分蘖期干旱胁迫及复水对水稻抗氧化系统及脯氨酸影响
2016-03-22赵宏伟王新鹏于美芳沙汉景于天聪邹德堂东北农业大学农学院哈尔滨150030
赵宏伟,王新鹏,于美芳,沙汉景,贾 琰,于天聪,邹德堂(东北农业大学农学院,哈尔滨 150030)
分蘖期干旱胁迫及复水对水稻抗氧化系统及脯氨酸影响
赵宏伟,王新鹏,于美芳,沙汉景,贾琰,于天聪,邹德堂
(东北农业大学农学院,哈尔滨150030)
摘要:为明确水稻耐旱机制,试验以松粳6号(干旱敏感型)和东农425(耐旱型)为材料,采用盆栽法,于分蘖期开始干旱胁迫(土壤水势分别为0、-10、-25、-40 kPa),处理21 d后复水,研究分蘖期干旱胁迫及复水对两个不同耐旱型水稻品种抗氧化系统影响。结果表明,随胁迫时间延长,干旱胁迫下两个品种水稻超氧阴离子(O2-)、脯氨酸含量,超氧化物歧化酶(SOD)、过氧化物酶(POD)、抗坏血酸过氧化物酶(APX)活性均随胁迫时间上升,且干旱胁迫越严重(水势越低),上升幅度越大;除APX外,其余各项指标复水后均快速下降。同时,与松粳6号相比,耐旱品种东农425干旱处理后O2-含量较低,而SOD、POD、APX活性和脯氨酸含量较高,复水后东农425抗氧化酶活性及脯氨酸含量可在较高水平维持更长时间。APX对干旱胁迫响应时间早于POD。
关键词:水稻;干旱胁迫;抗氧化系统;脯氨酸
赵宏伟,王新鹏,于美芳,等.分蘖期干旱胁迫及复水对水稻抗氧化系统及脯氨酸影响[J].东北农业大学学报, 2016, 47(2): 1-7.
Zhao Hongwei, Wang Xinpeng, Yu Meifang, et al. Effect of drought stress and rewatering on antioxidant system and proline in rice during tillering stage[J]. Journal of Northeast Agricultural University, 2016, 47(2): 1-7. (in Chinese with English abstract)
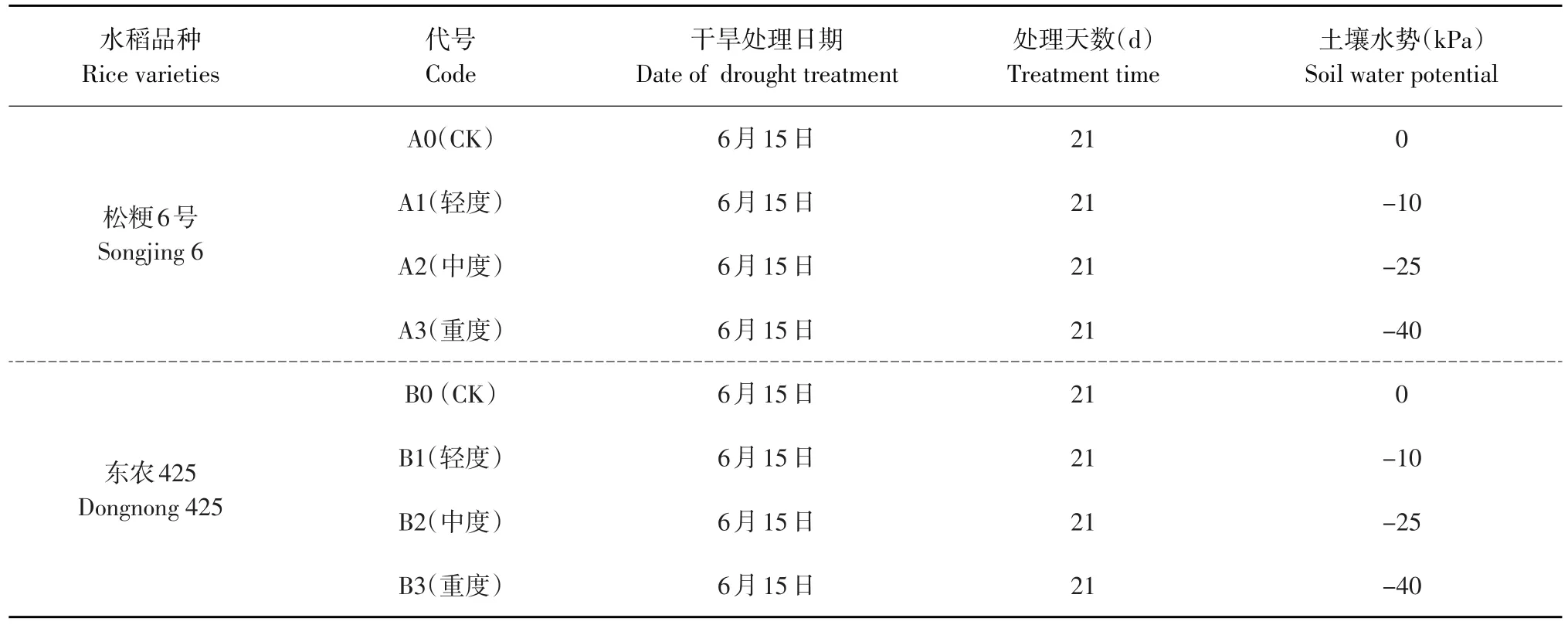
表1试验设计及处理代号Table 1 Test design and code
2结果与分析
2.1干旱胁迫及复水对水稻超氧阴离子含量影响
