白菜型油菜自交亲和性、角果及种子相关性状的QTL定位
2016-10-14孙倩倩王晓武梁建丽申书兴
孙倩倩,武 剑,程 锋,王晓武,梁建丽,申书兴
白菜型油菜自交亲和性、角果及种子相关性状的QTL定位
孙倩倩1,2,武 剑2,程 锋2,王晓武2,梁建丽2,申书兴1
(1河北农业大学园艺学院/河北省蔬菜种质创新与利用重点实验室/河北省蔬菜产业协同创新中心,河北保定071000;2中国农业科学院 蔬菜花卉研究所,北京100081)
【目的】研究白菜型油菜的自交亲和性、角果及种子相关性状的QTL定位,为白菜型油菜自交亲和品种、优良自交系的选育及其产量和品质的遗传改良提供依据。【方法】以自交亲和性较差的菜薹L58和自交亲和性较好的白菜型油菜R-O-18构建的包含117个株系的RIL群体为试验材料,利用已构建的包括372个InDel标记的分子遗传图谱,采用区间作图法(IM)对花期亲和指数、结角率、角果长度、角果喙长度、角果长宽比、角果喙长度/角果长度、每角果种子数、千粒重及种子颜色9个性状分别进行QTL分析。【结果】RIL群体在自交亲和性、角果及种子相关的9个性状上表现为连续变异,并且变异幅度较大,均呈正态分布或偏正态分布,具有典型的数量性状遗传特点。花期亲和指数与结角率、角果长度、角果喙长度、角果长宽比、每角果种子数、千粒重均存在极显著的正相关,其中花期亲和指数与每角果种子数间的相关系数最大,达到0.8487。与其他几个性状的相关性大小依次为:千粒重>结角率>角果喙长度>角果长度>角果长宽比。每角果种子数及千粒重均与结角率、角果长度、角果喙长度、角果长宽比呈现极显著正相关,而每角果种子数与千粒重之间也存在极显著的正相关(0.6477)。结角率、角果长度、角果喙长度与角果长宽比之间呈现显著正相关;而角果喙长/角果长除了与角果长呈负相关,角果喙长度存在极显著正相关性外,与其他性状均不相关。共检测到12个QTL位点,其中6个QTL位于A09连锁群,10个QTL解释大于10%的表型变异。2个控制花期亲和指数的QTL均位于A09连锁群上,分别可解释13.1%和16.7%的表型变异;6个角果相关性状的QTL,分别位于A06、A09和A10连锁群上,单个QTL可解释13.4%—17.7%的表型变异;4个种子相关性状的QTL,分别位于A02、A05、A06和A09连锁群上,可解释7.9%—42.1%的表型变异,位于A09连锁群的种子颜色QTL为主效位点,其加性效应值为1.22。【结论】共检测到12个控制自交亲和性(2个)、角果(6个)及种子相关性状(4个)的QTL,其中,2个控制自交亲和性的QTL,与前人得到的调控自交亲和性主位点(S位点)不同,可能为控制自交亲和性的非主效QTL位点。1个种子颜色QTL为主效位点。
白菜型油菜;自交亲和性;角果;种子相关性状;QTL
0 引言
【研究意义】白菜类作物(,2=AA=20)是十字花科芸薹属重要的蔬菜和油料作物,包括大白菜、不结球白菜、芜菁、菜薹(心)和白菜型油菜等多个栽培种群。其中白菜型油菜具有早熟、抗旱、黄籽、含油量高等优良性状,是世界上重要的油料作物[1-3]。对于此类作物,种子产量和质量是重要的农艺性状。由于绝大部分白菜类作物属于自交不亲和类型,其品种群体是由不同基因型个体组成的杂合群体,限制了产量的进一步提高。利用杂种优势是提高白菜型油菜产量和品质的重要途径。由于其自交不亲和的特性,在育种过程中对材料的纯化必须进行人工剥蕾自交,制种成本高,而且后代由于自交衰退严重[1,4],给利用自交不亲和系或雄性不育系的杂交种选育工作带来很大的难度。但是,在白菜型油菜中存在自交亲和性较强的品种类型。因此,培育自交亲和品种,加快优良自交系的选育,是利用杂种优势来提高白菜型油菜产量和品质的最有效途径[5]。