水稻根基粗适宜取样量及其与抗倒伏性状的相关分析
2016-07-23王小虎方云霞张栋
王小虎++方云霞++张栋

摘要:利用典型的籼粳交(窄叶青8号/京系17)的F1代花培加倍单倍体(DH)分离群体为材料,通过水培法在齐穗期考查了与抗倒伏相关根基粗等性状。根基粗在DH群体中呈明显的连续性变异分布。根基粗总平均数与10、20、30、40、50、60、70、80、90、100个最粗根基粗平均数均存在极显著正相关,且根基粗总平均数(Y)与10~100个最粗根基粗平均数(X)两者关系符合直线回归方程Y=a+bX。根基粗总平均数与30、40个最粗根基粗平均数的相关系数最大。根基粗与水稻抗倒伏相关的地下部根系性状以及地上部性状的相关分析表明,根基粗与总根数、最大根长、根干物质量、根冠比等根系性状之间存在着极显著正相关;除分蘖数外,根基粗与地上部株高、株周长、单蘖直径、地上部干物质量之间也存在着极显著正相关。根基粗与地上部性状的多元回归分析表明,根基粗只与单蘖直径、地上部干物质量成显著线性关系,这说明单蘖直径和地上部干物质量对根基粗的影响较大。本研究旨在寻求水稻根基粗的最适调查方法,为水稻根系育种提供理论参考。
关键词:水稻;根基粗;取样量;抗倒伏性状;相关分析
中图分类号: S511.03文献标志码: A文章编号:1002-1302(2016)05-0085-05
根系是水稻重要的地下组织器官,具有固定植株、吸收水分和养分、合成内源激素等众多功能[1-2]。此外,水稻根系还是地上与地下各部分物质及信息交换的系统之一[3]。
根基粗是水稻根系统的重要性状之一,在水稻根系的遗传[4-5]、抗旱[6-8]、抗倒伏[9-10]等诸多研究中是必不可少的研究因子。其他的根系性状如总根数、根长、根干物质量、根冠比等数据调查相对简单,在实验室均可操作。而根基粗数据调查相对复杂,费时又费力[7,11],同时随着测量群体的增大,出现误差的概率相应增加。目前,公认调查根基粗较为准确、快速和便捷的方法是利用WinRHIZOLA6400XL根系专用大幅面透视扫描仪对根系进行透视扫描,然后用根系专用 WinRHIZOPro 软件分析扫描图片获取数据。这种方法虽可获得根系准确、完整的表观数据,但也有其缺点,应用此方法得到的数据不是直接测量的表型数据,而是通过处理扫描后的照片得出的数据,与性状的实际值之间还存在着一定误差[12-13]。
本研究利用水培法获得DH群体各株系完整的根系统[14-15],利用激光测量仪(型号为KEYENCE LS-7000,测量范围为0~30 mm)测量根基粗,同时考查了根基粗与抗倒伏相关的总根数、最大根长、根干物质量、根冠比等地下部性状以及与抗倒伏相关的分蘖数、株高、株周长、单蘖直径、地上部干物质量等地上部性状之间的相关性,以获得根基粗研究时最适取样样本量,为水稻根系育种研究提供参考依据。
1材料与方法
1.1试验材料
利用典型的籼稻品种窄叶青8号(ZYQ8)与粳稻品种京系17 (JX17)杂种F1代花药培养,获得127个稳定株系组成的DH群体。
1.2试验方法