如图1所示,干旱胁迫期间,随土壤水势降低,两个水稻品种叶片内O2-水平逐渐增加。干旱胁迫初期,各处理植株叶片内O2-水平变化不大,但在处理18 d后O2-迅速升高,并在干旱处理第21天达到最大值。干旱胁迫初期松粳6号各处理间O2-含量差异明显,干旱胁迫第18天后,中度干旱胁迫处理下O2-水平逐渐接近重度胁迫。如表2所示,干旱处理第21天时,松粳6号在-25和-40 kPa下叶片内O2-水平分别是胁迫初期的3.74和3.96倍,而东农425仅是胁迫初期的3.14和3.41倍。此时东农425的O2-含量仅相当于松粳6号的83.4%、84.5%。说明耐旱型水稻品种在干旱胁迫期间能维持较低O2-水平,而干旱敏感型水稻品种随胁迫时间和胁迫程度加剧,对O2-清除能力逐渐降低。
复水后两个品种O2-含量均迅速下降,东农425复水后,各处理O2-含量均表现相同下降速度。中度与重度干旱处理下松粳6号下降速度快于东农425,但轻度干旱处理下趋势相反。表明干旱胁迫解除后,干旱敏感型水稻品种清除活性氧能力得到恢复,而耐旱型水稻品种清除活性氧能力能够维持相对稳定水平。
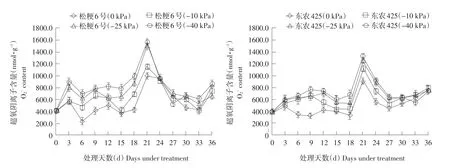
图1干旱胁迫下水稻O2-含量变化Fig. 1 Change of rice O2-content under drought stress
2.2干旱胁迫及复水对水稻抗氧化酶系统影响
水分胁迫下,水稻启动抗氧化酶系统,通过SOD清除因碳同化受阻在强光下形成的超氧阴离子,产生的H2O2可由POD进一步解毒形成H2O,减轻自由基对植株伤害。APX活性升高,可使O2-产生速率显著下降,脂质过氧化作用减弱。如图2所示,随干旱处理时间增加,SOD活性逐渐升高,中度与重度干旱处理第18天开始叶片SOD活性明显高于其他处理,21 d达到峰值。处理9~15 d POD活性甚至出现下降趋势,但在处理18 d后迅速升高,并在第21天达到最大值。APX活性在处理后第3天迅速提高,松粳6号与东农425分别于第9、12天迅速达到最大值。达到峰值后,两品种APX活性迅速下降,处理期间东农425的APX活性均高于当日松粳6号。如表2所示,相比松粳6号,东农425叶片内APX活性提升显著高于SOD与POD。APX活性达到峰值时,在轻度、中度、重度干旱下,东农425叶片内APX活性分别是胁迫初期的9.06、11.26和12.19倍。而此时松粳6号APX活性是胁迫初期的7.87、9.21和9.53倍。东农425轻度、中度、重度干旱胁迫处理下APX活性较松粳6号分别提高15.6%、22.3%、24.5%。表明在干旱初期,耐旱品种东农425较松粳6号有更大幅度提升APX酶活性的能力,减轻由于O2-积累造成的脂质过氧化。
