猪苓和茯苓的研究现状及展望*
2015-12-24刘晶,王华,郭尚
刘 晶,王 华,郭 尚
(山西省农业科学院食用菌研究所,山西 太原 030031)
猪苓和茯苓的研究现状及展望*
刘 晶,王 华,郭 尚**
(山西省农业科学院食用菌研究所,山西 太原 030031)
猪苓和茯苓是两种著名的多孔菌科药用真菌。为进一步认识和利用这类宝贵资源,加快其基础研究及应用的步伐。从分类学地位、生物学特性、分布、分子生物学和人工培养等方面对猪苓和茯苓的研究现状进行总结,指出研究中存在的问题,并对研究前景进行了展望。
猪苓;茯苓;分类学;生物学特性;分布;分子生物学;人工培养
猪苓(Grifola umbellatus) 是著名的药用真菌。猪苓的子实体生于地下菌核上,质地柔软,味道鲜美,常被食用,即我们常说的“猪苓花”[1],而其地下菌核为药用部位。根据中医临床研究报道,猪苓的地下菌核对于急性胃炎、肝炎、肾炎,排尿问题、中暑腹泻、糖尿病及周身浮肿等疾病有很好的治疗效果[2]。
茯苓(Wolfiporia cocos)是中国常见的医药真菌之一,常在马尾松(Pinus massoniana)和赤松(Pinus densiflora)等松科植物根部寄生。其菌核生于地下,可以入药,在安神、利尿、镇定、降血糖等方面具有很高的医用价值[3-4]。茯苓多糖是茯苓的重要成分,与灵芝多糖的功效较为类似,尤其在抗肿瘤、抗诱变、增强免疫力以及促细胞分裂等生物活性方面有极高的应用前景[5]。
我国医疗事业正逐步发展,许多企业正在开发利用以猪苓和茯苓为原料的成品药物,其科学问题研究及人工栽培技术备受关注。本文对猪苓和茯苓的分类学地位、生物学特性、分布、生境、遗传多样性及人工栽培技术的研究现状进行总结,以期为猪苓和茯苓种质资源的保护、产品开发及利用提供理论依据。
1 分类学
猪苓 [Grifola umbellatus(Pers.ex Fr.)Pila],隶属于真菌门 (Eumycophyta) 担子菌纲 (Basidiomycetes)多孔菌目(Polyporales)多孔菌科(Polyporaceae)猪苓属(Grifola)[6-7]。1821年定名为Polyponus umbellatus(Pers.) Fr.,至今我国许多专业著作、科学杂志和药典仍沿用这一学名[8]。
猪苓的菌核常以3种形态呈现:(1)“猪屎苓”(P.umbellatusLBZ) 呈块状,长条形,体形大,分支少,表面光滑;(2)“铁蛋猪屎苓”(P. umbellatusLBT)常呈不规则球形;(3)“鸡屎苓”(P.umbellatusLBJ) 呈姜状,形像鸡爪,体形小,分支较多,表面有很深凹陷。已有分子生物学研究表明,这三种不同形态的猪苓可能属于同一物种[9],药效差异方面的内容尚未见报道。
茯苓 [Wolfiporia cocos(Schwein.)Ryvarden& Gilb.]是一种高等担子菌,隶属于多孔菌目(Polyporales)多孔菌科(Polyporaceae)茯苓属(Wolfiporia)[10-11]。茯苓的分类学地位被Ryvarden和Gilbertson(1984年)重新界定,新建茯苓属,茯苓被归于其中。此前,茯苓菌一直被看作低等真菌。这一命名和分类得到了广泛的认可并一直沿用到现在[12-15]。
