自噬参与低氧预处理抗PC12细胞糖氧剥夺损伤过程*
2015-04-27魏林郁王宝英李新娟李超堃白瑞樱李东亮新乡医学院基础医学院生理与神经生物学教研室河南新乡453003
卢 娜,魏林郁,王宝英,李新娟,李超堃,白瑞樱,李东亮(新乡医学院基础医学院生理与神经生物学教研室,河南新乡453003)
自噬参与低氧预处理抗PC12细胞糖氧剥夺损伤过程*
卢娜△,魏林郁,王宝英,李新娟,李超堃,白瑞樱,李东亮
(新乡医学院基础医学院生理与神经生物学教研室,河南新乡453003)
[摘要]目的:观察低氧预处理(HPC)对氧糖剥夺(OGD)损伤的PC12细胞的作用,同时探讨自噬在其中的作用。方法:培养的PC12细胞按照处理因素分为6组:对照组、HPC模型组、3-甲基腺嘌呤(3-MA)组、低氧预处理后氧糖剥夺(HPC+ OGD)组、3-甲基腺嘌呤预处理后行低氧预处理及氧糖剥夺(3-MA+ HPC+ OGD)组和OGD组。CCK-8检测细胞存活率; caspase-3活性检测试剂盒检测酶活性; TUNEL染色和流式细胞术检测细胞凋亡的情况; Western blot法检测自噬相关蛋白LC3、beclin-1和凋亡相关蛋白caspase-3的表达。结果:与对照组相比,OGD组细胞存活率明显降低;与3-MA+ HPC+ OGD组及OGD组相比,HPC+ OGD组细胞存活率明显升高(P<0.05)。与对照组相比,OGD组细胞的caspase-3酶活性明显升高;与3-MA+ HPC+ OGD组及OGD组相比,HPC+ OGD组的caspase-3酶活性明显降低(P<0.05)。与对照组相比OGD组细胞凋亡明显增多; HPC+ OGD组与OGD组相比凋亡明显减少(P<0.05)。此外,与对照组相比,OGD组的激活型caspase-3蛋白水平明显升高(P<0.05) ;且HPC+ OGD组与OGD组相比激活型caspase-3蛋白水平明显减少而LC3、beclin-1的蛋白水平明显升高(P<0.05)。结论: HPC抗OGD损伤的机制可能与其激活细胞自噬有关。
[关键词]自噬;细胞凋亡;低氧预处理;氧糖剥夺
[修回日期]2015-06-01
脑血管病以其高发病率、高死亡率、高致残率而严重危害人类健康,其中缺氧缺血性脑血管病约占70%。但遗憾的是,目前没有有效预防和治疗的方法。对其发病机制的研究及如何启动内源性保护机制以寻找有效的防治手段仍然是该领域研究的重点。低氧预处理(hypoxic preconditioning,HPC)是指1次或多次短暂、非致死性低氧刺激后,机体获得的对更严重甚至致死性缺血或缺氧的耐受性。HPC是机体抗缺氧或缺血的一种内源性保护现象,可发生在脑、心、肺、肝和肾等组织,其中脑HPC由Kitagawa等于1990年首报道,此保护现象为临床治疗脑缺血/低氧性损伤提供了新思路[1]。此后大量实验及我实验室前期工作表明,HPC能改善以后出现的脑缺氧缺血[2-3]。若预先多次给于人体能耐受的低氧条件,则可明显减少缺血性脑卒中的发病率、死亡率和致残率。目前,脑低氧预处理的机制尚不十分清楚,尤其是其细胞保护的信号通路、分子机制还需要深入研究。本实验以PC12细胞为对象,从不同层面对HPC抗脑缺氧缺血的机制进行研究,探索自噬在其中的作用。
材料和方法
1材料
