PRRSV新亚型毒株JL580的基因组变异特征研究
2015-04-27常晓博赵款叶超姜成刚王淑杰蔡雪辉张辉安同庆
常晓博,赵款,叶超,姜成刚,王淑杰,蔡雪辉,张辉,安同庆∗
(1.吉林农业大学动物科学技术学院,长春130118;2.中国农业科学院哈尔滨兽医研究所兽医生物技术国家重点实验室,哈尔滨150001)
PRRSV新亚型毒株JL580的基因组变异特征研究
常晓博1,2,赵款2,叶超2,姜成刚2,王淑杰2,蔡雪辉2,张辉1∗,安同庆2∗
(1.吉林农业大学动物科学技术学院,长春130118;2.中国农业科学院哈尔滨兽医研究所兽医生物技术国家重点实验室,哈尔滨150001)
为了解猪繁殖与呼吸综合征病毒(PRRSV)新分离株JL580遗传变异特征,研究扩增得到了该病毒的全基因组序列,并对GP5和NSP2序列进行了比对分析。结果表明:JL580株属于美洲株,且不同于中国现流行的其他毒株,与中国代表毒株CH-1a的核苷酸同源性是84.9%,与HP-PRRSV代表株JXA1、HuN4、TJ的核苷酸同源性分别是84.8%、84.9%、85%。对基因组的进一步分析表明,JL580株GP5蛋白与CH-1a、VR-2332、HuN4相比,R13→Q13,V27→A27,H38→N38,R151→K151;且NSP2存在3处缺失,与HP-PRRSV的30个氨基酸的缺失有明显的不同。JL580分离株的出现表明中国出现了新的PRRSV亚型,这对于PRRSV的防治和新疫苗的研制提供了科学依据。
猪繁殖与呼吸综合征病毒;JL580;基因组;遗传变异
猪繁殖与呼吸综合征(porcine reproductive and respiratory syndrome,PRRS)是由猪繁殖与呼吸综合征病毒(PRRS virus,PRRSV)引起的猪的一种急性、高度接触性传染病。PRRS于1987年[1]首次发生于美国的衣阿华、北卡罗来纳等州,随后迅速在美洲蔓延,我国于1996年[2]首次分离到PRRSV,目前该病在全球范围内暴发和流行[3]。PRRSV有两种基因型,分别为欧洲型(EU-type)和北美洲型(NA-type),我国主要流行毒株为北美洲型[4]。PRRSV在自然界中不断变异,不断出现新的病毒株,给PRRS的防治带来了巨大的挑战。PRRSV基因组中ORF5是高变区之一,其编码的蛋白是PRRSV主要的宿主保护性抗原蛋白[5],PRRSV的变异可以导致PRRS疫苗株交叉保护率低下。编码PRRSV的非结构蛋白基因中,位于ORF1a的NSP2基因最易发生变异,主要表现为氨基酸的突变、缺失和插入。2006年,我国南方暴发的HP-PRRSV基因组中NSP2存在30个不连续氨基酸的缺失[6];Darwich等[7]通过分析PRRSV基因I型毒株的基因和免疫多样性发现他们所研究的基因I型PRRSV中NSP2存在74个氨基酸的缺失。因此,针对GP5和NSP2的研究有助于了解整个基因组的遗传衍化特征。
2013年,吉林省某猪场发生疑似PRRS,临床表现为高热,耳朵发绀,24 h全身发绀,最后死亡,病程较短,发病率较高。通过病料采集、检测及病毒的分离鉴定,我们将新分离株命名为JL580。为弄清JL580新分离株的基因组变异特征,特别是ORF5和NSP2的变异规律,我们着重对JL580基因组中的GP5和NSP2进行了序列比对分析,从而了解JL580株的遗传变异情况。
1 材料和方法
1.1 样品和细胞 样品:采自吉林省某猪场疑似PRRSV感染猪的肺脏和淋巴结;细胞:Marc-145细胞由中国农业科学院哈尔滨兽医研究所动物疫病诊断及技术服务中心提供。