2 生物学特性
猪苓和茯苓的生活史大致分为4个阶段:即菌丝体、菌核、子实体和担孢子。成熟的担孢子从菌管中弹射到外界,在适当的条件下萌生形成初生菌丝,有横膈膜,通过锁状联合形成双核菌丝。双核菌丝生长到一定阶段且受到外界刺激时,交叉、绕结,最终形成了菌核[16]。菌核可以将营养暂时贮存起来,当外部环境不适合时则发生休眠,在环境适宜时即可重新萌发形成初生菌丝[17]。
2.1菌核
猪苓的菌核是一种球形休眠体,由菌丝聚集而成,多年生,埋于地下,直径约3.0 cm~15.0 cm,其菌核的形状差异较大,以片形、块状和长条状居多。菌核表皮细胞趋于木质化,细胞壁加厚,表皮由几层排列紧密的“菌丝层”组成,其作用是维持体内的水分并阻止病虫害[18]。
猪苓菌核一般有3种,即表面呈黑色、白色和灰色,常被称为“黑苓”、“白苓”和“灰苓”。到了4月、5月,当地温达到10℃左右,菌丝即可萌发。当菌核表皮有菌丝突破,可看到有萌生出的绒毛状、洁白色的菌丝,随着菌丝的繁殖变化,形成的菌球如米粒般大小。表面有一层致密的菌丝排成白色的膜(对菌丝起保护作用),即“白苓”。到了6月、7月,当地温约18℃时,“白苓”快速生长。入秋后,随着地温的下降,菌核又渐慢生长,越冬后颜色变黄,即成为“灰苓”。经过夏、秋两季以后,“灰苓”变成黑褐色,又一个冬季过后,菌核几乎全部变成黑色,即成为“黑苓”。实际上,之所以产生3种菌核,是因为猪苓菌核生长年限的不同(第1年、第2年、第3年),才分别把它们称作“白苓”、“灰苓”和“黑苓”[19-20]。
经显微观察,发现猪苓菌核由表皮和髓质组成,且含有发生于菌丝中间或顶端的含晶细胞[21]。
茯苓的菌核即平常所讲的“茯苓”。大量菌丝相互集结而分化形成茯苓菌核,常在土壤中的松木或松根上发育,具特殊的味道。菌核形态各异,有椭圆形、球形等,常呈板状、不规则块状等。菌核表面较为粗糙,有瘤状褶皱,新鲜时为棕褐色,风干后为黑褐色,其表面为一层皮壳状的外皮,俗称“茯苓皮”[22]。
当菌丝体大量集结而占满了整个营养基质之后,茯苓菌丝才开始在适宜处发育,形成菌核。随着菌核的长大,茯苓皮被胀破,许多裂痕形成。新生菌丝和分泌的内含物相互弥合,导致“茯苓皮”的表面积不断增加,从而适应内部增长的需求。这种方式在生物界中是很特殊的。
因此,裂痕多又有乳状液汁溢出的茯苓菌核可作为茯苓生长旺盛的重要标记,可作为筛选优良菌种的重要根据[22]。
2.2子实体
每年的7月~9月是猪苓子实体生长成熟的时节。在夏秋季,当外界条件适宜时,菌核顶端即可从地下(接近地表)或刚出地表处生出子实体,形成多个分支的菌柄,每个分支都有一个菌盖在顶端。这种圆形菌盖直径约1 cm~4 cm,灰白色,有放射状、无环纹的淡黄色鳞片,摸起来像软毛,边缘较薄,向内翻卷[23]。猪苓子实体的菌丝细胞狭长,分支较多,直径在0.95 μm~4.55 μm,具有很薄的细胞壁[24]。
