甘蓝型油菜WRKY基因家族碱基组成及密码子使用特性分析
2013-12-03李冠英张志燕方和娣谭小力
李冠英,王 政,张志燕,方和娣,谭小力
(江苏大学生命科学研究院,镇江212013)
密码子的非均衡使用在生物界中普遍存在[1]。通用遗传密码子共有64个,除去3个终止密码子外,其余61个密码子编码构成生物体的20种氨基酸,这表明同一种氨基酸由几个不同的密码子编码,通常将这些编码同一种氨基酸的不同密码子称为同义密码子(synonymous codon)。某一物种或某一基因家族对特定的同义密码子的选择使用具有偏好性,此现象称为密码子偏好性(codon bias),这一现象与遗传信息的载体分子DNA和生物功能分子蛋白质相关联,所以具有重要的生物学意义[2]。密码子偏好性是长期进化的结果,在进化过程中受很多因素的影响,以至于不同密码子承受的压力不同,从而产生密码子的偏好使用。影响密码子使用偏好性的因素有多种,如 tRNA 分子的丰度[3,4]、突变偏好性[5]、氨基酸疏水性[6]、基因表达水平[7]等。密码子的偏好使用还与基因的碱基组成相关,如G+C(GC)含量,尤其与GC3s(密码子第三位的 G+C)的含量相关[8]。另外,不同折叠类型蛋白的密码子使用偏好性有着显著的区别,这表明,在蛋白质二级结构和蛋白质折叠类型的预测过程中,除蛋白质一级结构之外,密码子偏好性是另一项重要的预测参考[9]。密码子偏好性的分析有助于在预测基因的表达水平、基因异源表达最适宿主的选择和优化密码子以提高异源表达水平等方面具有重要意义[2]。
WRKY基因家族是植物特有的一类转录因子,在拟南芥中的研究显示,该家族广泛地参与植物器官的发育和衰老、植物对生物、非生物胁迫的应答反应等一系列生理活动[10-12]。据报道显示,在甘蓝型油菜(Brassica napus)中,菌核病(Sclerotinia sclerotiorum)、黑斑病(Alternaria raphani)等油菜主要病原菌的侵染,以及各种植物激素的处理都能显著地诱导WRKY基因的表达[13-15],表明WRKY基因家族在油菜抗逆反应中起着重要的作用。
甘蓝型油菜(Brassica napus)是中国最重要的油料作物之一,其所产菜籽油在目前位居中国食用油市场第三位,占食用油消费量的18%[16]。本文主要从基因的碱基组成出发,分析甘蓝型油菜WRKY(BnWRKY)基因家族密码子的组成、使用特性、及其密码子偏好性,为更好实现该家族基因异源表达,进一步分析BnWRKY基因家族的性质奠定基础。
1 材料和方法
1.1 BnWRKY基因家族序列的获得
BnWRKY基因家族序列来源与Genbank数据库(http://www.ncbi.nlm.nih.gov/),踢除重复序列与不完整的CDS序列,最后得到44条完整CDS序列。
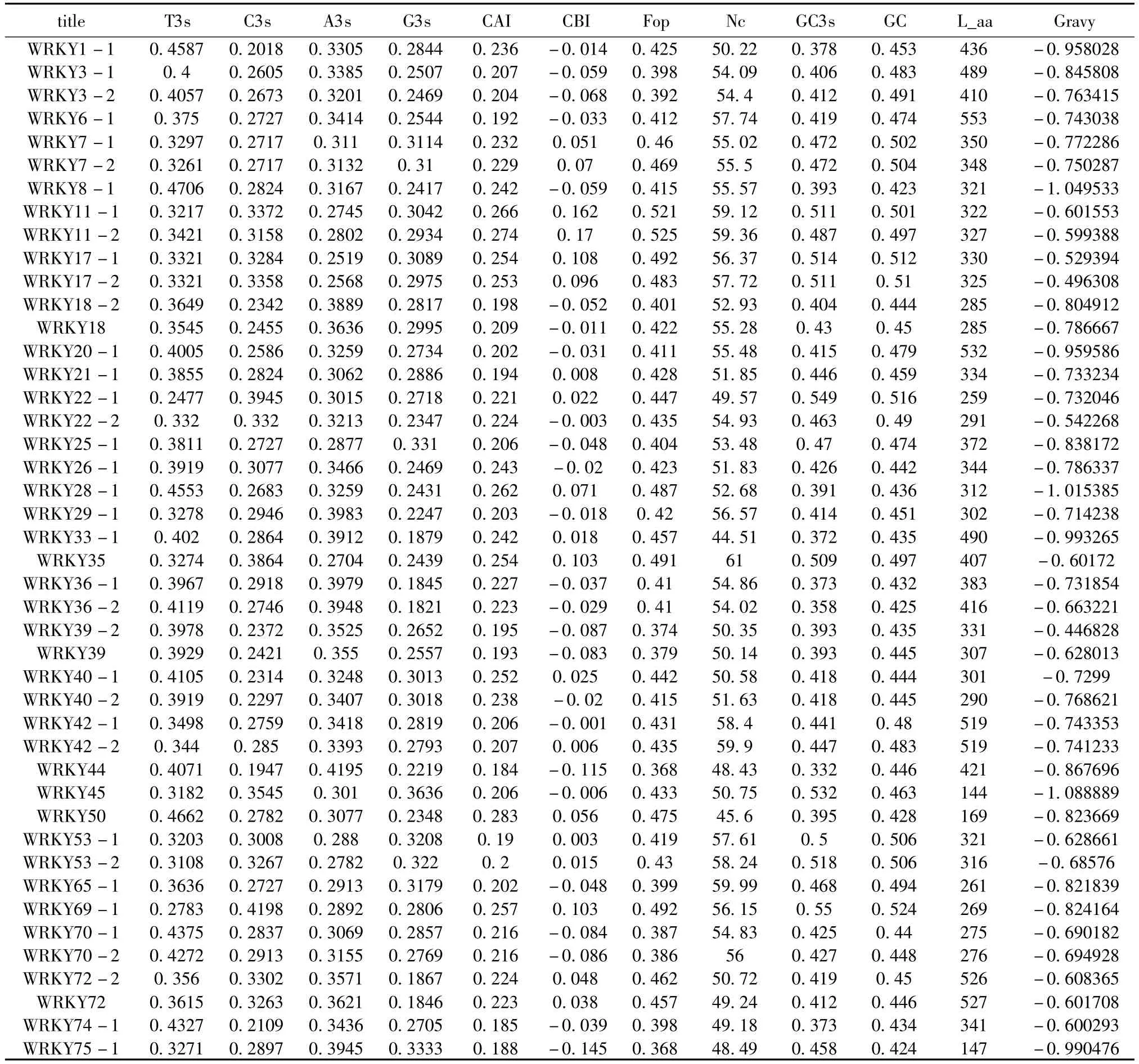
