北虫草培养基添加量对酱油制曲及酶活性的影响
2015-07-13张杰张映赵博崔成斌张国财王滨松
张杰,张映,赵博 ,崔成斌,张国财*,王滨松
(1.东北林业大学a.生命科学学院;b.林学院,黑龙江 哈尔滨150040;2.黑龙江大学化学化工与材料学院,黑龙江 哈尔滨150080)
酱油作为中国传统发酵食品调料之一,以其丰富的营养成分和滑腻的口感而深受人们喜爱,是人们餐桌上不可缺少的调味食品[1]。伴随着人们对食品营养的高质量要求,单调、传统的发酵酱油已经不能满足人们的需求,开发具有保健功效的营养型酱油具有很大的市场前景[2]。酱油制曲的实质是通过培养曲霉菌得到曲菌所分泌的蛋白酶和糖化酶。在酱醅发酵中,蛋白酶分解蛋白质产生的各类氨基酸是酱油的主要呈味物质。糖化酶分解淀粉类物质产生葡萄糖。葡萄糖和氨基酸在较高温度下发生美拉德反应,生成褐色物质,使酱油呈红棕色,可见,酶在制曲过程中起到了很重要的作用[3]。北虫草培养基中不仅含有发酵基质,还含有北虫草中所特有的虫草素、虫草酸、虫草多糖等保健物质[4]。笔者以米曲霉沪酿3.042 单菌种[5]进行酱油制曲,研究北虫草培养基添加量对酱油制曲及酶活性的影响,旨在为规模化生产虫草酱油提供参考。
1 材料与方法
1.1 菌种与原料
试验菌种米曲霉沪酿3.042 来自于山东省淄博市源康源生物科技有限公司。制曲原料豆粕和麦麸均为市售。
1.2 主要试剂
主要试剂有福林试剂(Folin)、二硝基水杨酸(DNS)试剂及缓冲液。
1.3 方法
1.3.1 种曲制备
将豆粕40g 和麸皮60g 混合均匀(豆粕和麸皮的质量比为2∶3),润以100%的水,筛去粗粒,将混合好的料装入1 000mL 三角瓶(装入厚度1 cm 左右),再于121℃、0.1 MPa 条件下维持30min 进行高压灭菌。冷却后分别添加5、10、15、20g 北虫草培养基(分别称其为处理1、处理2、处理3、处理4),以不添加北虫草培养基的处理为对照,再分别以0.3%接种量加入沪酿3.042,30℃水浴恒温加热,转入摇瓶培养以防止结块,待瓶中充满孢子即可使用,或置冰箱备用。
1.3.2 种曲质量的测定
1) 孢子数的测定。 以每1g 曲料中孢子的含量为计量单位,采用血球计数板法检测种曲孢子数。一般要求每1g 干基的孢子数在60亿个以上。
2) 感官品质的观察。观察制曲24~60 h 成曲的感官品质。
1.3.3 酶活性的测定
蛋白酶活性采用Folin–酚法[6–8]进行测定。将40℃下每1min 水解酪蛋白产生1μg酪氨酸所需的酶量定义为1个蛋白酶活力单位(U)。
糖化酶活性参照文献[2,6]进行测定。将温度40℃、pH 5.0 条件下每1min 催化生成1mg 葡萄糖所需的酶量定义为一个酶活力单位(U)。
亮氨酸氨肽酶活性参考文献[9]的方法进行测定。将试验条件下每1min 水解L–亮氨酸–对硝基苯胺产生 1μg 对硝基苯胺所需的酶量定义为1个酶活力单位(U)。
纤维素酶活性参考文献[2]进行测定。将温度50℃、pH 4.8 条件下每1min 催化生成1μg 葡萄糖所需的酶量定义为一个酶活力单位(U)。
1.4 统计分析
用SPSS 13.0对数据进行分析,用Excel 2007辅助作图。利用SPSS One–Way ANOVA方差分析软件进行显著性分析。
2 结果与分析