PC12细胞株由上海中科院典型细胞培养库提供; DMEM培养液和新生牛血清购于HyClone; 3-甲基腺嘌呤(3-methyladenine,3-MA)购于Sigma;细胞存活与毒性检测试剂盒CCK-8、caspase-3活性检测试剂盒、Annexin V-FITC细胞凋亡检测试剂盒和一步法TUNEL细胞凋亡检测试剂盒均购自碧云天生物技术有限公司;兔抗LC3-Ⅱ多克隆抗体购自Abcam;兔抗beclin-1多克隆抗体购自Bioworld;兔抗cleaved-caspase-3单克隆抗体购自Abgent;辣根过氧化物酶标记山羊抗兔IgG(H+ L)购自Laboratories;蛋白Marker购自Fermentas。
2主要方法
2.1PC12细胞培养细胞置于10%胎牛血清的达尔伯克改良伊格尔培养基(Dulbecco’s modified eagle medium,DMEM)中,在37℃、5% CO2的饱和水汽培养箱中培养,每隔48 h换培养液,当单层细胞融合后,行传代处理。传代后随机分为对照(control)组、HPC组、3-MA组、HPC+氧糖剥夺(oxygen-glucose deprivation,OGD)组、3-MA+ HPC+ OGD组和OGD组。每次每组设6个平行复孔,实验重复3次。
2.2PC12细胞低氧预处理模型的建立每天定时将PC12细胞移至37℃恒温密闭容器内,连续充以低氧气体(93% N2、2% O2、5% CO2),30 min后立即取出,再恢复常氧培养,重复6次,建立细胞低氧预处理模型。
2.3PC12细胞OGD模型的建立选用培养6 d生长良好的PC12细胞,置换成无糖培养基孵育,放入
37℃厌氧培养箱中培养2 h(94% N2、1% O2、5% CO2),然后复氧复糖培养24 h。
2.4CCK-8比色法检测细胞存活率取对数生长期细胞,调整细胞密度,以2×104/L接种于96孔板,每孔细胞悬液100 μL。在37℃、5% CO2的饱和水汽培养箱中培养24 h。处理后,每孔加10 μL CCK-8溶液,避光孵育2 h,酶标仪在波长450 nm处读取吸光度(A)值,用8个复孔A值的均数计算存活率。存活率按下式计算:存活率=A处理/A对照×100%。本实验对照组的细胞存活率定为100%。
2.5Caspase-3活性检测试剂盒检测caspase-3酶活性冰浴裂解细胞15 min,4℃、20 000×g离心10 min,将上清转移到96孔板中。配置反应体系,再加入Ac-DEVD-pNA后混匀,37℃孵育60 min,测定A405。通过标准曲线对比计算出样品中催化产生多少量的pNA。
2.6一步法TUNEL细胞凋亡检测试剂盒检测凋亡取对数生长期细胞,调整细胞密度,以5×105/L接种于24孔板内载玻片上,每个孔细胞悬液500 μL;培养24 h后,分组处理细胞;弃培养液、PBS洗涤1次、4%多聚甲醛固定1 h、0.1% Triton X-100冰浴孵育5 min、在样品上加50 μL TUNEL检测液,37℃避光孵育60 min,封片后立即在荧光显微镜下观察。在100倍镜下,随机取5个视野,计数呈现绿色荧光的凋亡细胞,取平均值,计算细胞凋亡率(%)=凋亡细胞数/视野下细胞总数×100%,重复3次。
2.7流式细胞术检测细胞凋亡率PC12细胞经分组处理后,PBS洗2遍,收集104个细胞,加入195 μL Annexin V-FITC结合液轻轻重悬细胞,加入5 μL Annexin V-FITC,轻轻混匀,加入10 μL碘化丙啶染色液,轻轻混匀。室温(20~25℃)避光孵育10~20 min,随即进行流式细胞仪检测细胞凋亡率。