茯苓菌核发育到一定程度时向上膨大,部分菌核露出土面。在适宜条件下(温度约24℃~26℃,湿度约70%~85%),菌核会产生一层白色的“蜂巢状结构”,即茯苓的子实体。茯苓子实体的形成可以由菌丝体直接产生,不一定要经过菌核阶段。在实验室用斜面或平板培养茯苓菌种时,菌丝体密集的地方也可以形成子实体。
茯苓子实体幼时为白色,后逐渐变为淡黄白色,呈蜂巢状,无柄,厚约0.3 cm~2 cm。其孔管密集,长度和子实层几乎相等,直径约为0.5 mm~2 mm。在孔管内壁表面有子实层生长发育,子实层由担子组成[22]。
2.3担子和担孢子
猪苓的担子大小为(17.0~21.0) μm×(5.6~8.5)μm,短棒状,无色透明,顶端有四孢子。孢子大小为(7.0~10.0) μm×(3.0~4.2) μm,卵圆形,无色光滑。生殖菌丝双核,上有分生孢子梗产生,新生孢子具有较为稠密的细胞质。当孢子脱离分生孢子梗后,呈扁平脐部的椭圆形[25]。
茯苓的担孢子大小仅为(6.0~2.5)μm×(11.0~3.5)μm,在显微镜下才能看清其形态和构造,且不易察觉担孢子的颜色。近圆柱形或长椭圆形,有一肉眼很难觉察的歪尖。所以,茯苓的担孢子常被描述为透明无色。实际上,经观察研究,茯苓的担孢子应该呈灰白色[22]。
2.4菌丝体
在固体培养基上培养时,猪苓菌的菌丝呈絮状,白色,圆形菌落,生长较快,有发达的气生菌丝。猪苓菌丝分支较多,有狭长的细胞,直径约1 μm~5 μm。菌落表面有生长点形成,继而生长速度变快(尤其是菌落的体积和形态),且有一个小菌核(呈白色)会在菌落中央出现。平板上很快长满绒毛状的菌丝,当菌丝颜色加深,菌丝束开始分化。菌丝的老化会影响黑色素的出现、草酸钙结晶和无性孢子的生长。
茯苓的营养器官有2种结构:单核菌丝体和双核菌丝体。单核菌丝体又称为初级菌丝体,由担孢子萌发形成。每个细胞中仅含有一个细胞核是其主要特征,这在担孢子萌发的时候可以清楚地看到。2个不同性别的单核菌丝体细胞彼此连接沟通,形成一个双核细胞,这种双核细胞逐渐发育形成双核菌丝体,即次生菌丝体。茯苓菌丝体主要以双核菌丝体的形式存在,在子实体、担孢子、菌核和营养阶段的菌丝中,都能发现这种双核结构。
在光学显微镜下观察到茯苓的菌丝具有许多分支和横隔膜,把菌丝分成很多线形的细胞,其宽度为2 μm~5 μm。
3 分布和生境
在世界范围内,猪苓主要分布在亚洲、欧洲和北美洲等北纬地区;在国内,主要分布在陕西、山西、河南、河北、云南和湖北等13个省份[26]。其中,在山西有许多县市均有分布,如忻州(岢岚、五台、代县)、太原(阳曲)、临汾(吉县、古县)、吕梁(文水、交城、兴县、岚县、汾阳)、晋城(沁水)、长治(武乡、黎城)、晋中(灵石、左权、和顺、介休)、朔州(应县)、霍州、阳泉(盂县)等地均有分布[27]。陕西的猪苓药效最好,云南的产量最大[28]。