另外,此类作物的种子产量还受单株角果数、每角果种子数、角果长、角果宽、种子千粒重等产量构成相关性状的直接影响。而与黑籽或褐籽相比,黄籽品种种皮较薄,种子含油量及蛋白质含量较高,而多酚、木质素和纤维素含量较低,在食用油和饼粕品质方面远远优于黑籽或褐籽品种[6],可见种皮颜色与种子质量有关。因此,开展其自交亲和性、角果以及种子相关性状的遗传研究对其育种工作具有十分重要的意义。【前人研究进展】芸薹属二倍体植物属于自交不亲和种,研究表明,S位点糖蛋白基因(SLG)和S受体激酶基因(SRK)是2个控制自交不亲和性关键因子[7-11],在白菜型油菜中分离出一个控制自交不亲和性的M位点,主要参与SRK信号的传导[12]。而关于芸薹属植物的自交亲和性,研究认为主要是由于自交不亲和基因SLG或SRK功能突变导致的[13-16]。有关角果及种子相关性状的QTL研究在甘蓝型油菜和芥菜型油菜中进行的比较深入,许多国内外研究者利用不同作图群体对角果长度、每角果种子数、结角率和千粒重等进行了QTL定位及相关分析,并定位了控制相应性状的主效QTL[17-21],但在白菜型油菜研究较少。王玉刚等[22]利用白菜自交不亲和系和白菜型油菜自交系为亲本及杂交获得的4个基本世代为材料,对其角果相关性状进行遗传分析,发现角果相关各性状均以多基因遗传为主,角果宽度性状受环境因素影响较小;角果长度、喙长受环境因素影响较大。Bagheri等[3]利用白菜型油菜与菜薹构建的RILs群体对角果及种子相关性状进行了QTL定位,检测到7个控制角果相关性状的QTL,2个控制种子颜色的QTL;Lou等[16]利用白菜型油菜(YS-L143)与不同类型白菜的DH群体,定位了6个种子相关性状(包括角果长、宽度、角果喙长、种皮颜色及种子种性状)的QTL;Kebede等[23]利用白菜型黄籽莎逊(yellow sarson)油菜构建的RILs群体对种子颜色进行了QTL定位,检测到4个QTL,其中主效位点位于A09上;原玉香等[6]用大白菜DH群体进行了种皮颜色的遗传分析,检测到9个控制种皮颜色的QTL,其中最重要的QTL位于A06上。【本研究切入点】利用2个自交亲和性有差异的材料为亲本,其后代个体的自交亲和性存在显著差异的RIL群体,分析控制自交亲和性状的QTL,特别是非主效QTL,虽然已经检测到一些控制白菜型油菜角果及种子相关性状的QTL,但由于这些性状都是多基因控制的复杂性状,仍有一些QTL未被检测到。【拟解决的关键问题】本研究以自交亲和性强的白菜型油菜R-O-18和自交亲和性差的菜薹L58及其构建的包含117个株系的RIL群体为材料,对自交亲和性、结角率、角果长度、角果喙长度、角果长宽比、角果喙长/角果长、每角果种子数、种子千粒重以及种子颜色9个性状进行QTL定位的研究,旨在为此类作物自交亲和品种的选育以及产量与品质的遗传改良提供依据。
1 材料与方法
1.1 试验材料
R-O-18为白菜型油菜(ssp.),自交亲和性较强,叶缘有缺刻、生长势强、开花早、种子较大,种皮黄色;L58为菜薹(ssp.)自交亲和性较差,叶缘无齿、分枝性强、开花早、种子较小、种皮深褐色。以R-O-18和L58为亲本,构建的包含130个株系的重组自交系(recombination inbred lines,RILs)群体(由荷兰瓦赫宁根大学提供),选取其中的117个株系作为供试材料。
1.2 花期人工授粉
2015年3月,所有供试的117个RIL株系及两亲本播种于中国农业科学院蔬菜花卉研究所的纱棚内,每株系种植3个单株,随机排列,常规栽培管理。播种35 d后,植株陆续进入花期,在每个单株上选取一个生长健壮的枝条,去掉枝条上盛开的花朵后,用授粉袋套上整个花序,并挂牌标记。待授粉袋中出现超过15朵盛开的花朵时,将同一株系同株花粉均匀的抹到盛开的新鲜花朵的柱头上,每株授15—30个花朵,并摘除其余所有的未开放的花蕾,并记录授粉花朵数目,继续套袋。10 d后脱去授粉袋,待成熟时收获套袋角果。