复水后,东农425 SOD活性表现出相对缓慢的下降速度,表明复水后东农425能够在较长时间内维持较高的SOD活性。复水后各处理POD活性迅速下降,复水后第6天达到最低点,而后活性有提升。复水后APX表现出与POD类似变化,可能与气温升高有关。
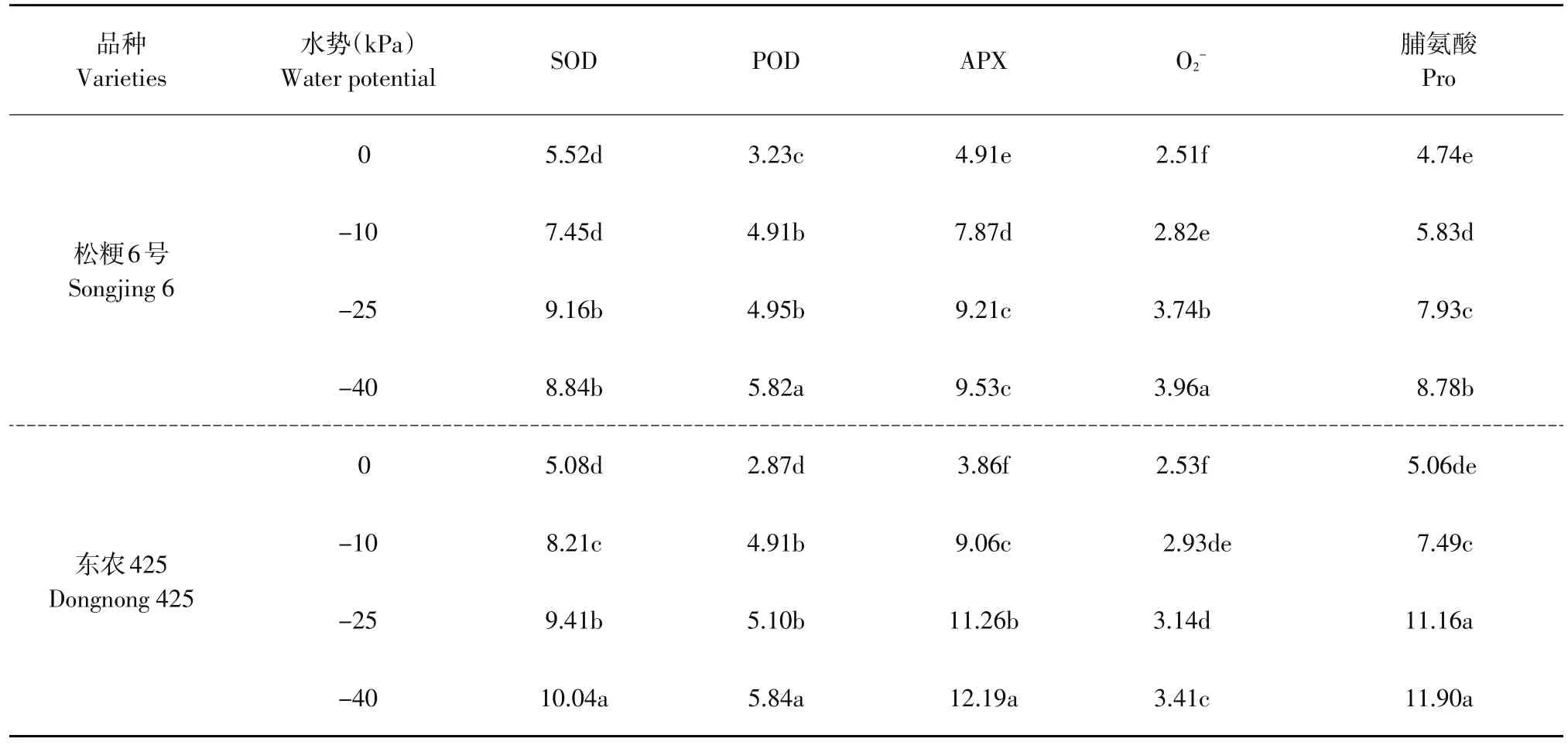
表2干旱胁迫下SOD、POD、APX、O2-、脯氨酸相对改变量Table 2 Relative change quantity of SOD, POD, APX, O2-, Pro under the drought stress
结果表明,东农425干旱胁迫下抗氧化酶尤其是APX活性显著提升,使O2-含量增加减缓,降低干旱胁迫对植物损害,对于增强抗旱性具有重要意义。
2.3干旱胁迫及复水对水稻脯氨酸含量影响
脯氨酸可保持细胞与环境渗透平衡,防止植物水分散失,可直接影响作物蛋白质稳定性[17]。如图3所示,随处理时间延长,两个水稻品种脯氨酸不断积累,在处理第21天达到峰值;东农425脯氨酸积累速率及脯氨酸含量均显著高于松粳6号。如表2所示,在轻度、中度、重度干旱下,东农425脯氨酸含量分别是胁迫初期的7.49、11.16和11.9倍;此时松粳6号分别是胁迫初期的5.83、7.93和8.78倍。此时东农425脯氨酸含量较松粳6号提高39.3%(-10 kPa)、48.2%(-25 kPa)、42.1%(-40 kPa)。表明干旱胁迫下东农425具有反应迅速、大量积累的特点。
复水后,东农425各处理均表现脯氨酸下降速率快。说明耐旱品种东农425具有迅速调节能力,干旱恢复能力较松粳6号更强。两品种脯氨酸含量在降至最低点后均略有回升。
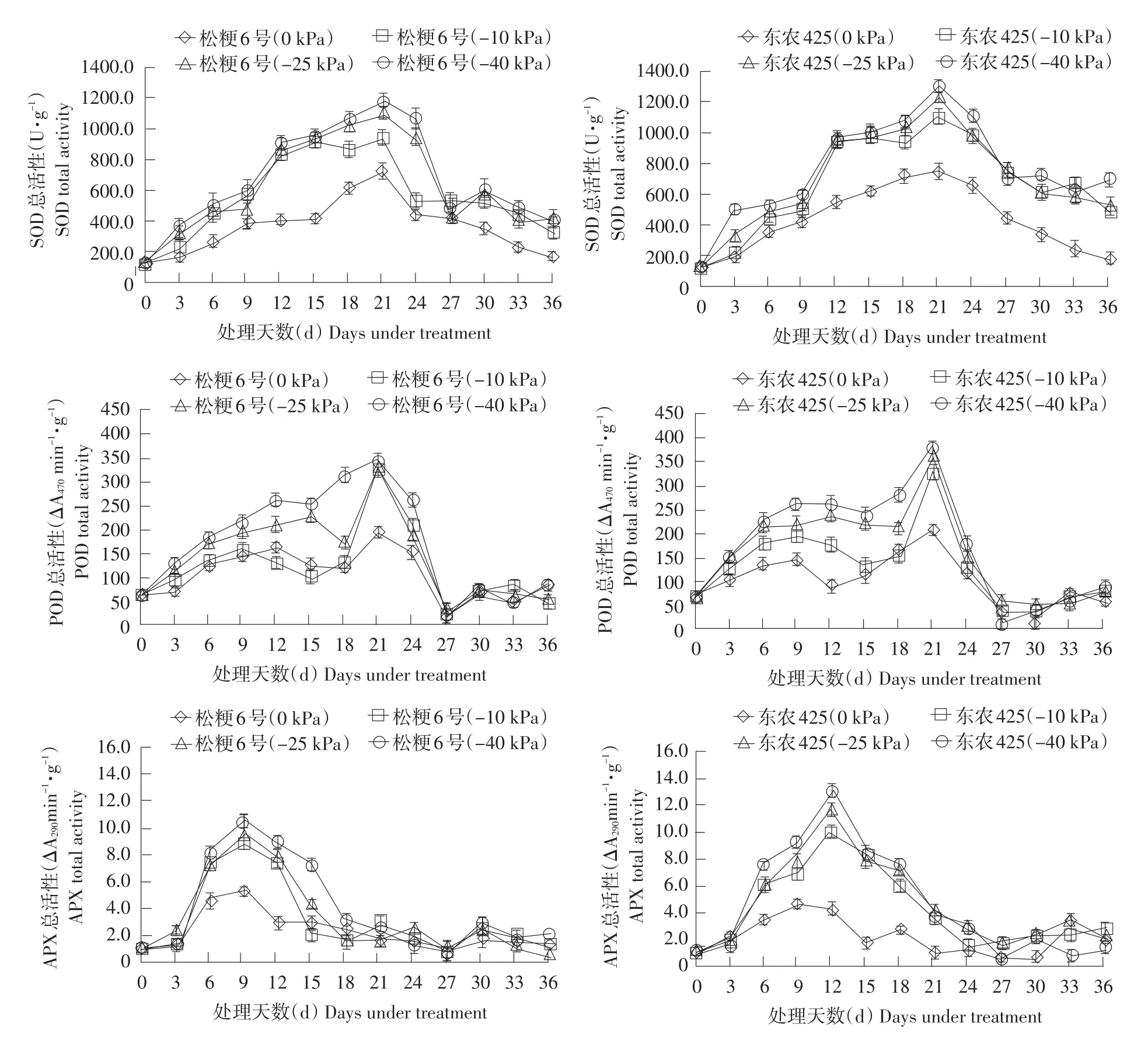
图2干旱胁迫下水稻SOD、POD、APX活性变化Fig. 2 Change of rice SOD, POD, APX activities under drought stress
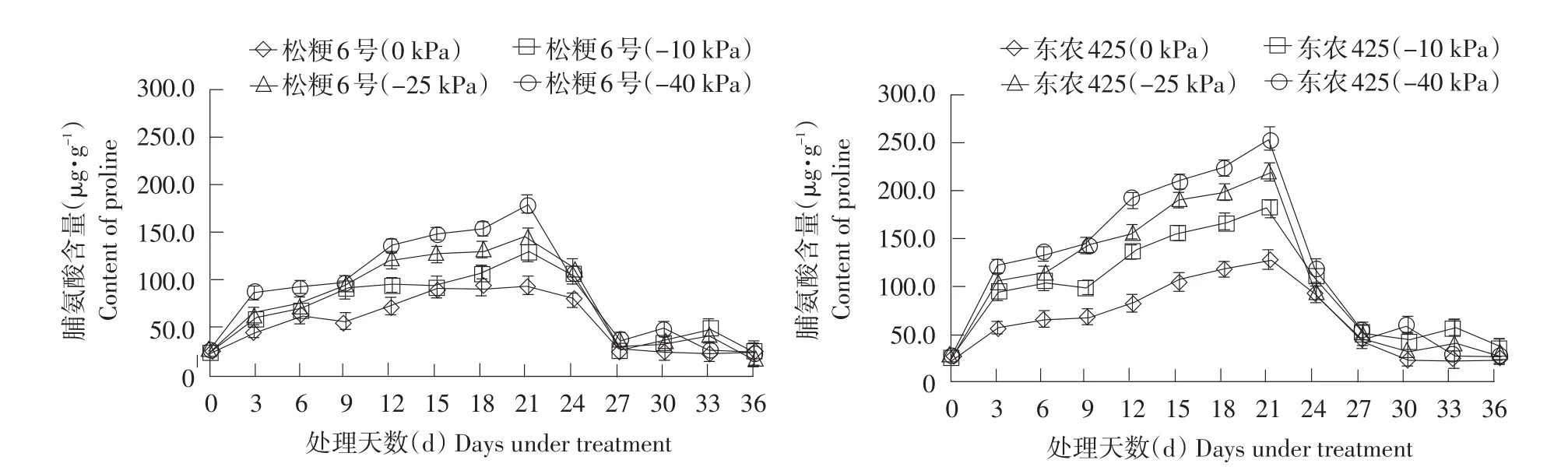