猪苓常生长于海拔1000 m~2 000 m的山地次生林中,尤其在二阳坡(半阴半阳)、湿度30%~50%、腐殖质丰富、疏松沙质土壤(中性或微酸性)的地方生长较好[26,29]。猪苓分布常常呈直线或斜线分布,很不规律[30],猪苓菌核埋于地下40 cm及以上,常会因雨水冲刷等自然因素而露出。猪苓菌丝在20℃~25℃条件下生长较好,温度过高容易褐变老化,过低则生长缓慢;在土壤pH5.0~6.7、相对含水量50%~60%时生长较好[31]。
茯苓是一种以腐生为主(兼营寄生)的真菌[32],其菌丝体可扭结形成休眠组织。茯苓子实体的产生需有适当的散射光刺激,并可产生成熟的担孢子。人工栽培的茯苓在偏酸性的土壤中生长较好,最适pH为3[33]。在自然界中,茯苓多生长于pH为5~6的偏酸性砂质土壤中[10]。
茯苓主要分布于温带及亚热带地区,在亚洲(中国、日本、韩国、印度等)、北美洲(美国)、大洋洲(新西兰)、非洲等地均有发现[34],一般生长于松属(Pinus)植物的根部,偶尔也生长于其他树种根部[35]。茯苓主要分布于西南、华南和华中等地区,在我国大约一半省份均有茯苓分布[36-37]。茯苓的自然分布则较少,经调查发现,仅云南省有少量野生茯苓分布[38]。人工种植的茯苓主要集中于两大产区:以湖北、安徽为中心的大别山产区和以湖南、贵州为中心的产区。
茯苓主要分布于海拔700 m~1000 m,在1000 m以上因气温太低,不适合茯苓生长;而在700 m以下的低山地区,则因森林资源被破坏以及人类的开垦等因素影响了其自然分布。茯苓的分布常受土壤制约,茯苓适宜在酸性、含沙量60%~70%和疏松通气的土壤中生长。当坡度呈10°~35°时,土壤的排水和通气的状况被显著改善,适宜茯苓生长;若坡度过大,则易造成干旱和水土流失,不适于猪苓的生长和发育[22]。
4 遗传多样性
遗传多样性指种内不同群体间或群体内不同个体间遗传变异的总和,是生物多样性的基础和重要组成部分,其大小直接关系着一个物种的生存能力和竞争能力[39],研究物种的遗传多样性能更好地了解物种起源和种质资源分布,对野生资源保护和可持续发展具有重要意义[40]。
有关猪苓和茯苓遗传多样性的研究较少,使用的分子标记主要涉及随机扩增多态性DNA(random amplified polymorphic DNA,RAPD)、简单序列重复扩增多态性(inter-simple sequence repeat,ISSR)、核糖体DNA内转录间隔区(nuclear ribosomal DNA internal transcribed spacer,nrDNA ITS)。相关序列扩增多态性(sequence related amplified polymorphisms,SRAP)的序列见表1。虽然这些研究使用的样品数和取样范围比较有限,但这些研究大都表明猪苓菌种群存在明显的遗传分化,而茯苓菌种群存在较小差异,分化程度较低[41]。
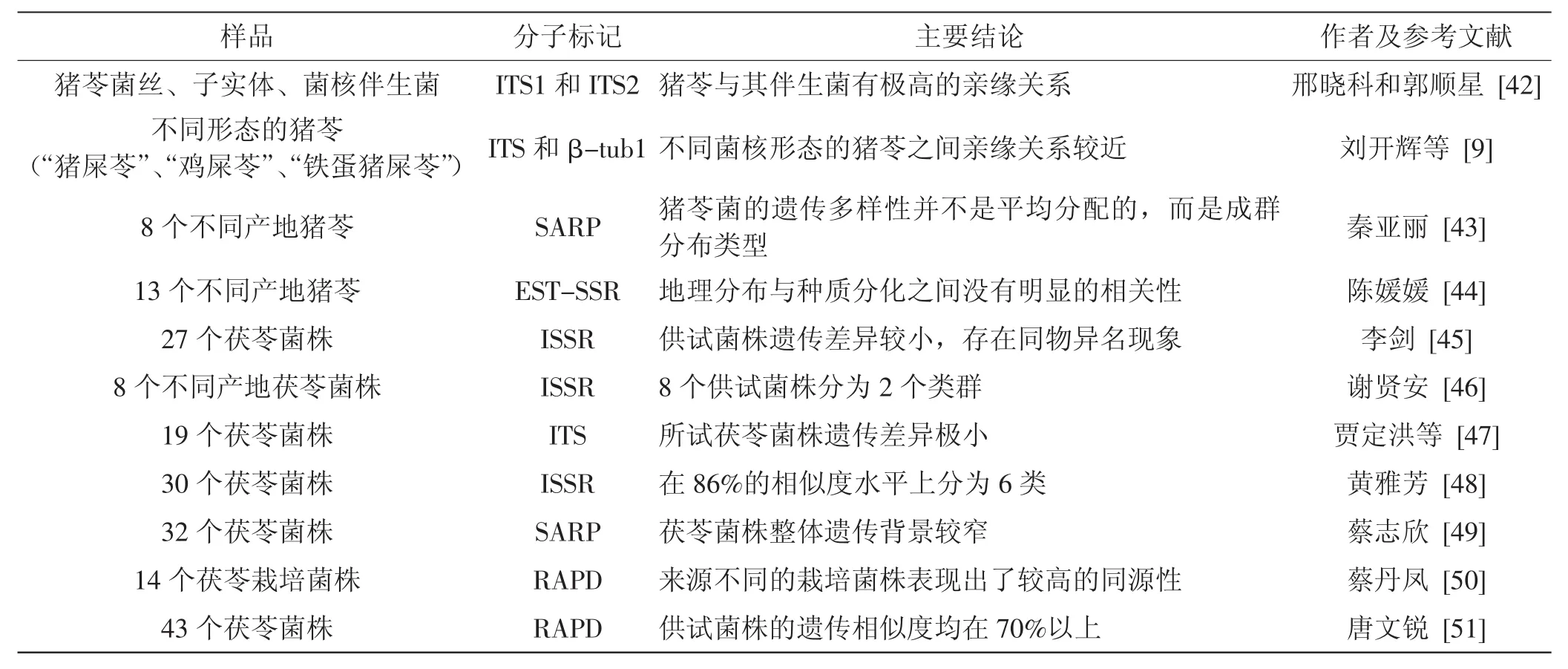
表1 猪苓和茯苓遗传多样性研究的相关文献Tab.1 Literature summary on genetic diversity studies of G.umbellatus and W.cocos