1.3 性状调查
对RIL群体的自交亲和性、角果及种子相关的9个性状进行调查,包括花期亲和指数(compatible index of flowering time,CIFT)、结角率(pods rate,PR)、角果长度(silique length,SL,图1-A)、角果喙长度(silique beak length,SBL,图1-A)、角果长宽比(silique length/silique width,SL/SW)、角果喙长/角果长(silique beak length/silique length,SBL/SL)、每角果种子数(number of seeds per silique,NSPS)、种子千粒重(thousand seed weight,TSW)、种子颜色(seed color,SC,图1-B)。其中,花期亲和指数=结籽数/花期授粉花朵数;结角率=角果数/授粉花数;每角果种子数=种子数/角果数;角果长度(cm)、宽度以及角果喙的长度(cm)等性状通过对角果拍照记录,然后每个单株选取5个角果,用ImageJ软件测量计算后,求取3个重复植株性状的平均值;种子千粒重(g)=1000粒种子的重量;种子颜色作为数量性状,将其从黄色到黑色分为6个等级(图1-B)。
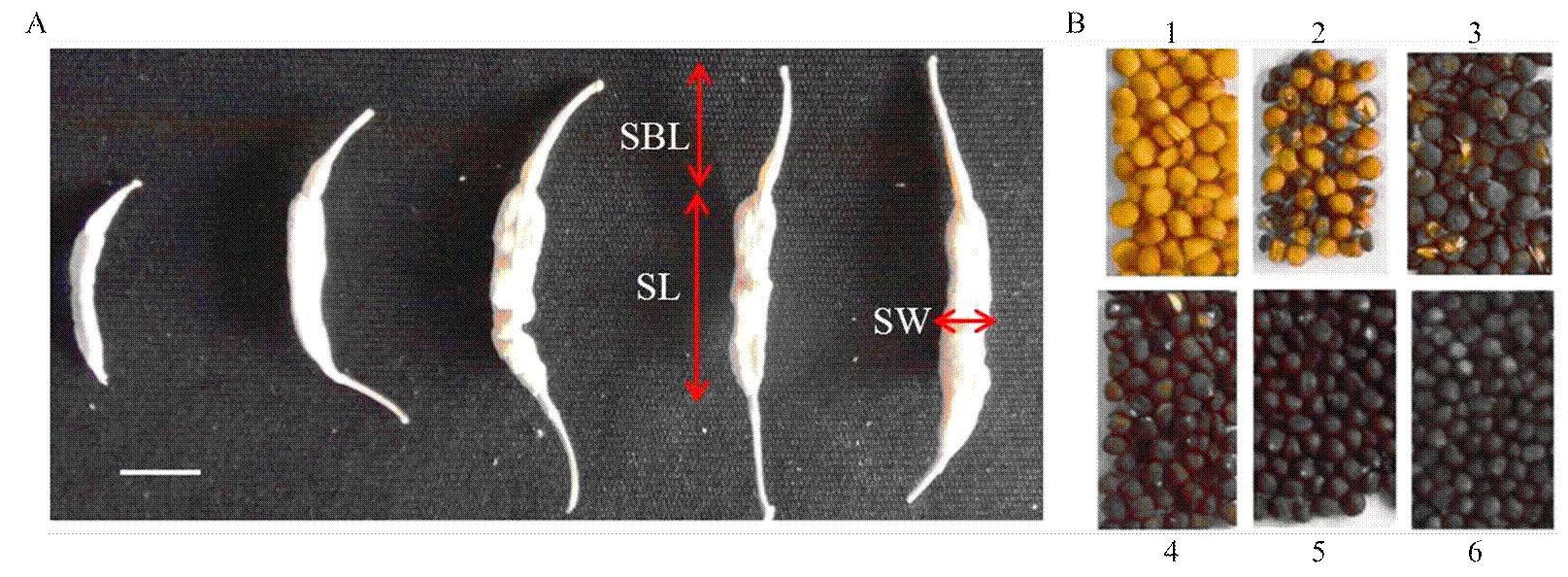
A:角果性状。角果长度(SL),角果宽度(SW),角果喙长度(SBL);B:种子颜色的6个等级
1.4 数据整理及QTL分析
采用R软件(www.r-project.org)进行数据统计分析,并获得频率分布柱形图;使用SAS 8.1软件对9个种子性状的相关性进行分析,并利用一般线性模型(Glm)计算各个性状的广义遗传力(H2)。
基于已经构建的分子遗传图谱[24],利用MAPQTL 4.0软件对自交亲和性及其种子相关的9个性状进行QTL分析。首先使用区间作图法(interval mapping,IM)找到可能存在QTL的图谱位置,以及与其紧密连锁的标记。根据模拟计算并确定全基因组范围内的LOD阈值。使用自动选择的协同因子(automatic cofactor selection)对于IM检测到的QTL紧密连锁的标记进行选择,在<0.02水平上显著的标记作为协同因子,用于多模型QTL作图(mutiple QTL model,MQM)。通过置换测验(1 000次重复)估算基因组范围内α=0.05水平上的LOD阈值,IM和MQM都以5 cM的步长扫描整个基因组。
2 结果
2.1 亲本及RIL群体表型变异
亲本R-O-18和L58在花期亲和指数(CIFT)、角果长度(SL)、角果喙长度(SBL)、角果长宽比(SL/SW)、角果喙长/角果长(SBL/SL)、每角果种子数(NSPS)、种子千粒重(TSW)、种子颜色(SC)8个性状的表型具有明显的差异(表1),利用这两个基因型构建的群体后代也存在较丰富的表型变异。例如,在RIL群体中,花期亲和指数均值为1.53,变异范围为0—7.07;角果长度均值为2.63,变异范围为1.24—4.37;每角果种子数均值为3,变异范围为0—13.27。虽然2个亲本结角率的性状差异不大,但后代单株结角率的表型分离明显,而且在角果长度、角果长宽比、角果喙长/角果长这些性状均出现明显的超亲分离。总之,RIL群体在9个性状上表现为连续变异,并且变异幅度较大,均呈正态分布或偏正态分布(图2),具有典型的数量性状遗传特点。
此外,对种子颜色之外的8个性状进行广义遗传力的分析,结果表明,这8个性状的广义遗传力均大于或者等于0.5,其中花期亲和指数的广义遗传力最高,达到0.67(表1),说明这些性状主要受基因调控,受到环境的影响相对较小。