图3干旱胁迫下水稻脯氨酸含量变化Fig. 3 Change of rice Pro content under drought stress
3讨 论
作物抗旱性不仅与作物种类、品种基因型、形态性状及生理生化反应有关,受干旱程度发生时期、强度及持续时间影响,是植物水分与生理功能相互作用结果,也是植物与环境相互作用结果[18]。而分蘖期干旱胁迫阶段是水稻节水栽培技术实施主要时期。不同程度干旱胁迫对植物内部生理代谢产生影响,不同作物或品种适应干旱方式多样,具有不同的抗旱机制或多种抗旱机制共同发挥作用。SOD、POD和APX等酶类和物质是逆境条件下植物保持活性氧积累与清除系统平衡的重要物质[6]。王贺正等研究认为,水稻抗氧化酶活性干旱胁迫下显著提高[18-19]。本试验研究表明,干旱处理下,东农425的O2-积累显著低于松粳6号。SOD、POD和APX活性随干旱程度增强而提高,减轻活性氧对水稻伤害。抗旱品种具有较高抗氧化酶活性[7, 20]。干旱处理后东农425各处理SOD、POD活性较松粳6号均有小幅提升,APX活性则较松粳6号显著提升。复水后较长时间内两品种SOD酶活性均高于对照组,与陈小荣研究结果一致[9]。相比松粳6号,东农425 SOD酶在复水后活性下降较慢,使其拥有较长高酶活期,对O2-清除有重要作用。干旱胁迫后两品种APX酶活性均迅速提升,松粳6号与东农425分别于第9天及第12天达到峰值,当水稻H2O2积累时,APX迅速提高并分解H2O2,POD活性迅速提升取代APX功能,APX活性降低。东农425较松粳6号具有更高APX酶活性与更长的高酶活持续期,对清除H2O2,减缓O2-积累具有重要意义。
渗透调节是水稻适应水分胁迫的主要生理机制,通过渗透调节可使植物在干旱条件下保持一定膨压,维持细胞生长、气孔开放和光合作用等生理过程[21]。脯氨酸是植物在水分胁迫下最有效渗透调节物质[22],与植物抗旱性关系密切[23]。水稻游离脯氨酸积累不仅取决于水稻水分变化,同时与水稻存在的抗旱机制有关。随处理时间延长,水稻游离脯氨酸积累量增加[24-25]。蔡昆争等认为,干旱敏感型水稻干旱胁迫后脯氨酸含量上升幅度大于耐旱型[25],而杨建昌等研究则认为低土壤水分条件下耐旱型水稻脯氨酸累积较早、持续时间长[26]。但水分胁迫下脯氨酸累积和植物抗旱性关系以及是否可作为抗旱指标尚存争议[27-30]。本研究结果显示,两品种干旱胁迫下脯氨酸含量、积累量与干旱程度呈正相关。相对抗旱的东农425脯氨酸积累量及积累速率均显著高于敏感型品种松粳6号。复水后,东农425脯氨酸含量较松粳6号下降更迅速,说明东农425较松粳6号具有更快速的渗透条件能力,对于减轻膜脂损伤具有重要意义。