2.1 各处理种曲在制曲过程中的感官变化
通过对种曲孢子数进行测定,测得处理1、处理2、处理3、处理4 每1g 发酵液中的孢子数分别为334亿、355亿、343亿、336亿个,处理2 中每1g 发酵液中的孢子数比对照高30亿个。由表1 可见,在制曲过程中,不同虫草培养基添加量对种曲感官产生的影响不同,与对照组相比,处理组米曲霉菌丝体生长厚实,处理2 米曲霉菌丝体在培养制曲48 h 曲料蓬松,菌丝丰满,米曲霉孢子呈淡黄色,接近成熟,孢子生长旺盛,在54 h 后蓬松的曲料中出现黄绿色丰满的成熟米曲霉孢子。综合孢子数分析和感官观察,认为处理2 发酵基料中的种曲质量最优。
2.2 各处理种曲在制曲过程中酶活性的变化
2.2.1 蛋白酶酶活性的变化
沪酿3.042 制曲能够产生3 种蛋白酶,且碱性蛋白酶、中性蛋白酶、酸性蛋白酶的酶活性依次减小,其原因是在单菌种米曲霉制曲过程中,初期制曲的pH 值较高,呈现出碱性,随着蛋白的分解,pH 值逐渐降低。
中性蛋白酶活性的变化:由图1 可知,对照和各处理在制曲36~48 h 均呈上升趋势,在制曲48~60 h均呈下降趋势;对照和各处理均在制曲48 h 出现最大值,且以处理2 的酶活性最大(每1g 干基的酶活性达1 801.21 U,比对照高9.01%);与对照相比较,在整个制曲过程中,各处理的中性蛋白酶活性均显著高于对照;各处理之间相比较,在制曲过程中,处理2 的中性蛋白酶活性均显著高于其他处理。

图1 制曲过程中各处理中性蛋白酶的活性 Fig.1 Neutral proteinase activity in the propagation process
碱性蛋白酶活性的变化:由图2 可见,对照和各处理在制曲36~48 h 均呈较快的上升趋势,在制曲48~60 h 呈缓慢下降趋势;对照和各处理均在制曲48 h 出现最大值,每1g 干基的酶活为2 078.05~2 253.96 U,其中处理2 的酶活性最大(每1g 干基的酶活性为2 253.96 U,比对照高8.46%);与对照相比,整个制曲过程中各处理的碱性蛋白酶活性均显著高于对照;各处理之间相比较,整个制曲过程中处理2 的碱性蛋白酶活性均显著高于其他处理。

图2 制曲过程中各处理碱性蛋白酶的活性 Fig.2 Alkaline protease activity in the propagation process
酸性蛋白酶活性的变化:由图3 可知,酸性蛋白酶的活性远远小于中性蛋白酶和碱性蛋白酶的活性;对照和各处理在制曲36 ~42 h 呈缓慢上升趋势,酶活性整体相差不大,且在制曲48~60 h 呈下降趋势;对照和各处理酶活性均在制曲42 h 出现最大值,其中处理2 的酶活性最大(每1g 干基的酶活为259.66 U,比对照高6.02%);与对照相比,除60 h 外,整个制曲过程中各处理的酶活性均显著高于对照;各处理之间相比较,整个制曲过程中处理2的酸性蛋白酶活性均高于其他处理(在制曲36、60 h,处理2 均显著高于其他处理,在制曲42 h,处理2、处理3 显著高于处理1 和处理4,在制曲48 h,处理2 显著高于处理1 和处理4)。
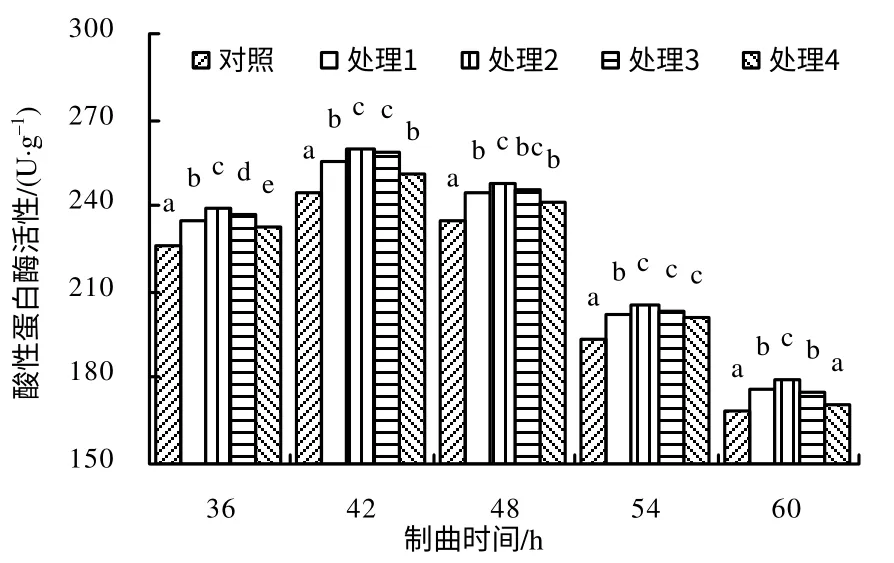
图3 制曲过程中各处理酸性蛋白酶的活性 Fig.3 Acid proteinase activity in the propagation process