1.2 主要试剂 QIAamp Viral RNA Mini Kit购自QIAGEN公司;Reverse Transcriptase M-MLV、Recombinant RNase Inhibitor、TaKaRa EX Taq Hot Start Version、rTaq DNA聚合酶、3’-Full RACE Core Set Ver.2、Clontech SMARTerTMRACE cDNA Amplification Kit、pMD18-T Vector Kit、E.coli DH5a感受态均购自宝生物工程(TaKaRa)有限公司;胶回收试剂盒购自OMEGA公司。
1.3 全基因组分析
1.3.1 引物及参考序列 根据GenBank中已登录的国内外经典毒株序列的保守区及GP5测序结果,参考美国NADC30株[8](GenBank no.JN654459)设计并合成9对引物(表1)。

表1 JL580全基因组序列扩增所需引物
1.3.2 RNA提取与PCR扩增、克隆及测序 按照QIAamp Viral RNA Mini Kit操作手册提取病毒总RNA,反转录成cDNA后用于目的片段的扩增,基因5’端和3’端序列分别参照Clontech SMARTerTMRACE cDNA Amplification Kit和3’-Full RACE Core Set Ver.2使用说明书进行扩增。利用胶回收试剂盒回收PCR产物后分别与pMD18-T载体16℃过夜连接,转化并筛选重组阳性质粒,每个片段选取5个单克隆菌落,重复进行3次测序。
1.4 全基因组序列的分析 应用DNAStar软件中的Seqman程序拼接序列,Megalign进行序列比对及同源性分析;应用MEGA5.2软件对该毒株的全基因组序列进行遗传进化分析。
2 结果
2.1 JL580全基因组序列扩增结果 将JL580病毒提取的RNA反转录成cDNA后,利用9对引物对其进行扩增,得到9条与预期大小相符的片段(图1)。将测序所获得的9个首尾重叠的目的基因片段进行拼接,测定的基因组全长为15019 bp。

图1 PRRSV全基因组PCR扩增产物
2.2 JL580全基因组序列分析结果 遗传进化树(图2)结果显示,JL580株和NADC30株不同于CH-1a株、VR-2332株和HuN4株,它们形成了一个新的独立分支。JL580分离株与18株国内外参考株(表2)相比,核苷酸同源性在83.2%~92.9%,氨基酸同源性在70.8%~83.1%。与美国NADC30株相比核苷酸同源性为92.9%,氨基酸同源性为83.1%;与经典株CH-1a、VR-2332株的核苷酸同源性为84.9%和84.3%,推导的氨基酸同源性为74.1%和72.9%;与HP-PRRSV中的代表株HuN4、JXA1、TJ株的核苷酸同源性分别为84.9%、84.8%、85%,氨基酸同源性为75.7%、75.3%、75.7%。
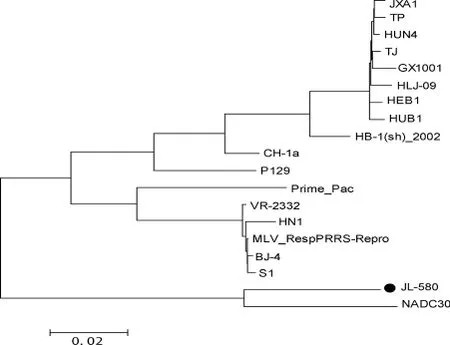
图2 PRRSV全基因组核苷酸序列的系统进化树
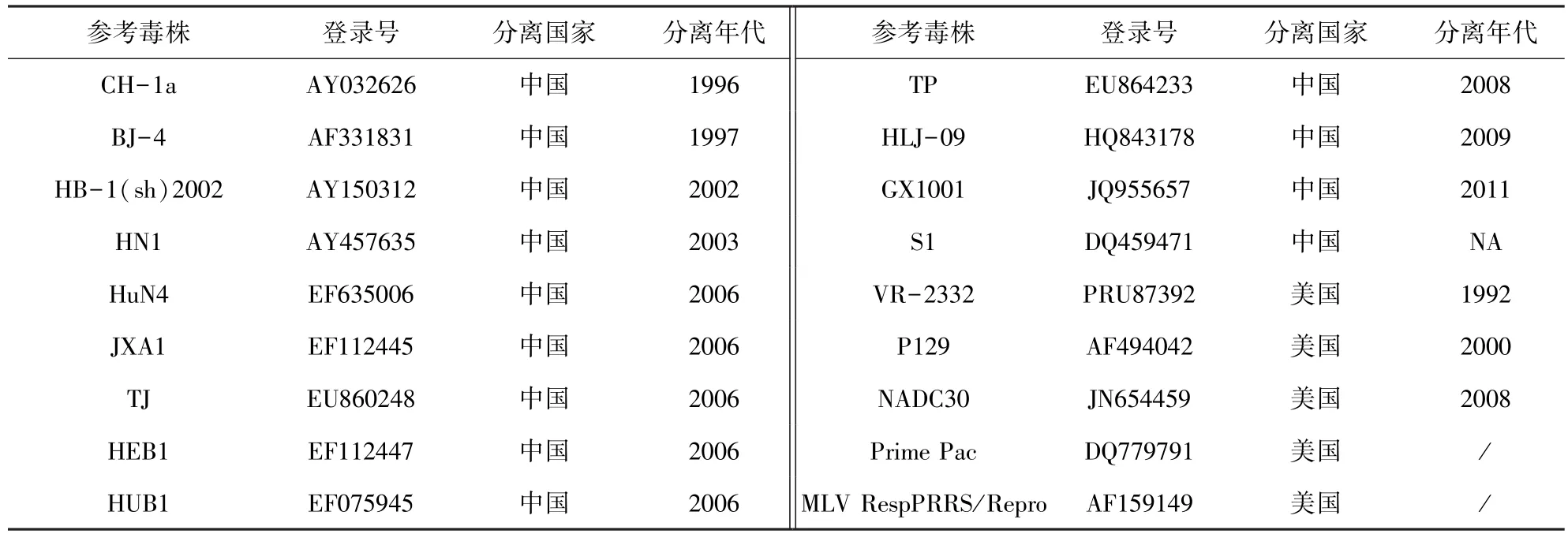
表2 参考毒株信息
2.3 JL580株ORF5遗传特征分析 JL580株与CH-1a株相比,GP5核苷酸同源性为87.4%,氨基酸同源性为86.1%,与HP-PRRSV中的HuN4毒株相比,核苷酸同源性为85.6%,氨基酸同源性为84.6%。此外,JL580病毒株GP5蛋白与CH-1a、VR-2332、HuN4相比发生了显著变异,主要变异位点分别是R13→Q13,V27→A27,H38→N38,R151→K151。