5 人工栽培
随着野生环境被不断破坏,再加上人们肆意地采挖猪苓,致使野生猪苓不断减少。我国早从20世纪70年代就开始研究了猪苓的人工栽培技术,大致可以分为5个阶段,见表2。

表2 猪苓人工栽培现状和存在问题Tab.2 Artificial cultivation advances and issues of G.umbellatus
然而,虽然一些基本的栽培技术已经被掌握,但至今离产业化的实际要求尚有以下4个方面的关键问题(表2),有待突破解决。
目前,实现茯苓人工栽培的国家只有中国,其栽培历史主要经历了4个阶段:据南北朝时期的《本草经集注》记载,我国早在1500年前就采用砍伐松树的方法人工栽培茯苓;原始的栽培方法经过传承和演变,逐步形成了以松树段为原料,以新鲜茯苓菌核作为菌种的栽培阶段;20世纪70年代以后,茯苓纯种菌丝被成功分离,之前的“新鲜菌核”逐步被“茯苓纯菌种”取代;21世纪以来,茯苓的“诱引栽培法”得到推广和应用,使茯苓的单产和总产量得到了大幅提升。
在栽培类型上,茯苓主要有树桩原地栽培、段木栽培、活树栽培、树兜栽培、松木屑生料栽培、松枝松叶栽培等人工栽培方法[59]。最常用的是短段木栽培法,但这种方法消耗大量的木材,会对产区的生态环境造成影响。茯苓产业的发展导致产区松木减少,从而又制约了产业自身的发展。但目前的研究说明代料栽培可以替代松木栽培,具有很大的推广前景[45,60-61]。
尽管我国茯苓的人工栽培技术已经较为成熟,研究主要集中在栽培基质和育种方面[45,60-61],但茯苓的育种工作进展相对缓慢。目前主要采用组织分离的传统方法来筛选优良菌株。研究发现,通过紫外诱变、原生质体融合、菌丝尖端转接、单孢配对[62-63]等方法均可达到筛选优良菌株的目的,但采用原生质体融合和紫外诱变的方法所育成的菌株尚未大量用于生产。研究还发现,采用液体发酵时,其菌丝产量显著高于原始菌株,且胞外多糖和菌丝内的多糖含量远高于原始菌株[64-65]。
6 存在问题及展望
近年来,猪苓和茯苓在很多方面都取得了重要的研究进展。然而,研究中也存在一定问题。例如,目前有关猪苓菌系统发育学的研究较少,而使用多个核基因序列进行这方面研究的报道更是少之又少。研究所取的样品范围也极其有限。猪苓有着丰富的遗传多样性[66-67],Xing XK et al[68]使用nrDNA ITS和LSU(large subunit) 序列讨论了猪苓菌的遗传多样性,提出陕西、河南和甘肃的猪苓产地应该重点保护。为了更系统地研究猪苓的种质关系、确保药效、合理利用和保护现存的有限种质资源,对其遗传多样性的研究已迫在眉睫。猪苓、蜜环菌(Armillaria mellea)和半生菌是否具有协同进化的关系,猪苓与蜜环菌共生的信号机制等,也是很值得研究的课题。
不少研究已经表明茯苓菌的遗传分化程度较低(表1)。熊斌[65]通过栽培试验发现某些并不适合用于栽培生产的菌株,也具有某种突出的优良性状,但是随意引种可能带来减产甚至绝收的风险,在茯苓的栽培生产过程中应加强引种的规范性。由于对茯苓菌的交配型还不是很清楚,无法进行杂交育种。因此,利用遗传育种学和分子生物学研究茯苓菌的交配型,对茯苓育种和实现大规模人工栽培具有重要意义。