1.2.1材料种植DH群体及双亲种植于中国水稻研究所试验基地,6叶期时移入聚苯乙烯泡沫板中,每板规格为150 cm×100 cm×5 cm,共35穴,行株距为20 cm×15 cm,种植孔径4.5 cm,以海绵为固定基质。采用水稻固体混合肥料,含大量元素N、P、K和各种微量元素,折算纯氮 180 kg/hm2。根据历年调查资料,每块板按照DH群体的株高不同由低到高排列种植,每行为1株系,共7株。所有材料均种植于室外的大水池中。
1.2.3性状考察齐穗7 d后,每株系取3株,用激光测量仪(型号为KEYENCE LS-7000,测量范围为0~30 mm)测量离根基部2 cm处的根粗;用直尺量取最长根长度;记录根总数、根和地上部干物质量、根冠比、分蘖数;测量计算株高、株周长及单蘖直径等数据,其中株周长为贴地稻基部的周长。采用Excel 2003和SPSS 17.0软件对数据进行统计分析(相关和回归分析的样本量n均为DH株系数)。
2结果与分析
2.1根基粗及其他根系性状在双亲和DH群体中的表现
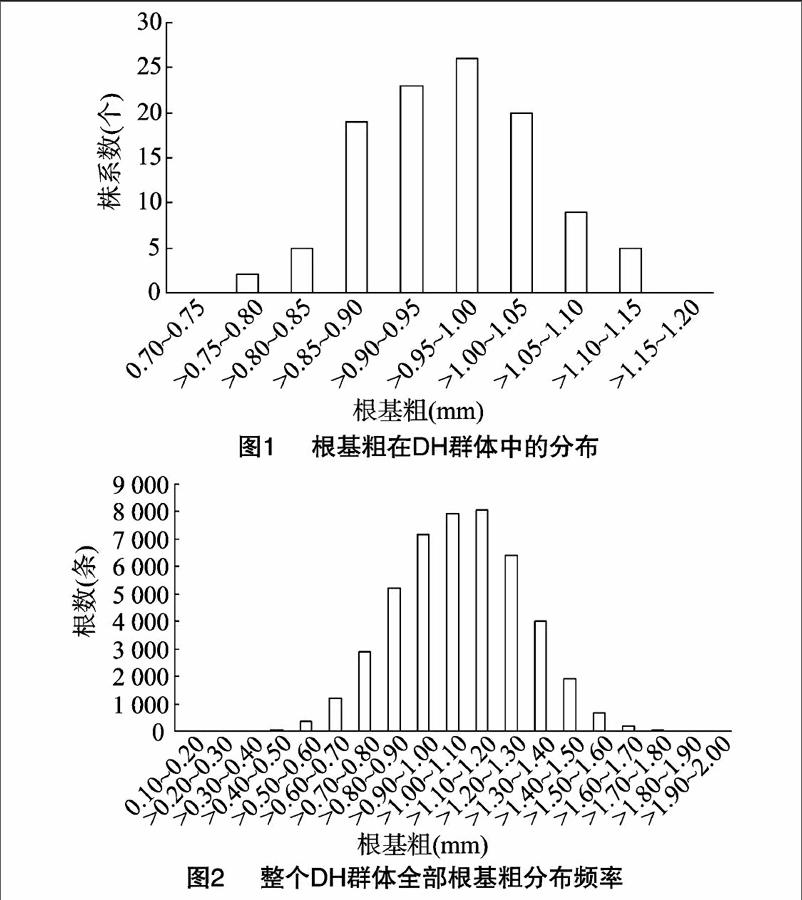
从图1可以看出,根基粗在DH群体中表现出连续性的正态分布,表明水稻根基粗是由数量性状基因控制的。整个群体全部根基粗也表现正态分布(图2),其中,在整个群体中根基粗在1.00~1.20 mm区段分布较多,而在0.10~0.60 mm 和1.60~1.90 mm区段分布最少,这表明整个DH群体根基粗的大小大部分集中在1.00~1.20 mm这个区段,特别细或者特别粗的根系较少。从表1可以看出,亲本间除了根基粗、最大根长差异显著或极显著外,其他3个性状差异均不显著。在考查DH株系时,5个性状均表现出超亲现象,除根干物质量的偏度和峰度以及总根数的峰度大于1外,其余均小于1。
2.2根基粗总平均值与10~100个最粗根基粗的平均值相关分析
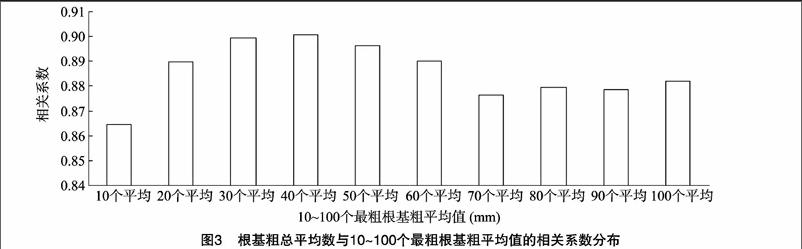
相关分析表明,根基粗的总平均数与10~100个最粗根基粗的平均值均存在极显著的正相关,相关系数变异范围为0.864 7~0.900 7。从图3可以看出,根基粗总平均值与 10~40个最粗根基粗平均值的相关系数呈逐渐增大的趋势,与50~70个最粗根基粗平均值的相关系数呈下降的趋势,与80~100个最粗根基粗平均值的相关系数又呈逐渐增大的趋势。根基粗总平均值与40个最粗根基粗平均值的相关系数最大,与10个最粗根基粗平均值相关系数最小,相关系数大小排列顺序为40个平均>30个平均>50个平均>60个平均>20个平均>100个平均>80个平均>90个平均>70个平均>10个平均。
2.3根基粗总平均值与10~100个最粗根基粗平均值的回归关系分析
从图4可以看出,根基粗总平均值与10~100个(只列出了10个、30个、40个、100个)最粗根基粗平均值在散点图上呈明显的线性关系,其数量关系可用一个线性方程来表示[16]。如表2所示,根基粗总平均值与10~100个最粗根基粗平均值线性回归方程的估计标准误(s)很小,而s是回归精确度的度量,s愈小,由回归方程估计y的精确性愈高。且F值远大于F0.01值,故根基粗总平均值与10~100个最粗根基粗平均值呈直线回归关系。从表2中回归截距a置信区间来看,30、40个最粗根基粗平均值在0.118 5~0261 4 mm、0140 0~0.278 3 mm之间时,这一推断的可靠度为95%。表明可以通过测量30~40个最粗的根基粗平均值来推算根基粗的总平均值。