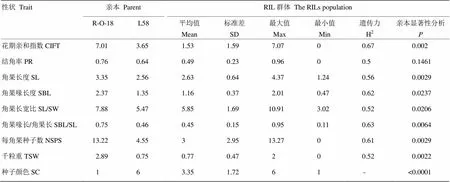
表1 亲本及RIL群体自交亲和性及种子相关性状的表型分布
2.2 自交亲和性、角果及种子相关性状间的相关性
在调查的自交亲和性、角果及种子相关性状中,各性状之间表现出不同程度的相关性(表2)。花期亲和指数与结角率、角果长度、角果喙长度、角果长宽比、每角果种子数、千粒重均存在极显著的正相关,其中花期亲和指数与每角果种子数间的相关系数最大,达到0.8487;与其他几个性状的相关性大小依次为:千粒重>结角率>角果喙长度>角果长度>角果长宽比。每角果种子数及千粒重均与结角率、角果长度、角果喙长度、角果长宽比呈现极显著正相关,而每角果种子数与千粒重之间也存在极显著的正相关(0.6477);结角率、角果长度、角果喙长度与角果长宽比之间呈现显著正相关;而角果喙长/角果长除了与角果长呈负相关,角果喙长度存在极显著正相关性外,与其他性状均不相关。结果表明,花期亲和指数与角果相关的性状与种子的产量有关,而角果喙长/角果长对种子的产量影响较小。

表2 L58×R-O-18 RIL群体自交亲和性、角果及种子性状之间的相关性分析
*(<0.05)代表差异显著;**(<0.01)代表差异极显著
* (value<0.05) represents significant difference; ** (value<0.01) show extremely significant difference
2.3 自交亲和性、角果及种子相关性状的QTL定位与分析
孟霖[24]利用该RIL群体已经构建了包括372个InDel标记的遗传连锁图谱。该连锁图覆盖总长度为968.9 cM,标记间平均遗传距离为2.60 cM,整个连锁图中的平均标记数为0.38 maker/cM。本研究基于该连锁图谱和表型调查数据,对自交亲和性、角果及种子相关性状进行了定位分析,共检测到12个QTL(表3),这些QTL分布在A02、A05、A06、A09和A10连锁群上(图3),可解释的表型变异为7.9%—42.1%;LOD值变异范围为3.2—13.79。

表3 L58×R-O-18 RIL群体自交亲和性、角果和种子性状的QTL

黑色区段为QTL区间 The black block shows the QTL confidence interval
检测到2个控制花期亲和指数的QTL,均位于A09连锁群上,其LOD值分别为4.55与3.6,单个QTL对花期亲和指数表型变异的贡献率分别为16.7%和13.1%,这两个位点的加性效应作用相反,分别为-0.77与0.6;检测到控制结角率、角果喙长度、角果喙的长度/角果长度、每角果种子数的QTL各1个(图3),分别位于A09、A09、A10和A09,LOD值范围为3.21—4.42,可解释13.4 %—14.9%的表型变异,这4个QTL 位点的加性效应均大于0,为增效位点;控制角果长度、角果长宽比与千粒重的QTL各1个,均位于A06连锁群上(图3),可以解释13.5%—15.9%的表型变异,这三个位点均为减效位点,加性效应值为-0.26、-0.66和-0.17。与种子颜色相关的QTL有3个,分别位于A02、A05和A09连锁群上,其LOD值变化范围为3.43—13.79,单个QTL解释7.9%、8.9%和42.1%的表型变异,其中位于A09连锁群上BrID11531与BrID11039区间的QTL对表型变异的解释率为42.1%,为主效QTL,其加性效应为1.22。
3 讨论