4结 论
本试验中,干旱胁迫时水稻快速提高APX酶活性,降低由于H2O2积累而导致的损伤,POD酶活性在APX酶达到峰值后迅速升高,取代APX酶成为清除H2O2的主要机制。东农425较松粳6号抗氧化酶活性具有更强的O2-、H2O2清除能力。复水后,东农425具有较长的高酶活持续期,对O2-清除具有重要意义。干旱处理后,东农425渗透调节物质脯氨酸含量显著高于同时期干旱敏感型品种松粳6号,且复水后恢复能力更强。
[参考文献]
[ 1 ]李树杏,郭慧,李敏,等.水稻对幼穗形成期水分胁迫后复水的响应[J].华北农学报, 2014(29): 206-212.
[ 2 ]信乃诠.农业水资源面临严重短缺的战略思考[J].灌溉排水学报, 1991(3): 23-26.
[ 3 ]姚林,郑华斌,刘建霞,等.分蘖期水分胁迫对不同栽培方式水稻生长发育及产量的影响[J].作物研究, 2014(5): 455-460.
[ 4 ]孙骏威,杨勇,蒋德安.水分亏缺下水稻的光化学和抗氧化应答[J].浙江大学学报:农业与生命科学版, 2004, 30(3): 278-284.
[ 5 ] Asada K. The water-water cycle in chloroplaste: Scavenging of active oxygens and dissipation of excess photons[J]. Annual Review of Plant Physiology & Plant Molecular Biology, 1999, 50 (4): 601-639.
[ 6 ] Mehdy M C. Active oxygen species in plant defense against pathogens[J]. Plant Physiol, 1994, 105: 467-472.
[ 7 ]李长明,蔡锡贵.水稻抗旱机理研究[J].西南大学学报:自然科学版, 1993(5): 409-413.
[ 8 ]蔡昆争,吴学祝,骆世明,等.不同生育期水分胁迫对水稻根系活力、叶片水势和保护酶活性的影响[J].华南农业大学学报, 2008, 29(2): 7-10.