2.8Western blot法检测caspase-3、LC3-Ⅱ和beclin-1的蛋白水平收获细胞,4℃预冷的PBS洗1次,加入适量的细胞裂解液,用细胞刮刮下细胞,低温离心机12 000×g离心10 min,取上清液,弃沉淀。用BCA法测定各组蛋白含量,每孔上样25 μL蛋白,经15% SDS-PAGE 80V分离2 h后,300 mA恒流转膜40 min,5%脱脂牛奶常温下封闭1 h后,在4℃孵育I抗(兔抗大鼠LC-3滴度为1∶1 000,兔抗大鼠bec-lin-1滴度为1∶1 500,兔抗大鼠cleaved caspase-3抗体滴度为1∶1 000,小鼠抗大鼠β-actin滴度为1∶3 000) ; 5%脱脂牛奶常温下封闭; II抗(羊抗兔辣根过氧化物酶IgG和羊抗小鼠辣根过氧化物酶IgG滴度均为1∶5 000,5%脱脂牛奶配制)孵育,ECL显影显示目的条带和内参照。用ImageJ软件分析光密度值,Sigma Plot 13软件分析蛋白表达的相对水平。
3统计学处理
用SPSS 18.0统计软件进行分析。数据均采用均数±标准差(mean±SD)表示,多组间比较采用单因素方差分析(one-way ANOVA),组间两两比较采用Student-Newman-Keuls检验(SNK-q检验)。以P<0.05为差异有统计学意义。
结果
1不同处理组PC12细胞的形态观察
倒置相差光学显微镜下观察PC12细胞形态的变化,正常PC12细胞大部分呈梭形,少数呈多边形,多数可见细胞突起,折光性好,贴壁能力强; OGD处理后,细胞贴壁数量明显减少,细胞形态逐渐变为圆形,体积皱缩,突起减少或消失,折光性减弱,脱壁的细胞增加; HPC+ OGD组细胞的缺氧缺糖损伤明显减轻; 3-MA+ HPC+ OGD组细胞的损伤无明显减轻,见图1。这提示HPC能减轻OGD引起的细胞损伤;自噬阻断剂3-MA可阻断这种保护作用。
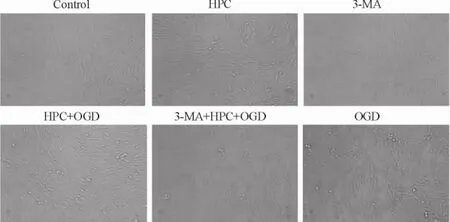
Figure 1.The effect of different treatment factors on the morphology of PC12 cells (×100).图1不同处理因素对PC12细胞形态的影响
2不同处理因素对PC12细胞存活率的影响
CCK-8法检测结果以空白对照组细胞存活率为100.0%,则HPC、3-MA、HPC+ OGD、3-MA+ HPC+ OGD和OGD组存活率分别为98.7%、97.9%、86.2%、63.5%和67.6%。HPC+ OGD、3-MA+ HPC+ OGD组、OGD组与对照组比较差异均有统计学意义(P<0.05) ; HPC+ OGD组与3-MA+ HPC+ OGD组及OGD组比较差异均有统计学意义(P<0.05),见图2。
3不同处理因素对PC12细胞caspase-3酶活性的影响
Caspase-3活性检测试剂盒检测结果以空白对照组细胞的caspase-3酶活性为1,则HPC、3-MA、HPC+ OGD、3-MA+ HPC+ OGD和OGD组的caspase-3酶活性分别为1.15、1.09、2.56、3.38和3.42。HPC+ OGD、3-MA+ HPC+ OGD组、OGD组与对照组比较差异均有统计学意义(P<0.05) ; HPC+ OGD组与3-MA+ HPC+ OGD组及OGD组比较差异均有统计学意义(P<0.05),见图3。
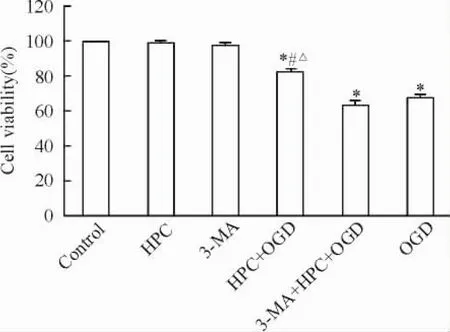