研究猪苓和茯苓的最终目标就是要开发出物美价廉的药物,但实现两种菌的规模化人工栽培还有很长的路要走。随着不同领域科学家的联合攻关及社会各界的重视,这些重要的药用真菌一定能够早日为人类造福。
[1]张长青,张松柏,孙正祥.猪苓的称谓及学名更替[J].现代农业科技,2009,22(1):211-212.
[2]罗英,李梁.猪苓生长的土壤条件研究[J].核农学报,2002,16(2):115-118.
[3]杨天星,张学莉.茯苓沿革及现代化发展[J].中国民族民间医药杂志,2004,70(1):289-291.
[4]刘忠义,曾虹燕.茯苓液体培养研究初探[J].湘潭大学自然科学学报,2002,24(4):49-51.
[5]丁琼,张俐娜,张志强.茯苓菌丝体多糖的分离及结构分析[J].高分子学报,2000,2(1):224-227.
[6]黄年来.中国食用菌百科[M].北京:中国农业出版社,1993:158-181.
[7]牛西午.北方食用菌栽培[M].北京:中国科学技术出版社,1994:274-276.
[8]徐锦堂.中国药用真菌学[M].北京:中国医科大学·中国协和医科大学联合出版社,1997.
[9]刘开辉,邓百万,陈文强,等.比较DNA序列分析不同猪苓菌种的亲缘关系[J].食用菌学报,2009,16(3):11-13.
[10]吕作舟.食用菌栽培学[M].北京:高等教育出版社,2006:311-342.
[11]赵继鼎等.中国真菌志(第三卷):多孔菌科[M].北京:科学出版社,1998:411-414.
[12]熊杰,林芳灿,王克勤,等.茯苓基本生物学特性研究[J].菌物学报,2006a,25(3):446-453.
[13]熊杰.茯苓性模式的研究[D].武汉:华中农业大学,2006b.
[14]孟虎.茯苓性不亲和模式的初步研究[D].武汉:华中农业大学,2012.
[15]王克勤,傅杰.药用茯苓名实考[J].中药材,2002,25(7):508-509.
[16]邓百万,陈文强.猪苓菌丝液体培养及富锌研究[J].中国食用菌,2003,22(1):33-34.
[17]周微微.猪苓菌核及发酵菌丝体化学成分研究及质量分析[D].北京:中国协和医科大学,2008.
[18]郭顺星,徐锦堂.猪苓菌核结构性质的研究[J].真菌学报,1991,10(4):312-317.
[19]徐锦堂,郭顺星,李灵玉,等.猪苓菌核生长发育规律观察[J].中国药学杂志,1991,26(12):714-716.
[20]许广波,傅伟杰,曹丽,等.猪苓菌丝体液体培养基配方的筛选研究初报[J].吉林农业大学学报,2002,24(2):64-65.
[21]邢晓科,郭顺星.猪苓与其伴生菌在几种不同培养基上的生长特性[J].中国药学杂志,2005,40(6):417-420.
[22]李益健.茯苓生物学特性和特征的研究[J].武汉大学学报:自然科学版,1979,3(1):107-115.