从表3可以看出,根基粗与总根数、最大根长、根干物质量、根冠比等根系性状分别呈极显著正相关,与株高、株周长、单蘖直径、地上部干物质量等地上部性状分别呈极显著的正相关。这表明株系的根基部越粗时,根越长,株高相对越高,株周长、单蘖直径越粗。说明根基粗是水稻根系统中重要性状之一。根系性状相互之间也都呈极显著正相关。除根冠比外,其他根系性状与株周长、单蘖直径、地上部干物质量等地上部性状都呈极显著正相关。分蘖数除与总根数呈极显著正相关外,与其他根系性状相关都不显著;株高除与根基粗呈极显著相关外,与其他根系性状相关不显著。2.5根基粗与地上部性状的多元回归分析
对水稻根基粗(Y1)与水稻地上部抗倒伏相关分蘖数(X1)、株高(X2)、株周长(X3)、单蘖直径(X4)、地上部干物质量(X5)等5个农艺性状进行逐步多元回归分析。表4结果表明,根基粗只与单蘖直径、地上部干物质量有显著的线性关系。回归方程Y1=0.792+0.118X4+0.001 78X5表明单蘖直茎、地上部干物质量分别每增加1个单位,根基粗将分别增加0.118个单位和0.001 78个单位,两者分别解释了6.81%和6.30%的根基粗的变异。
本研究还对总根数(Y2)、最大根长(Y3)、根干物质量(Y4)、根冠比(Y5)等4个地下部根系性状与地上部性状进行了多元回归分析。表4结果表明,除分蘖数对4个根系性状均没有显著线性效应外,株高、株周长、单蘖直径、地上部干物质量对4个根系性状均存在显著线性效应。多元回归方程表明,株高每增加1个单位,总根数、根干物质量分别减少0798、0.011 4个单位,而根冠比则增加0.000 404个单位,分别解释了4.24%的总根数、5.66%的根干物质量和942%的根冠比变异。株周长只与总根数存在显著线性效应,株周长每增加1个单位,总根数则增加14.385个单位,解释了1354%的总根数变异。单蘖直径每增加1个单位,最大根长和根冠比分别增加9.528、0.001 02个单位,分别解释了676%的最大根长和6.00%的根冠比变异。地上部干物质量每增加1个单位,总根数、最大根长和根干物质量分别增加3.544、0.204、0.076 4个单位,分别解释了21.25%的总根数、12.04%的最大根长和61.00%的根干物质量变异。
多元回归方程较好地反映了水稻抗倒伏相关的地下部根系性状和地上部农艺性状的关系,这与相关分析结果基本一致。但是总根数、根干物质量、根冠比与株高无显著关系,通过多元回归分析却发现它们之间的关联。
3结论与讨论
利用水培法对DH群体进行根系的相关研究,其优点是可以保证根系的完整性,并且生长条件容易控制,可很好地模拟水稻在自然条件下的生长状态,取得与水田相仿甚至更高的产量[17]。最重要的是,利用水培法获得的水稻根系性状与大田水稻根系性状能够表现较好的一致性[15]。因此,水培法是研究水稻根系的一种简便易行而且较可靠的方法。