Figure 2.The effect of different treatment factors on the cell viability of PC12 cells.Mean±SD.n=3.*P<0.05 vs control group;#P<0.05 vs OGD group;△P<0.05 vs 3-MA+ HPC+ OGD group.图2不同处理因素对PC12细胞存活率的影响
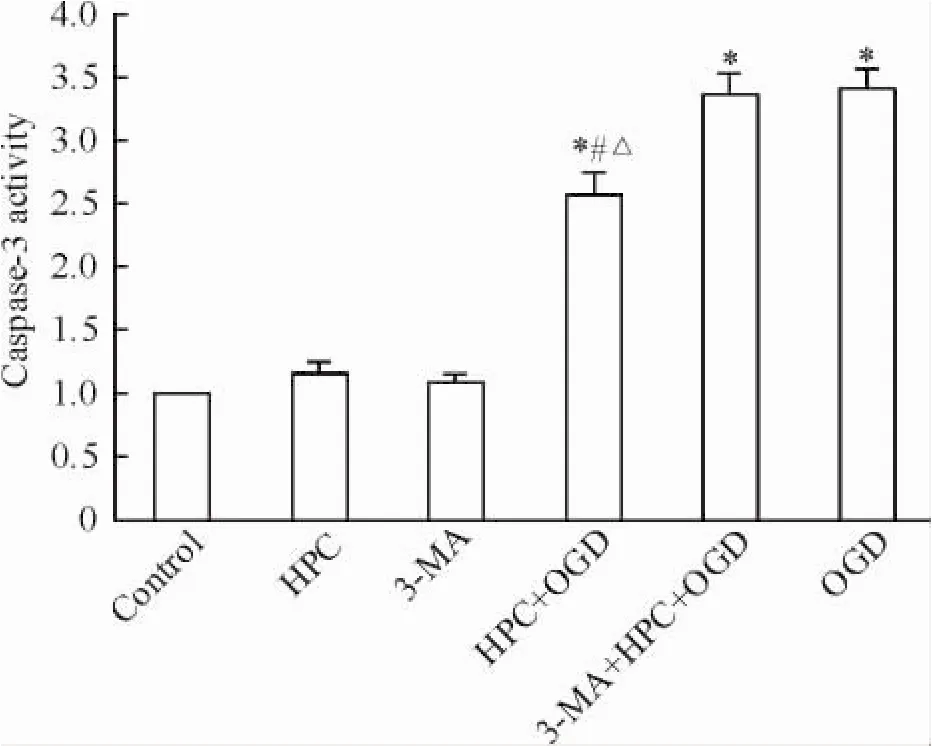
Figure 3.The caspase-3 activity in the PC12 cells with different treatments.Mean±SD.n=3.*P<0.05 vs control group;#P<0.05 vs OGD group;△P<0.05 vs 3-MA+ HPC+ OGD group.图3各组细胞caspase-3酶活性的比较
4 TUNEL法荧光染色检测细胞凋亡率
TUNEL荧光染色可见control组、HPC组和3-MA组阳性凋亡细胞数较少; HPC+ OGD组、3-MA+ HPC+ OGD组及OGD组阳性细胞数明显增多; HPC+ OGD组明显低于3-MA+ HPC+ OGD组及OGD组(P<0.05),见图4。
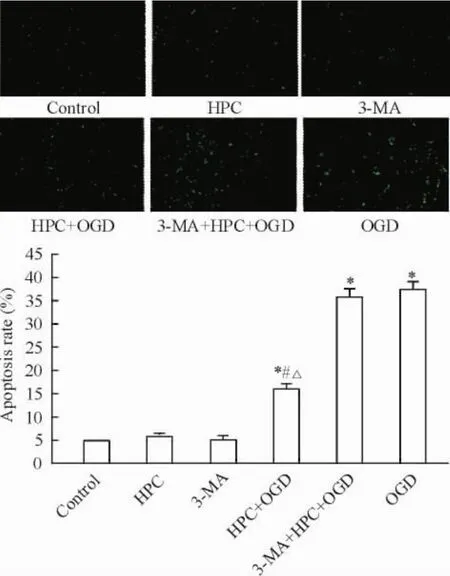
Figure 4.The TUNEL staining and apoptotic rate in the PC12 cells with different treatments (×100).Mean±SD.n=3.*P<0.05 vs control group;#P<0.05 vs OGD group;△P<0.05 vs 3-MA+ HPC+ OGD group.图4各组细胞的TUNEL荧光染色及凋亡率的比较
5流式细胞术检测细胞凋亡率
Annexin V-FITC/PI双染流式细胞仪检测结果显示control组、HPC组、HPC+ OGD组和OGD组的凋亡率分别为(3.40±0.36) %、(3.87±0.59) %、(18.67±0.97) %和(36.40±0.89) %;与正常对照组相比,HPC+ OGD组和OGD组凋亡率明显增加(P<0.05),HPC+ OGD组与OGD组比较明显减少(P<0.05),见图5。
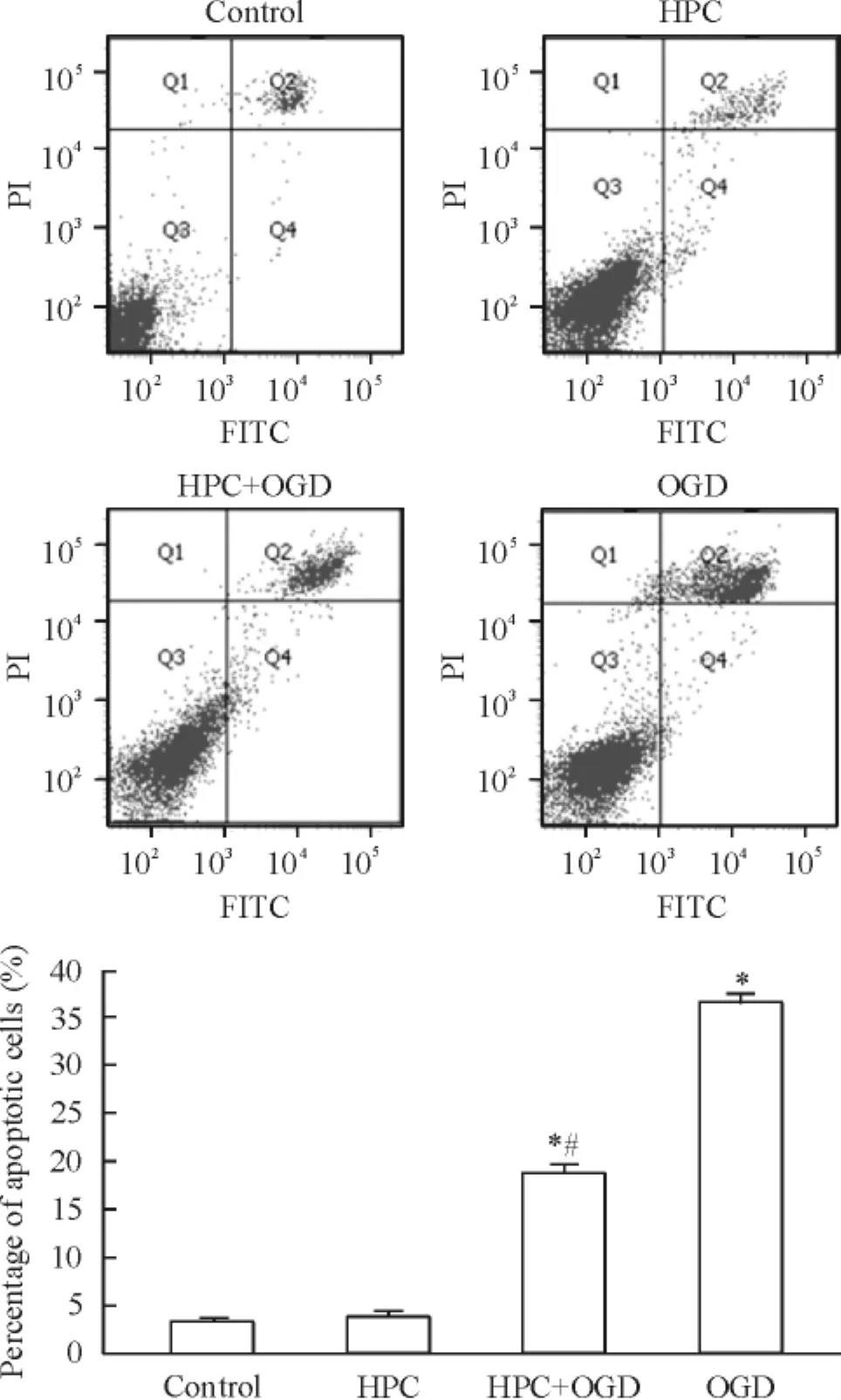
Figure 5.Apoptosis of PC12 cells detected by flow cytometry of Annexin V-FITC/PI staining.Mean±SD.n=3.*P<0.05 vs control group;#P<0.05 vs OGD group.图5流式细胞术检测细胞凋亡
6 Western blot法检测caspase-3、LC3-Ⅱ和beclin-1蛋白水平的变化
蛋白质印迹分析结果显示,与正常对照组比较,HPC+ OGD组和OGD组的cleaved caspase-3的蛋白量均增加(P<0.05) ;且HPC+ OGD组明显低于OGD组(P<0.05)。HPC+ OGD组的LC3-Ⅱ和beclin-1蛋白表达明显高于OGD组(P<0.05),见图6。
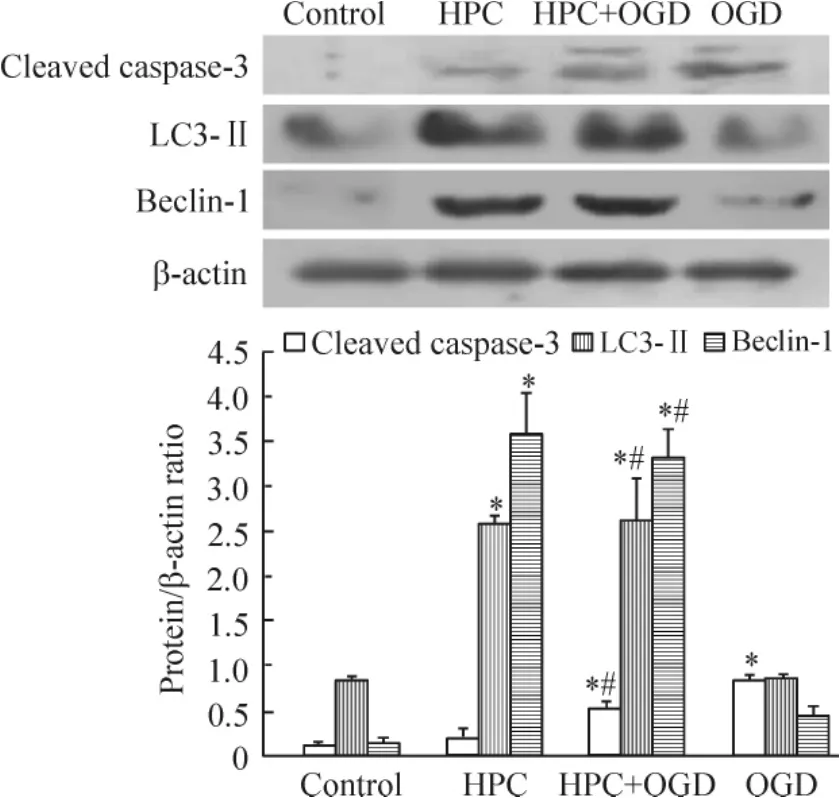