芸薹属植物属于自交不亲和植物,但也存在自交亲和的类型,前人对其自交亲和性开展了研究。何余堂与Schopfer等[7,25]研究发现,自交不亲和基因或的突变或缺失是导致白菜型油菜自交亲和的原因。Goring等[9]和Nasrallah等[10]对甘蓝及甘蓝型油菜自交亲和系的研究表明,自交不亲和基因发生缺失突变会使SRK蛋白丧失功能,进而导致自交亲和。这些结果表明,芸薹属植物自交亲和性主要是由自交不亲和位点(S位点)控制的。但本实验室前期对由2个自交亲和材料(R-O-18为高度自交亲和,L58自交亲和性较差)为亲本构建的RIL群体,进行蕾期套袋自交繁种时,发现个体间自交亲和性存在显著的分离,许多单株出现自交不亲和,根本无法结籽。推测自交亲和性除了受主要的S位点调控外,可能还受其他微效基因的影响。因此,本研究利用该RIL群体对自交亲和性进行了QTL分析,共定位到2个QTL,对表型变异的贡献率分别为16.7%和13.1%,与前人得到的调控自交亲和性的主位点(S位点)不同。这两个QTL可能为控制自交亲和性的非主效QTL位点,该结果不仅为进一步了解白菜自交亲和性的遗传基础、基因克隆奠定了基础,而且有助于分子标记辅助选择(MAS)选育自交亲和品种,从而加速优良自交系的选育,进一步利用杂种优势来提高白菜型油菜的产量和品质。
Bagheri等[3]利用与本研究相同的群体对角果及种子相关性状进行了QTL定位,分别检测到1个(位于A04连锁群)控制角果长度、4个(分别位于A08、A10、A04、A01连锁群)控制角果喙长度、2个(位于A09和A03连锁群)控制每角果种子数及2个(位于A09和A03连锁群)控制种子颜色的QTL。Lou等[16]利用白菜型油菜与小白菜、芜菁的DH群体定位了2个(分别位于A01与A07)控制角果长度、1个(分别位于A01与A07)控制角果宽度、3个(分别位于A05、A07和A09)控制角果喙长度、1个(位于A09)控制种子颜色的QTL。与上述两个QTL分析结果比较,本研究未检测到角果宽度QTL,角果长度QTL与上述两个QTL结果均不同,认为是新的QTL位点;在A09上检测到角果喙长度QTL与Lou等定位的其中1个QTL位于同一连锁群上,但与Bagheri等[3]的结果不一致;在A09连锁群上检测到1个控制每角果种子数的QTL(对表型变异的贡献率为13.4%),这与Bagheri等定位的一个效应较大的每角果种子数QTL(位于A09连锁群,贡献率为10%)处于同一连锁群区段,结果一致;关于种皮颜色,本研究在A09连锁群上定位到1个主效QTL,解释表型变异率为42%,这与Bagheri等[3]、Lou等[16]及Kebede等[23]检测到种子颜色QTL(贡献率分别为53.7%、61.65%和55%)所在的连锁群区段一致,但与原玉香等[6]在大白菜中定位的种子颜色主效QTL(位于A06上)不一致;定位在A05连锁群上的种皮颜色微效QTL与Bagheri等[3]的结果也一致。虽然本研究与Bagheri等[3]利用相同的群体,但得到的结果不完全一致,可能的原因是所用的遗传图谱不同、试验材料种植的环境不同等,这也说明角果长度及角果喙长度的性状受环境因素的影响较大,而种子颜色及每角果种子数受环境因素影响较小。相关性分析表明,结角率、花期亲和指数及每角果种子数性状之间呈显著正相关关系,而且这三个性状的QTL都定位到A09连锁群相同的区间;其中花期亲和指数与结角率和每角果种子数的相关性系数较高分别为0.5222和0.8487,而结角率与每角果种子数的相关性系数较低为0.37218,其原因可能是虽然角果膨大(结角),但由于部分胚珠没能正常发育成种子,使得结角率和每角果种子数之间的相关性并不是很高。角果长宽比和种子千粒重的呈显著正相关关系,且这两个性状的QTL定位到A06连锁群相同的区间,说明控制这些角果相关性状的QTL可能存在一因多效或紧密连锁效应。另外,花期亲和指数与产量构成相关的性状,包括结角率、角果长度、角果喙长度、角果长宽比、每角果种子数和千粒重均呈显著的正相关,说明自交亲和性与也与种子产量相关。因此,在育种上应该综合考虑各个角果、种子相关的性状来进行白菜型油菜产量和品质的遗传改良。