本研究对DH群体齐穗后7 d的根系性状进行取样分析,该时期的水稻根系能比较完整地反映水稻根形态特征[4]。相关统计分析表明,DH群体中的根基粗性状呈近似的正态分布,并且根基粗总平均值与10~100个最粗根基粗平均值分别存在着极显著正相关,且有明显的线性回归关系,回归截距和回归系数变化范围都很小。根基粗总平均值与30个、40个最粗根基粗平均值相关系数最大,这表明在研究根基粗时,考查30个或者40个最粗的根系基粗就能反映出整个株系根基粗。
倒伏已成为水稻高产稳产的重要限制因素之一[18]。根据倒伏发生的位置不同,水稻倒伏可分为茎秆倒伏和根倒伏2种。而在生产实践中发生的水稻倒伏,大部分兼有根倒伏和茎秆倒伏的综合性特征[19]。根倒伏即发生在稻株根际的全株倒伏,主要是由于水稻根系入土较浅、下层根系发育差所致[20]。茎秆倒伏由于较普遍且比较直观,所以研究报道较多[21-24]。然而关于根倒伏的研究却鲜有报道[20,25],主要原因是由于根系处于地下,挖起时容易损失其固有形态,损伤较大,研究相对困难。随着抛秧技术[26]和直播稻技术[27]的推广,根系倒伏的风险加大,人们对水稻抗倒伏性研究提出了新的要求,因而研究水稻根系与抗倒伏关系势在必行。本研究把水稻抗倒伏相关的地下部根系性状与抗倒伏相关的地上部性状结合起来分析,结果表明,地下部根系性状与地上部性状具有联动关系,除根冠比外,其他根系性状与株周长、单蘖直径、地上部干物质量等地上部性状都呈极显著正相关。其中,根基粗、最大根长、总根数与株周长、单蘖直径分别呈极显著的正相关。而茎秆越粗,植株的抗倒能力相对越强[28]。因此,根基粗、最大根长、总根数可以作为水稻抗倒伏育种的选择目标,同时这三者也是抗旱育种的选择目标[7]。研究还表明,地下部根系性状与地上部性状存在多元回归关系,回归方程较好地反映了地上部性状对地下部根系性状所起的作用。但是总根数、根干物质量、根冠比与株高无显著关系,通过多元回归分析却发现它们之间的关联,说明株高对地下部根系性状具有一定的影响。
参考文献:
[1]黄英金,石庆华,李木英,等. 水稻根系性状的遗传研究[J]. 江西农业大学学报,1995,17(4):368-374.
[2]石庆华,黄英金,李木英,等. 水稻根系性状与地上部的相关及根系性状的遗传研究[J]. 中国农业科学,1997,30(4):62-68.
[3]廖荣伟,刘晶淼. 作物根系形态观测方法研究进展讨论[J]. 气象科技,2008,36(4):429-435.
[4]吴伟明. 水稻根系性状的遗传及基因定位[D]. 北京:中国农业科学院,2006.
[5]梁永书. 超级稻协优9308衍生群体根系的遗传研究[D]. 北京:中国农业科学院,2011.
[6]穆平,李自超,李春平,等. 水、旱稻根系性状与抗旱性相关分析及其QTL定位[J]. 科学通报,2003,48(20):2162-2169.
[7]凌祖铭,李自超,余荣,等. 水、陆稻根部性状的研究[J]. 中国农业大学学报,2002,7(3):7-11.
