤组织的农杆菌侵染和瞬时表达检测
为了确定二穗短柄草成熟胚愈伤组织农杆菌侵染的最适菌液浓度和侵染时间,选用生长状态良好的胚性愈伤组织,用含有GUS报告基因质粒pCAMBIA3301的农杆菌侵染,共培养3 d后进行GUS瞬时染色。对照未染色,转化后的愈伤组织被染成蓝色(图6),对GUS染色后的阳性愈伤组织进行统计,结果如表3。随着侵染时间延长,愈伤组织褐化增加,GUS阳性率降低,菌液浓度OD600为0.6、侵染时间为5 min时,GUS阳性率最高,为66.00%。

表3 不同菌液浓度与侵染时间的GUS染色结果Table 3 GUS activity under different bacterial concentration and different infection duration
3 讨 论
在植物组织培养中,2,4-D是人们常用的、最有效的植物激素之一。低浓度的2,4-D有促进植物生长发育的作用,当2,4-D浓度为1.0~3.0 mg·L-1时,对胚性愈伤组织的形成具有非常重要的作用[18]。在早期的研究中,Bablak等[19]将3个二穗短柄草品系B200、B373和B377的成熟胚剥至N6和MS培养基上,发现加有2.5 mg·L-12,4-D的MS和N6培养基最适合二穗短柄草胚性愈伤组织的形成。之后,Draper等[10]以二穗短柄草ABR1幼胚为材料,研究其胚性愈伤组织诱导与再生,同样发现在加了2.5 mg·L-12,4-D的LS培养基上,其胚性愈伤组织诱导率为45%。本研究选用了Bablak所述的含2.5 mg·L-12,4-D的MS培养基进行成熟胚愈伤组织诱导,发现二穗短柄草BD21-3成熟胚愈伤组织的诱导率可达90%以上,且胚性愈伤组织诱导率达到了50%以上,与吴雪莉等[20]以幼胚为外植体得到的愈伤组织和胚性愈伤组织研究结果相似。同时高于Draper等[10]的报道,说明成熟胚可高效地诱导形成胚性愈伤组织。
在愈伤组织分化方面,Vogel等[9]报道,在0.23 mg·L-1KT下,短柄草19个基因型的幼胚愈伤组织分化率在0~90%。董 芳等[21]报道,在加有0.2 mg·L-1KT 和0.5~1.0 mg·L-16-BA下,ABR6和ABR102的幼胚最高分化率分别为64.12%和71.25%;KT较6-BA效果更明显,6-BA浓度越高褐化现象越严重,且分化率没有明显提高。本试验对KT浓度和6-BA浓度进行了摸索,发现在加有0.2 mg·L-1KT的分化培养基中,分化率最高达到了38.18%,同时还发现在加了6-BA的培养基中,胚性愈伤褐化严重,且分化受到严重抑制,最终导致愈伤组织死亡。
二穗短柄草遗传转化研究主要侧重于幼胚的转化。Vogel等[9]构建了一种农杆菌介导的遗传转化体系,成功转化了19个株系中的10种,其转化率为0.4%~15%。随后,Vogel等[22]在原有的转化体系基础上改进了转化方法,用滤纸替代培养基进行共培养,使得最终的转化率提高了15倍,使得二穗短柄草的遗传转化达到了与水稻转化体系相同的效率[23]。本试验对农杆菌侵染条件进行了摸索,初步确定二穗短柄草BD21-3的农杆菌侵染菌液的浓度为OD600=0.6,侵染时间为5 min,其GUS活性最高,达到了66.00%。该研究为二穗短柄草遗传转化以及利用二穗短柄草BD21-3进行小麦、大麦等谷类作物的基因功能研究奠定了基础。
[1]Havukkala I J.Cereal genome analysis using rice as a model [J].CurrentOpinioninGenetics&Development,1996,6:711-714.
[2]Tyagi A K,Mohanty A.Rice transformation for crop improvement and functional genomics [J].PlantScience,2000,158:1-18.
[3]Gaut B S.Evolutionary dynamics of grass genomes [J].NewPhytologist,2002,154:15-28.
[4]Caetano-Anolles G.Evolution of genome size in the grasses [J].CropScience,2005,45:1809-1816.
[5]Catalan P,Olmstead R G.Phylogenetic reconstruction of the genusBrachypodiumP.Beauv.(Poaceae) from combined sequences of chloroplastndhFgene and nuclear ITS [J].PlantSystemEvolution,2000,220:1-19.
[6]Kellogg E A.Evolutionary history of the grasses [J].PlantPhysiology,2001,125:1198-1205.
[7]王宏归,王保莉,林辰涛,等.二穗短柄草Bd21的形态学观察[J].西北农业学报,2007,16(6):296-300.
Wang H G,Wang B L,Lin C T,etal.Morphology ofBrachypodiumdistachyon(L.) [J].ActaAgriculturaeBoreali-occidentalisSinica,2007,16(6):296-300.
[8]Shi Y,Draper J,Stace C.Ribosomal DNA variation and its phylogenetic implication in the genusBrachypodium(Poaceae) [J].PlantSystematicsandEvolution,1993,188:125-138.