4 结论
定位2个控制自交亲和性的QTL,分别可解释13.1%和16.7%的表型变异;另外,定位7个产量构成相关性状的QTL,分别位于A06、A09和A10连锁群上,单个QTL可解释13.4%—17.7%的表型变异;3个种皮颜色的QTL,分别位于A02、A05和A09连锁群上,可解释7.9%、8.9%和42.1%的表型变异,其中位于A09连锁群的QTL为主效位点。
References:
[1] 唐国永, 徐青兰, 王瑞生, 李钧, 杜德志.白菜型大黄油菜自交亲和性初步研究.青海农林科技,2006,1:8-9.
Tang G Y, Xu Q L, Wang R S, Li J, Du D Z. Primary study on self-compatibility in Hahuang rape of., 2006, 1: 8-9. (in Chinese)
[2] 孙万仓, 范惠玲, 叶剑, 张亚宏, 刘雅利, 曾军, 王鹏.白菜型油菜自交亲和性变异分析.西北植物学报,2006,4:688-695.
Sun W C, Fan H L,Ye J, Zhang Y H, Liu Y L, Zeng J, Wang P. Variation ofL.self-compatibility., 2006, 4: 688-695. (in Chinese)
[3] Bagheri H, El-Soda M, van Oorschot I, Hanhart C, Bonnema G, Jansen-van den Bosch T, Mank R, Keurentjes J J, Meng L, Wu J. Genetic analysis of morphological traits in a new, versatile, rapid-cyclingLrecombinantinbred line population., 2012, 3: 183.
[4] 戚存扣, 陈玉卿, 张洁夫, 高冠军, 博寿仲, 惠飞虎.白菜型油菜自交亲和性分析及自交系基础群体筛选.中国油料,1997,4:11-13.
Qi C K, Chen Y Q, Zhang J F, Gao G J, Bo S Z, Hui F H. Analysis of self-compatibility and selecting of inbred lines population inL.., 1997, 4: 11-13.(in Chinese)
[5] 罗玉秀, 杜德志.白菜型油菜自交不亲和性状的遗传分析.青海大学学报,2008,6:6.
Luo Y X, Du D Z. Genetic analyses of self-incompatiblecharacterin., 2008, 6: 6.(in Chinese)
[6] 原玉香, 张晓伟, 蒋武生, 姚秋菊, 张强, 赵艳艳, 耿建峰.大白菜种皮颜色基因的QTL定位与分析. 华北农学报,2012, 27(1):74-78.
Yuan Y X, Zhang X W, Jiang W S, Yao Q J, Zhang Q, Zhao Y Y, Geng J F. QTL Mapping for seedcoat color in Chinese cabbage(L. ssp)., 2012, 27(1): 74-78.(in Chinese)
[7] Schopfer C R, Nasrallah M E, Nasrallah J B. The male determinant of self-incompatibility in.,1999, 286(5445):1697-1700.