[8]Li J Z,Han Y C,Liu L,et al. qRT9,a quantitative trait locus controlling root thickness and root length in upland rice[J]. Journal of Experimental Botany,2015,66(9):2723-2732.
[9]Ekanayake I J,Garrity D P,Toole J O. Influence of deep root density on root pulling resistance in rice[J]. Crop Science,1986,26(6):1181-1186.
[10]Jong G W,Hirahara Y,Yoshida T,et al. Selection of rice lines using SPGP seedling method for direct seeding[J]. Plant Production Science,1998,1(4):280-285.
[11]孙传清,张文绪. 水稻根系性状和叶片水势的遗传及其相关研究[J]. 中国农业科学,1995,28(1):42-48.
[12]王会民. 超级稻协优9308根系相关性状QTLs的精细定位[D]. 沈阳:沈阳农业大学,2013.
[13]梁永书,占小登,高志强,等. 超级稻协优9308衍生群体根系与地上部重要农艺性状的相关性[J]. 作物学报,2011,37(10):1711-1723.
[14]宋祥甫,应火冬,朱敏,等. 自然水域无土栽培水稻的研究[J]. 中国农业科学,1991,14(4):8-13,96.
[15]吴伟明,宋祥甫,邹国燕. 利用水上栽培方法研究水稻根系[J]. 中国水稻科学,2000,14(3):189-192.
[16]莫惠栋. 农业试验统计[M]. 上海:上海科学技术出版社,1984.
[17]宋祥甫,吴伟明,应火冬,等. 自然水域无土栽培水稻的生态适应性研究[J]. 中国水稻科学,1996,10(4):227-234.
[18]赵黎明,顾春梅,陈淑洁,等. 水稻倒伏研究及其影响因素分析[J]. 北方水稻,2009,39(4):66-70.
[19]张喜娟,李红娇,李伟娟,等. 北方直立穗型粳稻抗倒性的研究[J]. 中国农业科学,2009,42(7):2305-2313.
[20]Terashima K. Eco-physiological study of root lodging tolerance in direct-seeded rice (Oryza sativa) cultivars[J]. Japan Agricultural Research Quarterly,1997,31(3):155-162.
[21]胡江,藤本宽,郭龙彪,等. 水稻抗倒力及相关抗倒伏性状的QTL分析[J]. 中国水稻科学,2008,22(2):211-214.
[22]郭保卫,朱大伟,许轲,等. 有序摆抛栽对超级稻植株抗倒伏能力的影响[J]. 中国水稻科学,2015,29(1):45-55.
[23]张忠旭,陈温福,杨振玉,等. 水稻抗倒伏能力与茎秆物理性状的关系及其对产量的影响[J]. 沈阳农业大学学报,1999,34(2):81-85.
[24]Kashiwagi T,Togawa E,Hirotsu N,et al. Improvement of lodging resistance with QTLs for stem diameter in rice (Oryza sativa L.)[J]. TAG,2008,117(5):749-757.
[25]Terashima K,Akita S,Sakai N. Eco-physiological characteristics related with lodging tolerance of rice in direct sowing cultivation.Ⅲ.Relationship between the characteristics of root distribution in the soil and lodging tolerance[J]. Crop Science Society of Japan,1995,64(2):243-250.
[26]杨祥田,罗三镯,吴晓华.水稻抛秧栽培对其抗倒伏能力的影响[J]. 浙江农业学报,1999,11(3):151.
[27]许轲,唐磊,郭保卫,等. 不同水直播方式水稻植株抗倒特性研究[J]. 华北农学报,2014,29(6):226-232.
[28]饶玉春,李跃,董国军,等. 水稻抗倒伏研究进展[J]. 中国稻米,2009(6):15-19.
[29]吴伟明,程式华. 水稻根系育种的意义与前景[J]. 中国水稻科学,2005,19(2):174-180.
[30]魏磊,董华林,武晓智,等. 水稻根系育种研究进展[J]. 农业科学与技术,2015,16(4):675-678.陈锐浩,程凤娴,官利兰,等. 随水施肥技术在水稻上的应用研究[J]. 江苏农业科学,2016,44(5):90-93.