2.2.2 α–淀粉酶活性的变化
由图4 可知,在制曲36~48 h,对照和各处理α–淀粉酶的活性呈上升趋势,在制曲48~60 h 呈下降趋势;在制曲48 h,对照和各处理均出现酶活性最大值,且以处理2 的酶活性最大(每1g 干基的酶活性为50.37 U,比对照高38.27%);与对照相比,在制曲36 h 和42 h,各处理组与对照组酶活性的差异无统计学意义,在制曲48 h,处理1 和处理2 的酶活性均显著高于对照;在制曲54 h 和60 h,处理1、处理2 和处理3 的酶活性均显著高于对照,处理4 与对照酶活性的差异无统计学意义;各处理之间相比较,在制曲36 h,各处理间酶活性的差异无统计学意义,在制曲42~60 h,处理1、处理2 与处理3 酶活性的差异均达显著水平,与处理4 的差异也均达显著水平。
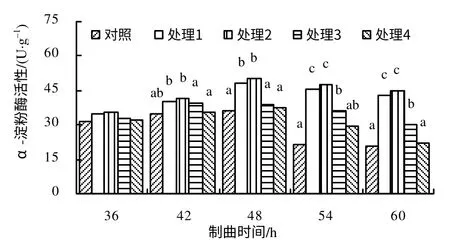
图4 制曲过程中各处理α–淀粉酶的活性Fig.4 α–amylase activity in the propagation process
2.2.3 亮氨酸氨肽酶活性的变化
由图5 可以看出,在制曲36~42 h,对照和各处理亮氨酸氨肽酶的活性呈缓慢上升趋势,42 h 后迅速增加,在48 h 出现酶活性峰值,之后迅速下降,其中处理2 的酶活性最高(每1g 干基的酶活为842.33 U,比对照高1.10%);整个制曲过程中,各处理的亮氨酸氨肽酶酶活性与对照间的差异均无统计学意义。
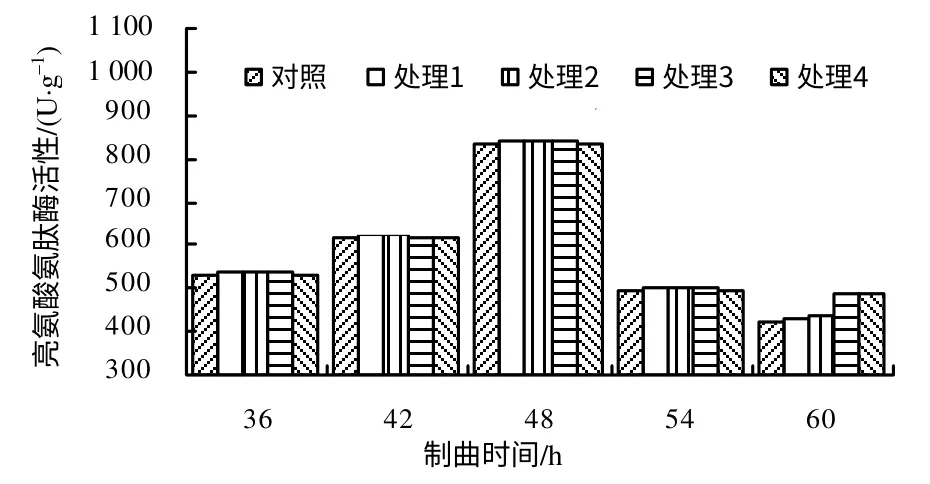
图5 制曲过程中各处理亮氨酸氨肽酶的活性 Fig. 5 Leucine aminopeptidase activity in the propagation process
2.2.4 纤维素酶活性的变化
由图6 可见,在制曲36~48 h,对照和各处理纤维素酶的活性缓慢上升,48~60 h 缓慢下降;对照和各处理均在制曲48 h 出现酶活性峰值,其中以处理2 的酶活性最高(每1g 干基的酶活为1 801.53 U,比对照高18.56%);与对照相比,整个制曲过程中,在制曲48、54、60 h,各处理的酶活性均显著高于对照;各处理之间相比较,整个制曲过程中处理2 的酶活性均高于其他处理(在制曲36、54 h,处理2 均显著高于其他处理组,在制曲48 h,处理2、处理3 均显著高于处理1 和处理4)。

图6 制曲过程中各处理纤维素酶的活性 Fig.6 Cellulase activity in the propagation process
3 结论与讨论
制曲是酱油发酵的关键。充分利用原料,改善酱油风味,提高制曲工艺水平,从而提升酱油品质,丰富酱油品种是酱油产业发展的必然趋势[10–11]。本研究结果表明:
在酱油发酵基料中分别添加5、10、15、20g北虫草培养基,以添加10g 北虫草培养基处理中每1g 发酵液中的孢子数最多,为355亿个,比对照高30亿个。该处理种曲的孢子数最多,孢子生长旺盛,制曲48 h 时曲料蓬松,菌丝丰满,米曲霉孢子为淡黄色;制曲54 h 时,蓬松的曲料中长出黄绿色丰满的成熟米曲霉孢子,由孢子数分析结果和感官观察结果可知,该处理发酵基料中的种曲质量最优。这与向培养基中添加丟糟和黄水提高制曲质量的研究结果[12–13]相似,且产生的孢子数有所增加。