[8] Watanabe M, Takasaki T, Toriyama K, Yamakawa S, Isogai A, Suzuki A, Hinata K. A high degree ofhomology exists between the protein encoded by SLG and the S receptor domain encoded by SRK in self-incompatibleL., 1994, 35(8): 1221-1229.
[9] Goring D R, Glavin T L, Schafer U, Rothstein S J. An S receptor kinase gene in self-incompatibilityhas a 1bp deletion., 1993, 5: 531-539.
[10] Nasrallah M E, Kandasamy M K, Nasrallah J B. A genetically define trans-acting locus regulatesS-locus function in., 1992, 2(4): 497-506.
[11] Fujimoto R, Sugimura T, Nishio T. Gene conver from SLG to SRK resulting in self-compatibility in., 2006, 580: 425-430.
[12] Kitashiba H, Liu P, Nishio T, Nasrallah J B, Nasrallah M E. Functional test ofself-incompatibility modifiers in., 2011, 108(44): 18173-18178.
[13] WATANABE M, ONO T, HATAKEYAMA K, TAKAYAMA S, ISOGAI A,HINATA K. Molecular characterization of SLG and S-related genes in a self-compatibleL. var. yellow sarson., 1997, 10(6): 332-340.
[14] Lim SH, Cho H, LeeJ, Cho YH, KimBD.Identification and classification of S haplotypes inby PCR-RFLP of the S locus glycoprotein (SLG) gene and the S locus receptor kinase (SRK) gene, 2002, 104: 1253-1262 .
[15] Robert SL, Allard S, Franklin M T, Trick M. Sequence and expression of endogenous S-locus glycoprotein genes in self-compatible., 1994, 242: 209-216.
[16] Lou P, Zhao J, Kim J S, Shen S, Del Carpio D P, Song X, Jin M, Vreugdenhil D, Wang X, Koornneef M. Quantitative trait loci for flowering time and morphological traits in multiple populations ofL.., 2007, 58(14):4005-4016.
[17] Zhang L, Li S, Chen L, Yang G. Identification and mapping of a major dominant quantitative trait locus controlling seeds per silique as a single Mendelian factor inL.., 2012, 125(4): 695-705.
[18] Yang P, Shu C, Chen L, Xu J, Wu J, Liu K. Identification of a major QTL for silique length and seed weight in oilseed rape(L.)., 2012, 125(2): 285-296.
[19] 张凤启, 程晓辉, 刘越英, 童超波, 董彩华, 于景印, 黄军艳, 刘胜毅. 甘蓝型油菜分枝数QTL定位及其候选基因预测. 中国油料作物学报,2015,1:15-20.
Zhang F Q, Cheng X H, Liu Y Y, Tong C B, Dong C H, Yu J Y, Huang J Y, Liu S Y. QTL mapping of branch inand candidate gene prediction., 2015, 1: 15-20.(in Chinese)
[20] 孙美玉, 华玮, 刘静, 王新发, 刘贵华, 王汉中. 甘蓝型油菜主花序有效角果数QTL定位. 中国油料作物学报,2013,1:1-7.
Sun M Y, Hua W, Liu J, Wang X F, Liu G H, Wang H Z. QTLs for effective silique number of main in florescence on rapeseed (L.)., 2013, 1: 1-7. (in Chinese)
[21] Zhang L, Yang G, Liu P, Hong D, Li S, He Q. Genetic and correlation analysis of silique-traits inLby quantitative trait locus mapping., 2011, 122(1): 21-31.
[22] 王玉刚, 修文超, 沈宝宇, 周泽峰, 古君伶. 白菜和白菜型油菜角果相关性状遗传分析. 植物遗传资源学报,2013, 14(3):547-552.
Wang Y G, Xiu W C, Shen B Y, Zhou Z F, Gu J L. Genetic analysis of pod correlated traits in., 2013, 14(3): 547-552.(in Chinese)
[23] Kebede B, Cheema K, reenshields D L, Li C, Selvaraj G, Rahman H. Construction of genetic linkage map and mapping of QTL for seed color in.., 2012, 55(12): 813-823.
[24] 孟霖. 白菜类作物氮利用效率的QTLs分析[D]. 昆明: 云南农业大学, 2012.
