B7家族在非霍奇金淋巴瘤中的表达及意义
2015-04-27韩丽娟慕竹青周林静孟娜娜马晓艳齐路霞项红霞何秋立张桂芳
鲁 一,韩丽娟,慕竹青,周林静,孟娜娜,马晓艳,齐路霞,项红霞,何秋立,张桂芳
(新乡市中心医院肿瘤内科,新乡 453000)
非霍奇金淋巴瘤(non-Hodgkin’s lymphoma,NHL)是恶性淋巴瘤中较常见的一大类型,主要发生于淋巴结,也可发生于结外淋巴组织和非淋巴组织,其肿瘤行为从惰性到侵袭性不等。预后较差的高度恶性淋巴瘤在我国发病率较高,约占NHL的30%[1]。NHL是一种异质性较强的疾病,根据不同的淋巴细胞起源,可分为B细胞淋巴瘤(B-NHL)、T细胞淋巴瘤(T-NHL)和NK/T细胞淋巴瘤(NKTL)。除少数类型外,T-NHL及NKTL较B-NHL侵袭性强、预后差。NHL患者往往存在着免疫抑制或免疫紊乱,我们考虑NHL可能通过某种机制逃避了机体的免疫监视和杀伤。而免疫逃逸机制的差异可能在决定NHL不同肿瘤类型中起到了关键作用。PD-L1和PD-L2是B7家族共刺激分子的重要成员。研究发现[2,3],多种恶性肿瘤细胞通过高表达PD-Ls,与其受体PD-1(programmed death-1,PD-1)结合传递负性调控信号,从而诱导肿瘤抗原特异性T细胞的凋亡和免疫无能,促使肿瘤的免疫逃逸。我们考虑NHL可能通过高表达B7家族,与其受体PD-1相互作用,从而介导肿瘤的免疫逃逸。本文主要检测PD-L1和PD-L2在NHL中的表达,并探讨其意义,旨在为NHL各淋巴瘤亚型寻找新的免疫治疗靶点。
1 材料与方法
1.1 试剂
RPMI 1640培养基购自Solarbio生物公司;无支原体胎牛血清(fetal booine serum,FBS)购自杭州四季青公司;人AB血清由河南省中心血站提供;非必需氨基酸购自Thermo公司;人淋巴细胞分离液购自天津灏洋生物公司;RNA提取试剂盒购自北京天根生物公司;逆转录试剂盒购自Fermentas;引物均由上海生工合成;SYBR Green Mixture购自北京康为世纪。
1.2 细胞系及培养
NKTL细胞系SNK-6及SNK-8由日本千叶大学Norio Shimizu教授惠赠,培养于含人AB血清(10%)和IL-2(1 000U/ml)的RPMI 1640培养基中。YTS细胞系由美国Mayo医学中心Scott Kaufmann教授惠赠,培养于含非必需氨基酸(1%)和FBS(10%)的RPMI 1640培养基中。急性T淋巴细胞白血病细胞系Jurkat、间变大细胞淋巴瘤细胞系Karpas-299、皮肤T-NHL细胞系Hut-78、B-NHL细胞系LY-1和LY-8、Burkitt’s淋巴瘤细胞系Ramos均购自中国科学院上海细胞生物研究所,均用含FBS(10%)的RPMI 1640培养基培养。所有细胞系均置于37ºC、5% CO2的细胞培养箱中培养,每2~3d换液。取对数生长期的细胞进行实验。
1.3 外周血单个核细胞的提取
收集健康志愿者新鲜外周血,肝素抗凝,用等体积的磷酸盐缓冲液(phosphate buffered saline,PBS;pH=7.2)稀释;吸取2倍体积的淋巴细胞分离液置于离心管中,毛细滴管吸取已稀释的血标本沿管壁缓慢加入分离液面上(切勿冲破液面),3 000转/min离心20min;然后吸取中间白色云雾层,所得即为外周血单个核细胞(peripheral blood monocyte cells,PBMCs),经PBS洗涤,2 000转/min离心5min后弃上清,重复操作3次,最后用少量PBS重悬。
1.4 总RNA的提取及逆转录
取5×106个/ml对数生长期的细胞,操作按离心柱法RNA提取试剂盒说明书,收集RNA约70µl。经纯度检测所有样品的D260/D280均在1.85~2.00之间。逆转录操作按Fermentas逆转录试剂盒程序,反应条件:42℃、60min,70℃、5min,4℃、暂停。所得cDNA保存于-20℃冰箱备用。