[9]Vogel J P,Garvin D F,Leong O M,etal.Agrobacterium-mediated transformation and inbred line development in the model grassBrachypodiumdistachyon[J].PlantCellTissueandOrganCulture,2006,84:199-211.
[10]Draper J,Mur L A J,Jenkins G,etal.Brachypodiumdistachyon.A new model system for functional genomics in grasses [J].PlantPhysiology,2001,127:1539-1555.
[11]Hasterok R,Marasek A,Donnison I S,etal.Alignment of the genomes ofBrachypodiumdistachyonand temperate cereals and grasses using bacterial artificial chromosome landing with fluorescenceinsituhybridization [J].Genetics,2006,173:349-362.
[12]Christiansen P,Andersen C H,Didion T,etal.A rapid and efficient transformation protocol for the grassBrachypodiumdistachyon[J].PlantCellReports,2005,23:751-758.
[13]Feldmann K A.T-DNA insertion mutagenesis inArabidopsis:Mutational spectrum [J].PlantJournal,1991,1:71-82.
[14]Jeon J S,Lee S,Jung K H,etal.T-DNA insertional mutagenesis for functional genomics in rice [J].PlantJournal,2000,22:561-570.
[15]Cheng M,Lowe B A,Spencer T M,etal.Invited review:Factors influencingAgrobacterium-mediated transformation of monocotyledonous species [J].InVitroCellular&DevelopmentalBiology-Plant,2004,40:31-45.
[16]吴雪莉.基因枪法和农杆菌介导二穗短柄草(Brachypodiumdistachyon)成熟胚愈伤组织的遗传转化[D].南京:南京农业大学,2009.
Wu X L.Particle bombardment andAgrobacterium-mediated transformation ofBrachypodiumdistachyonthrough embryogenic calli derived from mature embryos [D].Nanjing:Nanjing Agricultural University,2009.
[17]Jefferson R A,Kavanagh T A,Bevan M V,etal.GUS fusion:β-Glucuronidase as a sensitive and versatile gene fusion marker in higher plants [J].EMBOJournal,1987,6:3901-3907.
[18]Chang Y,von Zitzewitz J,Hayes P M,etal.High frequency plant regeneration from immature embryos of an elite barley cultivar(HordeumvulgareL.cv.Morex) [J].PlantCellReports,2003,21:733-738.
[19]Bablak P,Draper J,Davey M R,etal.Plant regeneration and micropropagation ofBrachypodiumdistachyon[J].PlantCellTissueandOrganCulture,1995,42:97-107.
[20]吴雪莉,刘金星,Nielsen K K,等.二穗短柄草幼胚再生体系及农杆菌介导转化的初步研究[J].草业学报,2010,19(5):9-16.
Wu X L,Liu J X,Nielsen K K,etal.Agrobacterium-mediated transformation ofBrachypodiumdistachyonthrough embryogenic calli derived from immature embryos [J].ActaPrataculturaeSinica,2010,19(5):9-16.
[21]董 芳,蔡高磊,张 荻,等.二穗短柄草幼胚愈伤组织诱导及高频再生体系的建立[J].麦类作物学报,2010,30(6):1048-1052,1064.
Dong F,Cai G L,Zhang D,etal.Callus induction and establishment of high-frequency plant regeneration system from immature embryos ofBrachypodiumdistachyon[J].JournalofTriticeaeCrop,2010,30(6):1048-1052,1064.
[22]Vogel J,Hill T.High-efficiencyAgrobacterium-mediated transformation ofBrachypodiumdistachyoninbred line Bd21-3 [J].PlantCellReports,2008,27:471-478.
[23]Tyagi A,Mohanty A.Rice transformation for crop improvement and functional genomics [J].PlantScience,2000,158:1-18.
Establishment ofBrachypodiumdistachyonRegeneration System from Mature Embryos and the Study ofAgrobacterium-mediated Transformation
HU Wenxiang1,YANG Guoyi1,LIU Tong1,ZHAO Huixian1,2,LIU Xiangli1,2
(1.College of Life Science,Northwest A&F University,Yangling,Shaanxi 712100,China;2.State Key Laboratory of Crop Stress Biology for Arid Areas,Yangling,Shaanxi 712100,China)
To establish the regeneration and genetic transformation system ofBrachypodiumdistachyon,the mature embryos ofBrachypodiumdistachyonBD21-3 were used as explant,and the factors affecting the callus induction and theAgrobacteriuminfection were studied in this research. The results showed that the frequency of callus induction was 93.83% on the basic media with 2.5 mg·L-12,4-D. The maximum regeneration frequency (38.18%) was observed on the differential media containing 0.2 mg·L-1KT. The GUS staining showed that the highest GUS activity was obtained when the concentration ofAgrobacteriumwas OD600=0.6 and infection incubation duration was 5 min.
Brachypodiumdistachyon; Mature embryo; Callus;Agrobacterium
时间:2016-05-10
2015-12-09
2016-01-04
国家自然科学基金项目(31101205);国家生命科学与技术人才培养基地大学生创新性实验计划项目(1310712082)
E-mail:huwenxiangapple@foxmail.com
刘香利(E-mail: liuxianglii@163.com)
S519;S330
A
1009-1041(2016)05-0589-06
网络出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20160510.1623.016.html