[23]戚淑威,赵琪,程远辉,等.猪苓的研究进展[J].云南农业科技,2011,5(1):7-9.
[24]周元,梁宗锁,段琦梅.不同来源的猪苓菌株菌丝生物学特性比较[J].微生物学杂志,2008,28(6):14-18.
[25]Xing XK,Guo SX.Electron microscopic study of conidia produced by the mycelium of Polyporus umbellatus[J].Myccosystema,2008,27(4):554-558.
[26]徐锦堂.中国药用真菌学[M].北京:中国医科大学·中国协和医科大学联合出版社,1997.
[27]李雯瑞,梁宗锁,陈德育.猪苓生物学特性的研究进展[J].西北林学院学报,2012,27(6):60-65.
[28]金若忠,范俊岗,栾庆书,等.猪苓生物学特性及菌种培养[J].辽宁林业科技,2010,3(1):35-37.
[29]田飞,曹晓燕,王喆之,等.影响猪苓产量因素的调查研究[J].陕西农业科学,2010,2(1):106-109.
[30]徐科焕,姚军强,赵万平,等.太白山野生猪苓化学成分、药用价值与生态分布研究初报[J].中国野生植物资源,2008,27(3):25-27.
[31]陈文强,邓百万,刘开辉,等.中低海拔地区猪苓人工栽培技术[J].江苏农业科学,2007(4):167-169.
[32]赵继鼎,张小青.中国多孔菌类群真菌生态、分布与资源[J].生态学报,1994,14(4):437-443.
[33]蔡丹凤,陈美元,郭仲杰,等.茯苓菌株生物学特性的研究[J].中国食用菌,2009,28(1):23-26.
[34]Wang YZ,Zhang J,Zhao YL,et al.Mycology,cultivatbn, traditional uses,phytochemistry and pharmacology of Wolfiporia cocos(Schwein.)Ryvarden et Gilb.:A review[J]. Journal of Ethnopharmacology,2013,147(2):265-276.
[35]Li GY,Huang LH,Qin TF,et al.Chemical compositions,infrared spectroscopy and X-ray diffractometry study on brown-rotted woods[J].Carbohydrate Polymers,2011,85(3): 560-564.
[36]喻宗源.茯苓研究进展[J].湖北化工,1991,1(1):10-16.
[37]王克勤,尹旭仁,黄鹤,等.湖北茯苓生产现状及产业化发展对策[J].中国现代中药,2013,14(12):24-27.
[38]於小波,昝俊峰,王金波,等.我国茯苓药材主要产区资源调查[J].时珍国医国药,2011,22(3):714-716.
[39]彭飒.红花种质资源的SRAP分子鉴定及其品质相关的分子标记研究[D].新疆:新疆农业大学,2006.
[40]张建博,桂明英,刘蓓,等.分子生物学在大型真菌遗传多样性研究中的应用[J].中国食用菌,2008,27(6):3-7.
[41]曲直,陶刚,朱国胜,等.茯苓种质资源的RAPD分析[J].菌物研究,2008,6(3):170-174.
[42]刑晓科,郭顺星.从ITS序列探讨猪苓与其伴生菌的亲缘关系[J].微生物学通报,2004,31(2):34-36.
[43]秦亚丽.猪苓遗传多样性及多糖合成酶UGPase基因的克隆[D].杨林:西北农林科技大学,2012.
[44]陈媛媛.猪苓EST-SSR标记的开发及遗传多样性研究[D].杨林:西北农林科技大学,2014.
[45]李剑.茯苓种质资源多样性与代料栽培技术初步研究[M].武汉:华中农业大学,2007.
[46]谢贤安,汪思迪,曾晓丽,等.茯苓菌属遗传多样性的ISSR分析[J].湖北农业科学,2008,4(10):1111-1113.
[47]贾定洪,王波,彭卫红,等.19个药用茯苓菌株的ITS序列分析[J].中国食用菌,2011,30(1):42-44.
[48]黄雅芳.茯苓菌种质量标准和种质资源研究[M].武汉:湖北中医药大学,2012.