2.4 JL580株NSP2遗传特征分析 JL580株与经典株CH-1a株相比,NSP2基因核苷酸同源性为72.2%,氨基酸同源性为68.6%,与HuN4株相比核苷酸同源性为71.5%,氨基酸同源性为69.1%(表3)。JL580分离株存在3处连续碱基的缺失,分别位于基因组2304~2636、2779~2781和2852~2908处;这3处缺失导致NSP2编码的蛋白在322~432位缺失111个氨基酸、481位缺失1个氨基酸、504~522位缺失19个氨基酸(图3)。

表3 JL580株GP5和NSP2与部分毒株氨基酸同源性分析

图3 NSP2氨基酸序列比对
3 讨论
猪繁殖与呼吸综合征自1995年在我国发现以来已有20年,虽然已研制多种疫苗防治,但PRRS并未根除。PRRSV为单股正链RNA病毒[9],在自然界以及免疫压力[10]下不断变异。2006年北美[11]和亚洲出现的高致病性PRRSV表明病毒的潜在变异导致PRRSV出现了新的变异株。本研究通过对JL580分离株进行分子流行病学及遗传进化分析发现,JL580病毒株属于北美洲型PRRSV,并且与国内PRRSV经典株CH-1a和HP-PRRSV株HuN4处于遗传进化树的不同分支中,同源性较低;而与美国NADC30株的全基因组序列具有较高的同源性。从JL580与国内之前报道的经典PRRSV和HP-PRRSV的遗传距离上来看,JL580属于国内出现的一个新亚型毒株。本研究结果表明我国PRRSV出现了新的遗传分支,揭示了我国PRRSV存在着更为复杂的遗传多样性。
GP5是PRRSV的主要结构蛋白,也是最易发生变异的蛋白,R13和R151是毒力相关位点[12],非中和表位A(aa27~aa30)和中和表位B(aa37~aa45)是影响免疫的关键位点。研究发现JL580株GP5蛋白与CH-1a、VR-2332、HuN4相比,糖基化位点并未发生变异,但是中和表位、非中和表位及毒力位点发生了显著的变异,主要表现在R13→Q13,V27→A27,H38→N38,R151→K151。NSP2是PRRSV非结构蛋白中最易发生变异的蛋白,欧洲型和北美洲型NSP2高变区氨基酸缺失的报道[13]表明NSP2对于病毒的复制不具有决定性作用但在宿主免疫方面可能具有非常重要的作用。JL580株NSP2编码的蛋白存在3处不连续氨基酸的缺失,与经典株CH-1a相比NSP2共缺失了131个氨基酸;与2006年暴发的HP-PRRSV基因组中NSP2存在30个不连续氨基酸的缺失相比,JL580株NSP2基因编码的蛋白共缺失101个氨基酸,除此之外,JL580株和HuN4株在481位具有一个相同的缺失位点。另外,JL580病毒株NSP2的缺失情况与2006年Han等[11]和2014年Choi等[14]所研究的PRRSV分离株NSP2缺失131个氨基酸情况相似。因此,JL580病毒株是目前国内存在的一种新的变异株,其GP5和NSP2基因的变异相对于以往国内发现的变异株具有显著的差异性。至于JL580新分离株GP5中和表位、非中和表位及毒力位点的变异和基因组中3处不连续氨基酸的缺失是否会影响其免疫原性和致病性,还有待进一步研究。
[1] Keffaber K K.Reproductive failure of unknown etiology[J].Am Assoc Swine Pract Newslett,1989,(2):1-9.