[ 9 ]陈小荣,刘灵燕,严崇虎,等.抽穗期干旱复水对不同产量早稻品种结实及一些生理指标的影响[J].中国水稻科学, 2013(27): 77-83.
[10]王文斌,王晓怡,张明辉,等.大豆(Glycine max(L.)Merr.)叶片抗氧化酶对干旱及复水的响应机制[J].山西农业大学学报:自然科学版, 2012(32): 193-197.
[11]张皓政,赵宏伟,陈宾宾,等.不同灌溉方式下氮肥施用量对寒地粳稻抗氧化酶活性及产量的影响[J].灌溉排水学报, 2014(3):86-91.
[12] Wang H H, Huang J J, Bi Y R.Nitrate reductase-dependent nitric oxide production is involved in aluminum tolerance in red kidney bean roots[J]. Plant Science, 2010, 179(3): 281-288.
[13] Kraus T E, Fletcher R A. Paclobutrazol protects wheat seedlings from heat and paraquat injury. Is detoxification of active oxygen involved?[J]. Plant Cell Physiol, 1994, 35: 45-52.
[14]王爱国,罗广华,广州,等.植物的超氧物自由基与羟胺反应的定量关系[J].植物生理学报, 1990(6): 55-57.
[15]邹琦.植物生理学实验指导书[M].北京:中国农业出版社,2000.
[16] Solomon A, Beer S, Waisel Y, et al. Effects of NaCl on the carboxylating activity of rubisco from Tamarix jordanis in the presence and absence of proline-related compatible solutes[J]. Physiologia Plantarum, 1994, 90(1): 198-204.
[17]龚明.作物抗旱性鉴定方法与指标及其综合评价[J].云南农业大学学报, 1989, 4(1): 73-80.