1.5 实时荧光定量PCR检测
实时荧光定量PCR(real time fluorescence quantitative PCR,RQ-PCR)检测B7家族PD-L1和PD-L2 mRNA在B-NHL、T-NHL及NKTL细胞系中的表达水平。按SYBR Green Mixture试剂盒说明书操作。以PBMCs的β-actin基因为内参照,引物序列如下。PD-L1:F1 CTT CTT ATT ATG CCT TGG TGT TGC A,R1 GGA ATA CGT CTC CTC CAA ATG TG;PD-L2:F1 CAA CAT GGC TGC TTC ACA TTT T,R1 TGT GGT GAC AGG TCT TTT TGT TGT;β-actin:F1 TCA CGT GGA CAT CCG CAA AG,R1 CTG GTA GGT GGA CAG GG。扩增产物分别为146bp,110bp及205bp。反应条件:预变性95℃、10min,变性95℃、15s,变性/延伸60℃、1min,40个循环。溶解曲线分析:95℃、15s,60℃、1min,95℃、15s,60℃、15s,得到相应Ct值,应用ABI 7500 Fast System软件进行分析,实验重复3次。
1.6 统计学处理
采用SPSS17.0软件进行统计学分析。计量资料以均数±标准差(±s)表示,组间比较采用单因素方差分析。P<0.05为差异有统计学意义。
2 结 果
2.1 PD-L1和PD-L2 mRNA的扩增曲线和溶解曲线
应用RQ-PCR对NHL各细胞系PD-L1和PD-L2 mRNA的表达进行相对定量分析,获得单峰溶解曲线。峰的形状锐利,显示扩增产物为特异序列(图1)。
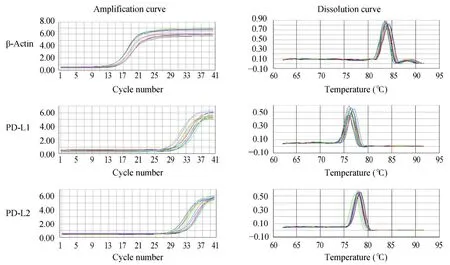
图1 NHL各细胞系中PD-L1和PD-L2 mRNA的扩增曲线和溶解曲线Figure 1 The amplification curves and dissolution curves of mRNA of PD-L1 and PD-L2 in NHL cell lines
2.2 NHL各细胞系中PD-L1、PD-L2 mRNA的相对表达量
以正常PBMCs为对照,NKTL细胞系(SNK-6、SNK-8和YTS)和T-NHL细胞系(Jurkat、Karpas-299和Hut-78)中,PD-L1和PD-L2 mRNA表达量均明显高于正常PBMCs(P<0.05)。B-NHL细胞系(LY-1、LY-8和Ramos)中,PD-L1和PD-L2 mRNA表达量与正常PBMCs无明显差异(P>0.05)。详见图2和图3。

图2 NHL各细胞系中PD-L1 mRNA的相对表达量Figure 2 The relative expression of PD-L1 mRNA in NHL cell lines (n=3, ±s )

图3 NHL各细胞系中PD-L2 mRNA的相对表达量Figure 3 The relative expression of PD-L2 mRNA in NHL cell lines (n=3, ±s )
3 讨 论
恶性淋巴瘤在全世界范围内的发病率及死亡率呈逐年增高趋势,占恶性肿瘤的4.1%,位居恶性肿瘤的第7位,主要包括霍奇金淋巴瘤和NHL。NHL由于组织学分类不同,其分子生物学行为、对治疗的反应、临床过程及预后等亦不同[1]。我们考虑肿瘤细胞的免疫逃逸机制在NHL不同的组织类型中起到了关键作用。