Meng L. QTL analysis of nitrogen use efficiency in[D]. Kunming: Yunnan Agricultural University,2012. (in Chinese)
[25] 何余堂, 龙卫华, 胡进平, 傅廷栋, 李殿荣, 陈宝元, 涂金星.白菜型油菜角果多室性状的遗传及解剖学研究. 中国油料作物学报,2003, 25(1): 1-4.
He Y T, Long W H, Hu J P, Fu Y D, Li D R, Chen B Y, Tu J X. Anatomic and genetic studies on multicapsular character inL.., 2003, 25(1): 1-4.(in Chinese)
(责任编辑 李莉)
QTL Mapping of Self-Compatibility, Silique and Seeds-associated Traits in
SUN Qian-qian1,2, WU Jian2, CHENG Feng2, WANG Xiao-wu2, LIANG Jian-li2, SHEN Shu-xing1
(1College of Horticulture, Agricultural University of Hebei/Key Laboratory for Vegetable Germplasm Enhancement and Utilization of Hebei/Collaborative Innovation Center of Vegetable Industry in Hebei, Baoding 071000, Hebei;2Institute of Vegetables and Flowers, Chinese Academy of Agricultural Sciences, Beijing 100081)
【Objective】is an important oil crop in the world. Higher seed yield with good quality is one of the important agronomic traits. Due to the self-incompatibility characteristic of most, it is difficult to purify breeding materials and realize maximum yield of this plant, since the seed yield of this crop is influenced by the silique related traits, such as the number of pods, number of seed per silique. Besides, the seed quality is closely related to seed color. The aim of this research is providing an important genetic basis for breeding self-compatibility varieties or inbred lines ofand genetic modification of yield with quality in this crop.【Method】The Recombinant Inbred Lines (RILs) population derived from the cross between genotypes R-O-18 with well self-compatibility and L58 with weak self-compatibility consisting of 117 lines was used to conduct QTL analysis. Based on previous linkage map generated by 372 InDel markers, the Interval Mapping (IM) method was used to survey a total of nine traits as follows: compatible index of flowering time (CIFT), pods rate (PR), silique length (SL), silique beak length (SBL), silique length/silique width (SL/SW), silique beak length/silique length (SBL/SL), number of seed per silique (NSPS), thousand seed weight (TSW) and seed color (SC).【Result】RILs population in the 9 self-compatibility, siliques and seeds-related traits showed continuous variation and variation margin, showed normal or partial normal distribution, with typical quantitative genetic characteristics. CIFT showed an extremely significant positive correlation with PR, SL, SBL, SL/SW, NSPS, and TSW, where the correlation coefficient between CIFT and NSPS was the largest, reaching 0.8487; and several other traits in order of size (high to low): TSW, PR, SBL, SL, and SL/SW. NSPS and TSW presents a very significant positive correlation with PR, SL, SBL, SL/SW, NSPS also had a significant positive correlation with TSW (0.6477). The PR, SL and SBL, SL/SW also showed a significant positive correlation. SBL/SL showed a negative correlation and a significant positive correlation with SBL, but not correlated with other traits. A total of 12 QTL loci were detected and the major ones were located on chromosome A09, and totally 10 QTLs explained phenotypic variation of more than 10%. Two QTLs controlling CIFT were located on chromosome A09, accounting for the phenotype variation 13.1% and 16.7%, respectively. Six QTLs related to silique were located on A06, A09 and A10, respectively, explaining phenotypic variance of 13.4%-17.7%. Furthermore, four seed-associated QTLs were located on chromosome A02, A05, A06 and A09, respectively, with QTL effects varied from 7.9% to 42.1%, of which the QTL located on A09 was the major QTL for SC, as it accounted for phenotypic variance of 42.1% and it had additive effect value of 1.22. 【Conclusion】A total of 12 QTLs which control self-compatibility (2), siliques (6) and seeds-related traits (4) were detected. For the two self-compatibility QTLs, they are different with the S locus identified previously, which indicated that the two loci detected in this study may be the non-major QTLs. Additionally, one QTL of the 4 seeds-related QTLs is the major QTL of this trait.
; self-compatibility; silique; seeds-associated traits; QTL
2016-02-01;接受日期:2016-05-13
国家“863”计划(2012AA100101)、国家“973”计划(2012CB113906)、国家自然科学基金(31272179,31301784)、河北省科技支撑计划(16226304D-2)
孙倩倩,E-mail:15200088028@163.com。通信作者申书兴,E-mail:。通信作者梁建丽,E-mail:liangjianli@caas.cn