Figure 6.The protein levels of cleaved caspase-3,LC3-Ⅱand beclin-1 in the PC12 cells with different treatments.Mean±SD.n=3.*P<0.05 vs control group;#P<0.05 vs OGD group.图6各组PC12细胞cleaved caspase-3、LC3-Ⅱ和beclin-1蛋白水平的比较
讨论
自噬是指存在于真核细胞中的生命现象,细胞在自噬相关基因的调控下形成自噬体,并与溶酶体融合形成自噬溶酶体,降解其所包裹的内容物,以实现细胞本身的代谢需要和某些细胞器的更新[4]。自噬是继坏死、凋亡后发现的第3种细胞死亡形式,参与了如脑损伤等多种疾病的发病过程[4]。细胞在受到外界因素刺激,如饥饿、缺氧、高温、应激等逆境条件下均可诱导细胞自噬的发生,可以在一定程度上对细胞进行保护[5-6]。自噬在机体的生命过程中随处可见,但是其关键的发生和作用机制尚未完全阐明[7]。尽管自噬现象很早就被发现,但对自噬与脑缺氧缺血关系的研究时间并不长。目前,大多数研究证实自噬具有脑保护作用。Erlich等[8]采用雷帕霉素干预大鼠颅脑损伤也发现了自噬现象的产生,证明颅脑损伤后4 h注入雷帕霉素在脑功能恢复上与未注入组有明显改善效果。Carloni等[9]研究结果显示细胞凋亡与自噬存在负相关,自噬激动剂雷帕霉素的使用加速后自噬现象的发生,减少细胞凋亡,减轻脑损伤;说明自噬激活在脑损伤早期,可能对神经细胞起着一定的保护作用。Zhang等[10]证明脑外伤后确实激活了细胞自噬的发生,且能够维持32天,自噬在损伤后早期主要保护周围神经细胞免于凋亡和退行性变,并在神经细胞损伤与修复过程中长期发挥生物学作用。Komatsu等[11]的研究表明一定程度自噬具有增强抗氧化、恢复认知功能、调节脑血流量等特异的作用。当神经元遭遇应激时,自噬最先被激活,当自噬不能维持神经元稳态后,细胞再继发凋亡,最后进入坏死;且自噬与凋亡可同时存在于神经元细胞,推测自噬参与脑缺氧缺血后神经元的保护[12-13]。事实上,自噬是一个动态过程,主要由自噬相关基因编码的蛋白来完成相应的生物学功能。LC3是哺乳动物细胞中酵母ATG8(Aut7/Apg8)基因的同源物,定位于前自噬泡和自噬泡膜表面,参与自噬体的形成。LC3有Ⅰ型和Ⅱ型之分,未发生自噬时,细胞内合成的LC3经过加工,成为胞质可溶性的LC3-Ⅰ,常规表达。当自体吞噬发生时,LC3-Ⅰ经泛素样加工修饰过程,与自噬膜表面的磷脂酰乙醇胺结合,形成LC3-Ⅱ。LC3-Ⅱ结合并始终位于胞内自噬体的膜上,其含量的多少与自噬泡数量的多少成正比。因此LC3-Ⅱ是目前观察自噬现象是否存在、研究自噬活性较为可靠的生物学标志物[14]。Beclin-1是激活自噬的上游信号分子,游离的beclin-1能与多种蛋白质共同形成Ⅲ型PI3K复合体,参与早期自噬体的形成。已经证实通过上调beclin-1在哺乳动物细胞中的表达能够刺激自噬的发生[15]。因此,常以beclin-1的表达来衡量自噬的发生。