[2] 郭宝清,陈章水,刘文兴,等.从疑似PRRSV感染的流产胎儿分离PRRSV的研究[J].中国畜禽传染病,1996,2:1-4.
[3] Rowland R R,Lunney J,Dekkers J.Control of porcine reproductive and respiratory syndrome(PRRS)through genetic improvements in disease resistance and tolerance[J].Front Genet,2012,3:260.
[4] 朱佳毅,任晓峰.猪繁殖与呼吸综合征病毒检测方法的研究进展[J].中国预防兽医学报,2014,36(1):77-80.
[5] Pirzadeh B,Dea S.Monoclonal antibodies to the ORF5 product of porcine reproductive and respiratory syndrome virus define linear neutralizing determinants[J].J Gen Virol,1997,78(8):1867-1873.
[6] Zhou Y J,Hao X F,Tian Z J,et al.Highly pathogenic porcine reproductive and respiratory syndrome virus emerged in China[J].Transbound Emerg Dis,2008,55(3/4):152-164.
[7] Darwich L,Gimeno M,Sibila M,et al.Genetic and immunobiological diversities of porcine reproductive andrespiratorysyndrome genotype I strains[J].Vet Microbiol,2011,150(1/2):49-62.
[8] Brockmeier S L,Loving C L,Vorwald A C,et al.Genomic sequence and virulence comparison of four type 2 porcine reproductive and respiratory syndrome virus strains[J].Virus Res,2012,169(1):212-221.
[9] Rossow K D.Porcine reproductive and respiratory syndrome[J].Vet Pathol,1998,35(1):1-20.
[10]Liu D,Zhou R,Zhang J,et al.Recombination analyses between two strains of porcine reproductive and respiratory syndrome virus in vivo[J].Virus Res,2011,155(2):473-486.
[11]Han J,Wang Y,Faaberg K S.Complete genome analysis of RFLP 184isolatesofporcinereproductiveandrespiratory syndrome virus[J].Virus Res,2006,122(1/2):175-182.
[12]Allende R,Kutish G F,Laegreid W,et al.Mutations in the genome of porcine reproductive and respiratory syndrome virus responsible for the attenuation phenotype[J].Arch Virol,2000,145(6):1149-1161.
[13]Rascon-Casteio E,Burgara-Estrella A,Mateu E,et al.Immunological features of the non-structural proteins of porcine reproductive and respiratory syndrome virus[J].Viruses,2015,7(3):873-886.
[14]Choi H W,Nam E,Lee Y J,et al.Genomic analysis and pathogenic characteristics of type 2 porcine reproductive and respiratory syndrome virus nsp2 deletion strains isolated in Korea[J].Vet Microbio,2014,170(3/4):232-245.
(编辑:李文平)
Genetic Diversity of Porcine Reproductive and Respiratory Syndrome Virus of New Subtype JL580 Strain
CHANG Xiao-bo1,2,ZHAO Kuan2,YE Chao2,JIANG Cheng-gang2,WANG Shu-jie2,CAI Xue-hui2,ZHANG Hui1∗,AN Tong-qing2∗
(1.College of Animal Science and Technology,Jilin Agricultural University,Changchun 130118,China;2.State Key Laboratory of Veterinary Biotechnology,Harbin Veterinary Research Institute,Chinese Academy of Agricultural Sciences,Harbin 150001,China)
To investigate the variation of pathogenic porcine reproductive and respiratory syndrome(PRRSV)of JL580,the full-length genomic sequence of the virus was amplified,and the GP5 and NSP2 gene were analyzed,respectively.Results showed that JL580 strain shared 84.9%nucleotide similarity of complete sequence with CH-1a strain,84.8%,84.9%,85%compared with JXA1,HuN4 and TJ strains,and clearly different from other prevalent strains in China,which belongs to North American genotype PRRSV.The GP5 protein sequence was variable in the newly isolated PRRSV when compared to CH-1a,VR-2332,HuN4,which were R13→Q13,V27→A27,H38→N38,R151→K151.In addition,the full-length genomic sequence of the virus revealed three distinct deletions in NSP2 which had significant difference with the 30 amino acid of HP-PRRSV.Therefore,the study of JL580 indicated that new subtype and new branch of PRRSV have emerged in China,which would provide scientific basis for the development of new vaccine and comprehensive prevention and control of PRRSV.
PRRSV;JL580;genome;genetic variation
2015-02-06
A
1002-1280(2015)05-0012-05
S858.28
国家自然科学基金(31270045);吉林省科技厅基础研究项目(20140101028JC)
常晓博,硕士研究生,从事PRRSV分子流行病学研究。
张辉,E-mail:huizhang01525@163.com;安同庆,E-mail:antongqing@163.com