B7家族作为唯一能从抗原提呈细胞(antigen presenting cell,APC)单向传递信号至T细胞的共刺激分子,其成员有8种:B7-1(CD80),B7-2(CD86),B7-H1(PD-L1/CD274),B7-DC(PD-L2/CD273),ICOS-L(CD275),B7-H3(CD276),B7-H4和B7-H6[4,5]。PD-L1主要诱导性表达于激活的T细胞、B细胞、APC及人类肿瘤细胞上[5]。PD-L2和PD-L1在结构、基因序列及功能上相似,有40%的一致性,PD-L2不仅限制性表达于APC(如:巨噬细胞和树突状细胞)上,而且诱导性表达在T细胞、Th2细胞及肿瘤细胞上[6,7]。PD-Ls主要通过与其受体PD-1结合传递负性调控信号,诱导肿瘤抗原特异性T细胞的免疫无能或促进其凋亡,从而使肿瘤细胞逃避机体的免疫监视和杀伤。
研究证实,PD-Ls/PD-1途径在慢性感染、炎症及自身免疫性疾病等中发挥负性调控免疫应答的作用[7,8]。最近的研究[9−11]显示,在多种肿瘤微环境中(如:恶性黑色素瘤、肾癌、乳腺癌、肺癌、卵巢癌及血液系统恶性肿瘤等),PD-L1参与了肿瘤细胞的免疫逃逸。朱等[12]发现PD-1/PD-L1途径调节了肺癌恶性胸腔积液中CD8+T淋巴细胞IFN-γ的分泌,并促进了CD8+T淋巴细胞的凋亡。非小细胞肺癌中PD-L1与表皮生长因子受体(epithelial growth factor receptor,EGFR)的突变状态相关,与PD-L1阴性病例相比,PD-L1阳性病例对酪氨酸激酶抑制剂(tyrosine kinase inhibitor,TKI)治疗(如:吉非替尼、厄洛替尼)更敏感,其疾病进展时间明显延长[13]。这不仅说明PD-L1通过某种途径抑制了肺癌患者的抗肿瘤免疫,而且有助于筛选靶向治疗的优势人群,预示了抗PD-L1免疫治疗的潜在临床价值。在研究卵巢癌细胞毒性T淋巴细胞(cytotoxic lymphocyte,CTL)介导的肿瘤细胞溶解中发现[14],PD-L1高表达的肿瘤细胞其细胞溶解受到抑制,而将肿瘤细胞PD-L1基因沉默反而促进了其溶解效应。在腹膜转移的卵巢癌小鼠模型中,破坏了PD-L1基因后不仅抑制了肿瘤的生长,且延长了其生存期[14]。这充分说明肿瘤细胞通过高表达PD-L1而逃避了CTL的细胞毒作用。
但大多数研究仅关注了PD-L1所扮演的角色,而忽视了PD-L2的表达及意义。Iwamura等[15]发现PD-L2与T细胞受体结合后T细胞IFN-γ的分泌受到抑制,当沉默PD-L2 siRNA后其抑制效应解除,这高度提示PD-L2是通过提供负性调控信号来调节T细胞功能的。本研究同时检测了PD-L1及PD-L2在NHL细胞系中的表达,发现PD-L1及PD-L2在T-NHL、NKTL及B-NHL细胞株中均呈不同程度表达。以正常PBMCs为对照,T-NHL细胞系及NKTL细胞系中,PD-L1和PD-L2 mRNA表达量均明显高于正常PBMCs。而B-NHL细胞系中,PD-L1和PD-L2 mRNA表达量与正常PBMCs均无明显差异。我们的研究结果充分显示,B7家族高表达于NKTL及T-NHL细胞系上,这高度提示PD-Ls可能通过对机体免疫产生抑制效应参与其肿瘤细胞的免疫逃逸。而在B-NHL中,PD-L1及PD-L2的表达量与正常PBMCs无明显差异,可能是由于PD-Ls在其免疫逃逸机制中的贡献较小,或者由其他复杂的调节机制调控。
本研究表明B7家族高表达于NKTL及T-NHL细胞系,这高度提示其可能参与了这两种淋巴肿瘤的免疫逃逸。PD-L1及PD-L2将成为T-NHL及NKTL免疫治疗的新靶点。我们将进一步建立功能试验及动物模型来阐述B7家族在NHL中的免疫调节功能。
【参考文献】
[1]Sun Y, Shi YK.Manual of Medical Oncology[M].5th ed.Beijing:People’s Medical Publishing House, 2013, 355−360.[孙 燕, 石远凯.临床肿瘤内科手册[M].第5版.北京:人民卫生出版社, 2013, 355−360.]