酱油制曲过程中米曲霉产酶活性的高低将影响原料的利用率及产品的品质,因此,提高成曲酶活性是提高酱油品质的重要途径[14]。本研究结果表明,所测各酶的活性数值均呈先上升、后下降的趋势。除酸性蛋白酶在制曲42 h 出现最大值外,其余各 酶的活性均在制曲48 h 出现峰值。在整个制曲过程中,除亮氨酸氨肽酶酶活性各处理间与对照间的差异无统计学意义外,各处理中性蛋白酶、碱性蛋白酶、酸性蛋白酶、α–淀粉酶及纤维素酶的酶活性均高于对照,且在酶活高峰时,处理2 酶活性均显著高于对照组(P<0.05),表明添加北虫草培养基可以提高制曲过程中酶的活性,且以添加10g 北虫草培养基处理的酶活性较高。
将北虫草培养基作为原料加入到酱油发酵基料中,不仅能实现废物利用,解决酱油发酵的原料问题,而且能制造出具有虫草风味的功能型酱油。
[1] Wei C L,Chao S H,Tsai W B,et al.Analysis of bacterial diversity during the fermentation of inyu,a high- temperature fermented soy sauce,using nested PCR– denaturinggradientgel electrophoresis and the plate count method[J].Food Microbiology,2013,33(2):252–261.
[2] 赵强强.酱油制曲及螺旋藻酱油发酵工艺的研究[D].青岛:中国海洋大学,2012.
[3] 张艳芳.多菌株制曲促进酶系优化与提高酱油品质的研究[D].无锡:江南大学,2009.
[4] 林群英,宋斌,李泰辉.北虫草研究进展[J].微生物学通报,2006,4(3):154–157.
[5] 林祖申.米曲霉制曲过程中酶活性变化及其工艺优化[J].中国酿造,2007(5):56–59.
[6] 周探春,刘焱,邓放明.斑点叉尾鮰下脚料蛋白酶水解工艺优化[J].湖南农业大学学报:自然科学版,2013,39(1):95–98.
[7] 刘晓蓉,谭才邓,陈小冰,等.米曲霉1228 制曲条件的优化及酱油酿造的研究[J].现代食品科技,2013,29(2):291–293,348.
[8] GB/T23527—2009 蛋白酶制剂蛋白酶活力的测定 福林法[S].
[9] 潘进权.毛霉亮氨酸氨肽酶的纯化及性质研究[J].食品科学,2012,33(7):163–167.
[10] 李秀婷,赵进,鲁绯,等.米曲霉固态发酵产酶条件及酶活力研究[J].中国酿造,2009,203(2):26–28.
[11] 王欣宏.螺旋藻酱油发酵工艺研究以及特征性成分检测[D].青岛:中国海洋大学,2013.
[12] 边名鸿,宗绪岩,刘绪,等.丢糟在制曲生产中的添加量与大曲质量关系的初步研究[J].食品与发酵科技,2012(4):82–84,99.
[13] 石磊.制曲过程中适量添加黄水来提高大曲质量的研究[J].山东食品发酵,2004(1):25–26.
[14] 赵文婷,王颖,邱璠,等.加工工艺及添加面粉发酵对豆酱抗氧化能力和抗氧化成分的影响[J].中国酿造,2011(6):23–26.