凋亡是由基因控制的细胞主动死亡的一种过程。Caspase是一大类凋亡调控基因,对神经元的凋亡起着非常关键的作用,而caspase-3被称为死亡蛋白酶,是caspase家族中最重要的细胞凋亡执行者之一[16]。脑组织缺血、缺氧后,神经细胞内能量代谢发生障碍,产生大量氧自由基、兴奋性氨基酸及内质网应激素等,均可诱导细胞自噬的发生。值得注意的是,细胞自噬是一把双刃剑。一方面,自噬的过度激活可能介导细胞程序性死亡的发生[17-18];另一方面,自噬的启动也有助于维持神经元的稳态,减少继发损伤,并有效清除已经被损伤的细胞器而对神经元起保护作用[19]。有报道称低氧预处理可诱导自噬,自噬可能参与神经保护作用[20]。那么在PC12细胞OGD损伤过程中自噬扮演什么角色?在HPC的影响下,自噬能否被激活?细胞自噬的增强能否对更严重的缺氧缺血起到保护作用?检索国内外文献未见相关报道。
本研究CCK-8结果表明,OGD组细胞存活率显著降低,HPC+ OGD组细胞存活率显著高于OGD组,3-MA+ HPC+ OGD组显著低于HPC+ OGD组。这提示HPC明显增加OGD导致的PC12细胞存活率降低,具有保护作用;且可被自噬抑制剂3-MA阻断。通过caspase-3酶活性检测显示,OGD组caspase-3酶活性显著升高,HPC+ OGD组的caspase-3酶活性显著低于OGD组,且3-MA+ HPC+ OGD组显著高于HPC+ OGD组。提示HPC明显降低OGD导致PC12细胞caspase-3酶活性升高,具有保护作用;且可被自噬抑制剂3-MA阻断。通过TUNEL荧光染色检测法及流式细胞术检测细胞凋亡率显示,与正常对照组相比,OGD组细胞凋亡率明显增多,HPC+ OGD组明显低于3-MA+ HPC+ OGD组及OGD组。提示HPC明显降低了OGD导致的PC12细胞凋亡率,具有保护作用,与相关研究结果基本相符[21-22],且这种保护效应可被自噬抑制剂3-MA阻断。我们进一步通过Western blot对凋亡相关蛋白caspase-3及自噬相关蛋白LC3-Ⅱ、beclin-1的表达进行检测,发现HPC+ OGD组、OGD组的激活型caspase-3蛋白量均增加,且HPC+ OGD组明显低于OGD组。HPC组及HPC+ OGD组LC3-Ⅱ和beclin-1蛋白表达明显高于OGD组。说明细胞自噬的激活在一定程度上可对抗凋亡,LC3-Ⅱ及beclin-1蛋白的调控和caspase-3活化存在负相关。因此,我们推测HPC诱导自噬的激活,可对抗神经元的氧糖剥夺损伤,延缓细胞凋亡过程的发生,对细胞存活起一定的保护作用[23]。研究低氧预处理启动和调控自噬的激活,或许能够为应对神经元损伤提供新的策略,为脑缺血的防治开辟新的途径,既有理论意义,又有潜在的临床应用前景。
[参考文献]
[1]Ara J,Fekete S,Frank M,et al.Hypoxic-preconditioning induces neuroprotection against hypoxia-ischemia in newborn piglet brain[J].Neurobiol Dis,2011,43(2) : 473-485.
[2]Zhang N,Yin Y,Han S,et al.Hypoxic preconditioning induced neuroprotection against cerebral ischemic injuries and its cPKC-gamma-mediated molecular mechanism[J].Neurochem Int,2011,58(6) : 684-692.