[2]Yamamoto R, Nishikori M, Kitawaki T,et al.PD-1-PD-1 ligand interaction contributes to immunosuppressive microenvironment of Hodgkin’s lymphoma[J].Blood, 2008, 111(6):3220−3224.
[3]Iwamura K, Kato T, Miyahara Y,et al.siRNA-mediated silencing of PD-1 ligands enhances tumor-specific human T-cell effector functions[J].Gene Ther, 2012, 19(10):959−966.
[4]Greaves P, Gribben JG.The role of B7 family molecules in hematological malignancy[J].Blood, 2013, 121(5):734−744.
[5]Li Y, Wang J, Li C,et al.Contribution of PD-L1 to oncogenesis of lymphoma and its RNAi-based targeting therapy[J].Leuk Lymphoma, 2012, 53(10):2015−2023.
[6]Lesterhuis WJ, Steer H, Lake RA.PD-L2 is predominantly expressed by Th2 cells[J].Mol Immunol, 2011, 49(1−2):1−3.
[7]Rozali EN, Hato SV, Robinson BW,et al.Programmed death ligand 2 in cancer-induced immune suppression[J].Clin Dev Immunol, 2012, 2012:656340.
[8]Lin C, Chen X, Liu JN,et al.Advances of PD-1/PD-L1 signaling pathway in immune escape and treatment for non-small cell lung cancer[J].Chin J Lung Cancer, 2014,17(10):734−740.[林 城, 陈 雄, 刘静南.PD-1/PD-L1信号通路在非小细胞肺癌免疫逃逸及其治疗中的研究进展[J].中国肺癌杂志, 2014, 17(10):734−740.]
[9]Spranger S, Spaapen RM, Zha Y,et al.Up-regulation of PD-L1, IDO, and T(regs) in the melanoma tumor microenvironment is driven by CD8(+) T cells[J].Sci Transl Med, 2013, 5(200):200ra116.
[10]Soliman H, Khalil F, Antonia S.PD-L1 expression is increased in a subset of basal type breast cancer cells[J].PLoS One, 2014, 9(2):e88557.
[11]Liu Y, Carlsson R, Ambjorn M,et al.PD-L1 expression by neurons nearby tumors indicates better prognosis in glioblastoma patients[J].J Neurosci, 2013, 33(35):14231−14245.
[12]Zhu YX, Chen C, Huang JA,et al.The expression of PD-1 on CD8+T cells in malignant pleural effusion of lung cancer and its biological significance[J].Tumor, 2010, 30(9):778−781.[朱云霞, 陈 成, 黄建安, 等.PD-1分子在肺癌恶性胸腔积液CD8+T细胞上的表达及其生物学意义[J].肿瘤, 2010, 30(9):778−781.]
[13]Dlncecco A, Andreozzi M, Ludovini V,et al.PD-1 and PD-L1 expression in molecularly selected non-small-cell lung cancer patients[J].Br J Cancer, 2015, 112(1):95−102.
[14]Abiko A, Mandai M, Hamanishi J,et al.PD‑L1 on tumor cells is induced in ascites and promotes peritoneal dissemination of ovarian cancer through CTL dysfunction[J].Clin Cancer Res, 2013, 19(6):1363−1374.
[15]Iwamura K, Kato T, Miyahara Y,et al.siRNA-mediated silencing of PD-1 ligands enhances tumor-specific human T-cell effector functions[J].Gene Ther, 2012, 19(10):959−966.