[18]王贺正,马均,李旭毅,等.水分胁迫对水稻结实期活性氧产生和保护系统的影响[J].中国农业科学, 2007, 40(7): 1379-1387.
[19]朱维琴,吴良欢,陶勤南.干旱逆境对不同品种水稻生长、渗透调节物质含量及保护酶活性的影响[J].科技通报, 2006(22): 176-181.
[20]李德全,邹琦.抗旱性不同的冬小麦品种渗透调节能力的研究[J].山东农业大学学报:自然科学版, 1991(4): 377-383.
[21] Liang J, Zhang J, Wong M H.Stomatal conductance in relation to xylem sap ABA concentration in two tropical trees, Acacia confusa and Litsea glutinosa[J].Plant Cell Environ, 1996, 19: 93-100.
[22]苏梦云,范铭庆.渗透胁迫和钙处理对杉木幼苗膜脂过氧化及保持酶活性的影响[J].林业科学研究, 2000, 13(4): 391-396.
[23]梁芳.水分胁迫下杂交稻幼苗脯氨酸积累对膜脂过氧化的影响[J].矿业工程研究, 2001(23): 83-86.
[24]孙耀中,东方阳,郭学民,等.干旱胁迫下转甜菜碱醛脱氢酶基因水稻花后生理特性及产量构成[J].干旱地区农业研究, 2005(23): 108-113.
[25]蔡昆争,吴学祝,骆世明.不同生育期水分胁迫对水稻根叶渗透调节物质变化的影响[J].植物生态学报, 2008, 32(2): 491-500.
[26]杨建昌,王志琴.水稻在不同土壤水分状况下脯氨酸的累积与抗旱性的关系[J].中国水稻科学, 1995(9): 92-96.
[27]刘娥娥,宗会,郭振飞,等.干旱、盐和低温胁迫对水稻幼苗脯氨酸含量的影响[J].热带亚热带植物学报, 2000, 8(3): 235-238.
[28] Bhaskaran S , Smith R H, Newton R J. Physiological changes in cultured sorghum cells in response to induced water stress: I. Free proline[J]. Plant Physiology, 1985, 79(1): 266-269.
[29] Hanson A D, Nelsen C E, Everson E H. Evaluation of free proline accumulation as an index of drought resistance using two contrasting barley cultivars1[J]. Crop Sci, 1997, 17: 720-726.
[30]王洪预,李秋祝,赵宏伟,等.不同生育时期干旱处理对春玉米保护酶活性及产量的影响[J].东北农业大学学报, 2007, 38(1): 13-17.
Effect of drought stress and rewatering on antioxidant system and proline in rice during tillering stage
ZHAO Hongwei, WANG Xinpeng, YU Meifang, SHA Hanjing, JIA Yan, YU Tiancong, ZOU Detang(School of Agriculture, Northeast Agricultural University, Harbin 150030, China)
Abstract:In order to clarify the mechanism of drought tolerance in rice, Songjing6 (drought sensitive) and Dongnong425 (drought resistance) were used as experimental materials, drought stress treatment begun with tillering stage,ended with 21 d after treatment (soil water potential was 0,-10,-25 and -40 kPa, respectively). the effects of drought stress and rewatering treatment on antioxidant system in different drought-tolerant rice varieties during tillering stage were investigated. The results showed that superoxide anionand proline contents, superoxide dismutase (SOD), peroxidase (POD) and ascorbic acid peroxidase (APX) activities in two rice varieties increased with the days of drought stress during tillering stage, furthermore the more serious of drought stress (the lower water potential), the greater the rise of that. Moreover, the indicators except APX rapidly declined after rewatering treatment. The content ofwas lower in Dongnong425 (drought resistance) than that of Songjing6 under drought stress, the activities of SOD, POD, APX and the content of proline were higher in Dongnong425 than that of Songjing 6 under drought stress. The antioxidant enzyme activities and proline content could