表1 BnWRKY基因家族密码子使用特性Table 1 Characterization of codon usage of rapseed WRKY genes
1.2 序列分析
利用CodonW软件分析WRKY家族基因密码子使用特性的参数包括:A3s,G3s,C3s,T3s(同义密码子在第3位上相应碱基的出现频率),GC(基因的 G+C含量),GC3s(密码子的第3位的G+C含量),ENC(有效密码子数),FOP(最优密码子使用频率),CBI(密码子偏爱指数),CAI(密码子适应指数),L_aa(氨基酸数),Gravy(平均亲水性值),RSCU(同义密码子的相对使用度)等。运用SPSS 19.0软件对BnWRKY家族的密码子特征各参数间相关性分析,运用Origin 8.0制作图表。
2 结果与分析
2.1 BnWRKY基因家族的密码子组成和使用性参数分析
表1所示,BnWRKY基因家族密码子各参数特征值。其中GC含量范围在42.3% ~52.4%,平均含量为46.6%,经分析发现其GC含量以平均值为界成双峰分布,主要集中在44.5%和50%左右(图1),总体来看,GC含量主要集中在50%左侧,说明BnWRKY基因家族略倾向于使用腺嘌呤(A)和胸腺嘧啶(T)这2种核苷酸。
从表1还可以看出,BnWRKY基因家族成员的Gravy值均为正值,表明该家族编码蛋白质的平均亲水性值均呈亲水性。
有效密码子数(Effective Number of Codon,ENC)是检测单个基因CDS区密码子偏好选择程度的一个参数,取值范围为20~61(单个基因每个氨基酸只选择使用一个密码子到每个密码子均被使用)[17,18]。Jiang 等[19]将ENc的值等于35时作为区分密码子选择偏好性强弱标准的界限,ENc值小于35时,其值越低,密码子的偏好性越强,ENc的值大于35时密码子没有偏好性。Bn-WRKY基因家族的ENC取值范围为44.51~61,集中在55左右(图2),均大于35,说明BnWRKY基因家族对密码子有一定的偏好性。BnWRKY基因家族的EENC取值范围为44.51~61,集中在55左右(图2),均大于35,说明BnWRKY基因家族对密码子有一定的偏好性。

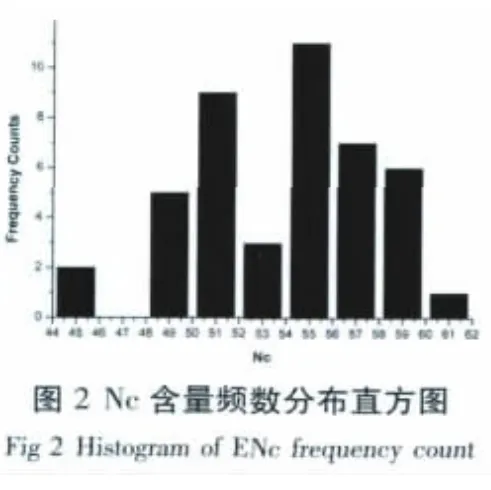
另外,通过比较ENC值可以预测内源基因表达量的相对高低,ENC值越大,该基因的表达量就越低,ENC值越小,该基因的内源表达量也越高[2]。BnWRKY基因家族,ENC值偏大,从而说明无胁迫条件下该家族基因表达水平一般。
CAI(密码子适应指数)是对单基因CDS区同义密码子与密码子最佳使用相吻合水平的检测,其值范围在0~1之间,CAI值一般也用来预测种内基因的表达水平[2]。BnWRKY 基因家族取值范围为0.184 ~0.283,进而说明该家族正常情况下表达水平不高。