[49]蔡志欣,蔡丹凤,陈美元,等.32个茯苓菌株的SRAP分析[J].食药用菌,2013,21(2):96-98.
[50]蔡丹凤.茯苓松蔸栽培优良菌株的筛选研究[J].中国食用菌,2013,32(1):14-16.
[51]唐文锐.“菌核诱引”对茯苓新菌核产量和品质影响的初步研究[D].武汉:华中农业大学,2013.
[52]李雯瑞.猪苓优良菌株筛选与菌核生长发育过程的研究[D].杨林:西北农林科技大学,2013.
[53]郭顺星,徐锦堂,肖培根.猪苓生物学特性的研究进展[J].中国药学杂志,1996,21(9):515.
[54]郭顺星,王秋颖,张集慧,等.猪苓菌丝形成菌核栽培方法的研究[J].中国药学杂志,2001,36(10):658-660.
[55]陈文强,邓百万,张娟.猪苓液体培养基筛选研究[J].氨基酸和生物资源,2002,24(3):23-24.
[56]兰进,徐锦堂,贺秀霞.药用真菌栽培实用技术[M].北京:中国农业出版社,2001:210-220.
[57]林树钱.中国药用菌生产与产品开发[M].北京:中国农业出版社,2000:318-330.
[58]刘瑰琦,郑焕春,于长有,等.野外栽培猪苓的实用技术[J].特种经济动植物,2001,4(6):28.
[59]马巾媛.茯苓不同栽培条件下的生物学特性及质量评价[D].昆明:云南中医学院,2013.
[60]刘文志.代料栽培与传统栽培茯苓的质量比较及其复方的减肥降脂功能研究[D].武汉:湖北中医药大学,2012.
[61]杨祺.茯苓的代料栽培研究[D].武汉:湖北中医药大学,2013.
[62]熊欢.原生质体技术在茯苓菌种复壮、育种和生活史研究中的应用[D].武汉:华中农业大学,2009.
[63]诸发会.茯苓高产优质菌株的复壮技术研究[D].贵州:贵州大学,2008.
[64]李羿,万德光.茯苓紫外线诱变育种[J].药物生物技术,2008,15(1):44-47.
[65]熊斌.茯苓种质资源评价及遗传多样性研究[D].武汉:华中农业大学,2014.
[66]Dai YC.Polypore diversity in China with an annotated checklist of Chinese polypores[J].Mycoscience,2012,53(1):49-80.
[67]Zhang Y,Kang Y,Qin Y,et al.Genetic diversity of endangered Polyporus umbellatus from China assessed using a sequence-related amplified polymorphism technique[J].Genetic and Molecular Research,2012,11(4):4121-4129.
[68]Xing XK,Ma XT,Miranda MH,et al.Genetic diversity and evolution of Chinese traditional medicinal fungus Polyporus umbellatus(Polyporales,Basidiomycota)[J].Plo One,2012, 8(3):15-18.
Research Advances,Issues and Perspectives on Polyporus umbellatus and Wolfiporia cocos
LIU Jing,WANG Hua,GUO Shang
(College of Life Sciences,Shanxi University,Taiyuan 030006,China)
Polyporus umbellatus and Wolfiporia cocos are two of the best-known medicinal fungi.In the paper,the recent research progresses of the fungi on their taxonomy,biological characters,distribution,molecular biology and artificial cultivation were reviewed.Meanwhile the major issues remaining in the current research for these two fungal organisms were also discussed,and the approaches for future studies were put forward.
Polyporus umbellatus;Wolfiporia cocos;taxonomy;biological character;distribution;molecular biology;artificial cultivation
S646.9
A
1003-8310(2015)04-0007-07
10.13629/j.cnki.53-1054.2015.04.002
山西省煤基重点科技攻关项目(FT2014-03)。
刘晶(1985-),女,本科,助理研究员,主要从事食用菌的栽培技术和推广方面研究。E-mail:liyuan_198711@sina.com
**通信作者:郭尚(1968-),男,博士,副研究员,主要从事食用菌的栽培技术和推广方面研究。E-mail:gs0351@sohu.com
2015-05-20