干旱是造成农作物减产主要灾害。水稻是主要粮食作物,也是耗水量最大的灌溉作物[1]。水资源短缺现已成为制约水稻生产主要矛盾[2],水稻生产中节水栽培研究非常重要。水稻不同生育阶段水分胁迫后反应不同,不同生育期敏感性不同。反应敏感阶段水分胁迫造成光合作用减弱,生长发育受阻,严重影响产量[3]。特定时期(如分蘖盛期)排干水分成为水稻生产重要措施[4]。
水稻在水分短缺时会启动响应机制以减少缺水对生长发育的不利影响,其中抗氧化系统应激表现最明显。Asada认为,CO2同化作用限制先于电子传递反应钝化,因此在遭遇干旱时,植株发生产能过剩,光合电子链过度还原,产生活性氧类物质[5]。抗氧化系统中SOD主要清除活性氧,产物H2O2被POD、CAT继续分解为对植物体无害物质。但随干旱强度加强和胁迫时间延长,活性氧不断积累最终超过SOD分解速率,使膜脂过氧化,生成MDA,水稻植株受到伤害。Mehdy研究认为,干旱胁迫对植物体伤害源于细胞内自由基产生与清除不平衡[6]。李长明等研究发现,水分胁迫条件下,水稻抗旱性强弱与叶片SOD活性升高呈正相关,有助于缓解水分亏缺影响[7]。植物耐旱性体现在胁迫期间对逆境适应性和胁迫解除后植物恢复性生长。蔡昆争、陈小荣等研究认为,SOD酶活性在干旱复水后仍高于对照[8-9],而POD、APX酶活性最终恢复到与对照相近水平[8]。王文斌等研究认为,大豆干旱复水过程SOD、APX协同作用能有效清除活性氧避免膜伤害[10]。目前尚缺乏不同耐旱型水稻品种在干旱胁迫及复水后抗氧化系统变化对比研究。本试验研究分蘖期干旱胁迫及复水对两个不同耐旱型水稻品种抗氧化系统影响,以期明确干旱敏感型与耐旱型品种不同水分亏缺下抗氧化系统变化规律,揭示水稻耐旱生理机制。
1 材料与方法
1.1试验材料
本试验采用东农425(耐旱型)及松粳6号(干旱敏感型)两个水稻品种[11-12]。
1.2试验设计
试验于2014年在东北农业大学香坊实验基地防雨棚内进行(东经126.73°,北纬45.74°)。采用35 cm(内径)×35 cm(高)塑料盆钵,每盆装土10 kg,土壤有机质含量(22.34±0.12)g·kg-1,全氮量(1.24±0.06)g·kg-1,全磷量(0.41±0.04)g·kg-1,缓效钾(706.5±2.34)mg·kg-1,速效氮38.64 mg·kg-1,碱解氮(129.8±4.34)mg·kg-1,速效磷26.22 mg·kg-1,pH(6.75±0.04)。试验于4月18日播种,5月30日移栽,每盆4穴,每穴3株。施肥标准按基肥每盆1.67 g尿素(150 kg·hm-2)、1.11 g二胺(100 kg·hm-2)、0.83 g硫酸钾(75 kg·hm-2);蘖肥1.11 g尿素(100 kg·hm-2)。
试验采用完全随机试验设计,土壤干旱胁迫设3个梯度,土壤水势分别为-10、-25和-40 kPa,正常灌溉为对照,每个处理30盆,干旱胁迫处理21 d后复水,各处理代号见表1。干旱处理前7~10 d开始控水,逐渐降低土壤水势,临近处理期缺水时通过供水管补充少量水(每6 h观察1次,逐步减少每次补水量),将土壤水势控制在设计水势以上,处理前2~3 d停止补水。处理期间盆钵置于防雨棚内。用土壤张力计(中科院南京土壤研究所生产)检测土壤水势,每日6: 00、12: 00、18: 00分别记录土壤水势1次,以3次平均值代表当日盆钵土壤水势值。缺水时及时补水,并向无张力计盆钵加入等量蒸馏水。孕穗期复水后管理同大田生产。
1.3测定项目与方法
自处理开始后取样,每3 d取样一次,至复水后15 d结束,每处理随机选取有代表性3穴取样,取上部完全展开的三片叶。
SOD采用氮蓝四唑(NBT)光氧化还原法测定[13],POD采用愈创木酚法测定[13],APX采用290 nm比色法测定[14],超氧阴离子采用王爱国等方法测定[14],脯氨酸采用茚三酮显色、甲苯萃取分光光度法测定[15-16]。相对改变量计算公式如下:
相对改变量=每个指标峰值/每个指标初始值(干旱处理0 d数值)。maintaine at a high level for longer time in Dongnong425 after rewatering. The response time of APX was earlier than that of POD under drought stress.
Key words:rice; drought stress; antioxidant system; proline
作者简介:赵宏伟(1967-),女,教授,博士,博士生导师,研究方向为作物高产理论与栽培技术。E-mail: hongweizhao@163. com
基金项目:黑龙江省重大科技招标项目(GA14B102-02);“十二五”农村领域国家科技计划课题(2013BAD20B04)
收稿日期:2015-10-16
中图分类号:S511
文献标志码:A
文章编号:1005-9369(2016)02-0001-07