[3]Sun HS,Xu B,Chen W,et al.Neuronal K(ATP) channels mediate hypoxic preconditioning and reduce subsequent neonatal hypoxic-ischemic brain injury[J].Exp Neurol,2015,263: 161-171.
[4]Shpcilka T,Elazar Z.Shedding light on mammalian microautophagy[J].Dev Cell,2011,20(1) : 1-2.
[5]Zou W,Wang X,Vale R,et al.Autophagy genes promote apoptotic cell corpse clearance[J].Autophagy,2012,8(8) : 34-38.
[6]Luo J.Autophagy and ethanol neurotoxicity[J].Autophagy,2014,10(12) : 2099-2108.
[7]Aveleira CA,Botelho M,Carmo-Silva S,et al.Neuropeptide Y stimulates autophagy in hypothalamic neurons[J].Proc Natl Acad Sci U S A,2015,112 (13) : E1642-E1651.
[8]Erlich S,Alexandrovich A,Shohami E,et al.Rapamycin is a neuroprotective treatment for traumatic brain injury [J].Neurobiol Dis,2007,26(1) : 86-93.
[9]Carloni S,Buonocore G,Balduini W.Protective role of autophagy in neonatal hypoxia-ischemia induced brain injury[J].Neurobiol Dis,2008,32(3) : 329-339.
[10]Zhang YB,LI SX,Chen XP,et al.Autophagy is activated and might protect neurons from degeneration after traumatic brain injury[J].Neurosci Bull,2008,24(3) : 143-149.
[11]Komatsu M,Waguri S,Chiba T,et al.Loss of autophagy in the central nervous system causes neurodegeneration in mice[J].Nature,2006,441(7095) : 880-884.
[12]Wang P,Guan YF,Du H,et al.Induction of autophagy contributes to the neuroprotection of nicotinamide phosphoribosyltransferase in cerebral ischemia[J].Autophagy,2012,8(1) : 77-87.
[13]Yamamoto A,Yue Z.Autophagy and its normal and pathogenic states in the brain[J].Annu Rev Neurosci,2014,37: 55-78.
[14]Mehrpour M,Esclatine A,Beau I,et al.Overview of macro-autophagy regulation in mammalian cells[J].Cell Res,2010,20(7) : 748-762.
[15]Gabryel B,Kost A,Kasprowska D.Neuronal autophagy in cerebral ischemia is a potential target for neuroprotective strategies[J].Pharmacol Rep,2012,64(1) : 1-15.
[16]Jan CR,Su JA,Teng CC,et al.Mechanism of maprotiline-induced apoptosis: role of[Ca2+]i,ERK,JNK and caspase-3 signaling pathways[J].Toxicology,2013,304: 1-12.
[17]Adhami F,Liao G,Morozov YM,et al.Cerebral ischemia-hypoxia induces intravascular coagulation and autophagy [J].Am J Pathol,2006,169(2) : 566-583.
[18]Balduini W,Carloni S,Buonocore G,et al.Autophagy in hypoxia-ischemia induced brain injury[J].J Matern Fetal Neonatal Med,2012,25 Suppl 1: 30-34.
[19]Zhao Y,Chen G,Zhang W,et al.Autophagy regulates hypoxia-induced osteoclastogenesis through the HIF-1α/BNIP3 signaling pathway[J].J Cell Physiol,2012,227 (2) : 639-648.
[20]Tzeng YW,Lee LY,Chao PL,et al.Role of autophagy in protection afforded by hypoxic preconditioning against MPP+-induced neurotoxicity in SH-SY5Y cells[J].Free Radic Biol Med,2010,49(5) : 839-846.
[21]周天恩,张萌,杨正飞,等.亚低温减轻氧糖剥夺所致的大鼠海马神经元损伤[J].中国病理生理杂志,2013,29(7) : 1165-1170.
[22]Cui C,Zhou T,Li J,et al.Proteomic analysis of the mouse brain after repetitive exposure to hypoxia[J].Chem Biol Interact,2015,236: 57-66.
[23]Sarkar C,Zhao Z,Aungst S,et al.Impaired autophagy flux is associated with neuronal cell death after traumatic brain injury[J].Autophagy,2014,10(12) : 2208-2222.
(责任编辑:林白霜,余小慧)
Hypoxic preconditioning alleviates oxygen-glucose deprivation in PC12 cells: the involvement of autophagy
LU Na,WEI Lin-yu,WANG Bao-ying,LI Xin-juan,LI Chao-kun,BAI Rui-ying,LI Dong-liang
(Department of Physiology and Neurobiology,Xinxiang Medical University,Xinxiang 453003,China.E-mail: 13462225817@163.com)
[ABSTRACT]AIM: To examined the effects of hypoxic preconditioning (HPC) on oxygen-glucose deprivation (OGD) -induced PC12 cells,and to investigate its possible mechanisms of autophagy.METHODS: Cultured PC12 cells were randomly divided into control group,HPC group,3-methyladenine (3-MA) group,HPC+ OGD group,3-MA+ HPC+ OGD group and OGD group.CCK-8 assay was used to detect the cell viability.The caspase-3 activity was also tested.TUNEL staining and flow cytometry were used to detect the cell apoptosis.The protein levels of apoptosis-related protein caspase-3 and autophagy-marked protein LC3-2 and beclin-1 were determined by Western blot.RESULTS: Compared with control group,the viability of PC12 cells was significantly reduced,and the activity of caspase-3 was significantly increased in OGD group.Compared with 3-MA+ HPC+ OGD group and OGD group,the viability of PC12 cells was significantly increased,and the activity of caspase-3 was significantly reduced in HPC+ OGD group (P<0.05).The PC12 cell injury was apparent after OGD with a great increase in the apoptotic rate (P<0.05).Compared with OGD group,the apoptotic rate significantly decreased in HPC+ OGD group (P<0.05).Compared with control group,the protein level of cleaved caspase-3 was significantly increased in OGD group (P<0.05).Compared with OGD group,the protein level of cleaved caspase-3 was significantly decreased,and the levels of LC3-2 and beclin-1 were significantly increased in HPC+ OGD group (P<0.05).CONCLUSION: OGD decreases cell survival and induces apoptosis.Activation of cell autophagy may be the mechanism by which hypoxic preconditioning protects the PC12 cells from OGD induced injury.
[KEY WORDS]Autophagy; Apoptosis; Hypoxic preconditioning; Oxygen-glucose deprivation
通讯作者△Tel: 0373-3029104; E-mail: 13462225817@163.com
*[基金项目]河南省教育厅科学技术研究重点项目(No.14A310009; No.14A310019)
[收稿日期]2015-03-27
[文章编号]1000-4718(2015)09-1627-06
[中图分类号]R363.21
[文献标志码]A
doi:10.3969/j.issn.1000-4718.2015.09.017