2.2 BnWRKY基因家族密码子使用相关参数的相关性分析
为了研究多种可能因素对密码子偏好性有无影响及影响程度,我们作了各种参数的相关性分析(表2)。分析结果表明,GC含量与CBI、Fop、ENC均成极显著正相关(r>0.4,P <0.01),另外,GC3s、G3s、T3s、C3s分别与 CAI、CBI、Fop、ENC 等均有显著(P <0.05)或极显著(P<0.01)的相关性,显示出碱基组成对密码子的选择偏好性有重要影响;A3s与CAI、Fop、ENc等高表达基因相关的指数均成极显著负相关(r<-0.5,P<0.01),表明碱基在第三位上的腺嘌呤核苷酸(A3s)对基因的表达水平有显著的影响,并且还直接影响密码子的偏好性。

表2 各参数之间的相关系数Table 2 Correlation coefficients of the factors
2.3 BnWRKY基因家族 ENC-plot分析
由于碱基组成变异对密码子偏好性有重要影响,我们以ENC为纵坐标,以GC3s为横坐标,做ENc-plot分析(图3),检测碱基组成对密码子偏好性的影响[20]。图中的曲线表示在无进化压力的情况下ENC和GC3s的函数关系,如果密码子的偏好性仅受碱基突变的影响,代表基因的点应该落在理论曲线或其附近位置;在存在选择压力的情况下,相应的点会落在曲线下方较远处。从图3中可以看出,BnWRKY基因对应的点只有少数几个落在理论曲线上,暗示这些基因密码子的用法仅受碱基组成的影响,但大多数点落在曲线下方,则说明存在自然选择等其它压力对密码子偏好性也有重要影响。
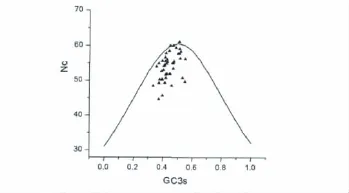
图3 BnWRKY基因的ENC-GC3s图Fig 3 ENc-GC3s plot of BnWRKY gene
2.4 BnWRKY基因家族密码子的同义密码子相对使用度
同义密码子相对使用度(Relative Synonymous Codon Usage,RSCU)是一个氨基酸所对应的密码子实际运用个数与理论运用个数之间的比值[22]。当实际运用个数与理论运用个数相同时,RSCU=1,此时密码子不存在偏好性;当RSCU>1时,则说明该密码子出现的频率比其它同义密码子高,为基因偏好使用的密码子;当RSCU<1时,则密码子出现的相对频率较低,是基因较少使用的密码子。在64个通用密码子中,TAA、TAG和TGA为终止密码子,不编码任何氨基酸,ATG、TGG分别是编码甲硫氨酸、色氨酸的唯一密码子,以上5个密码子均不存在偏好性,分析时排除这些密码子。
利用CodonW软件计算甘蓝型油菜WRKY基因家族的RSCU值(表3),RSCU>1的密码子如TTC、TTA、CTT、AGA等有29个,多以 T(10个)或 A(9个)结尾,说明BnWRKY基因家族偏爱使用以A或T结尾的密码子。
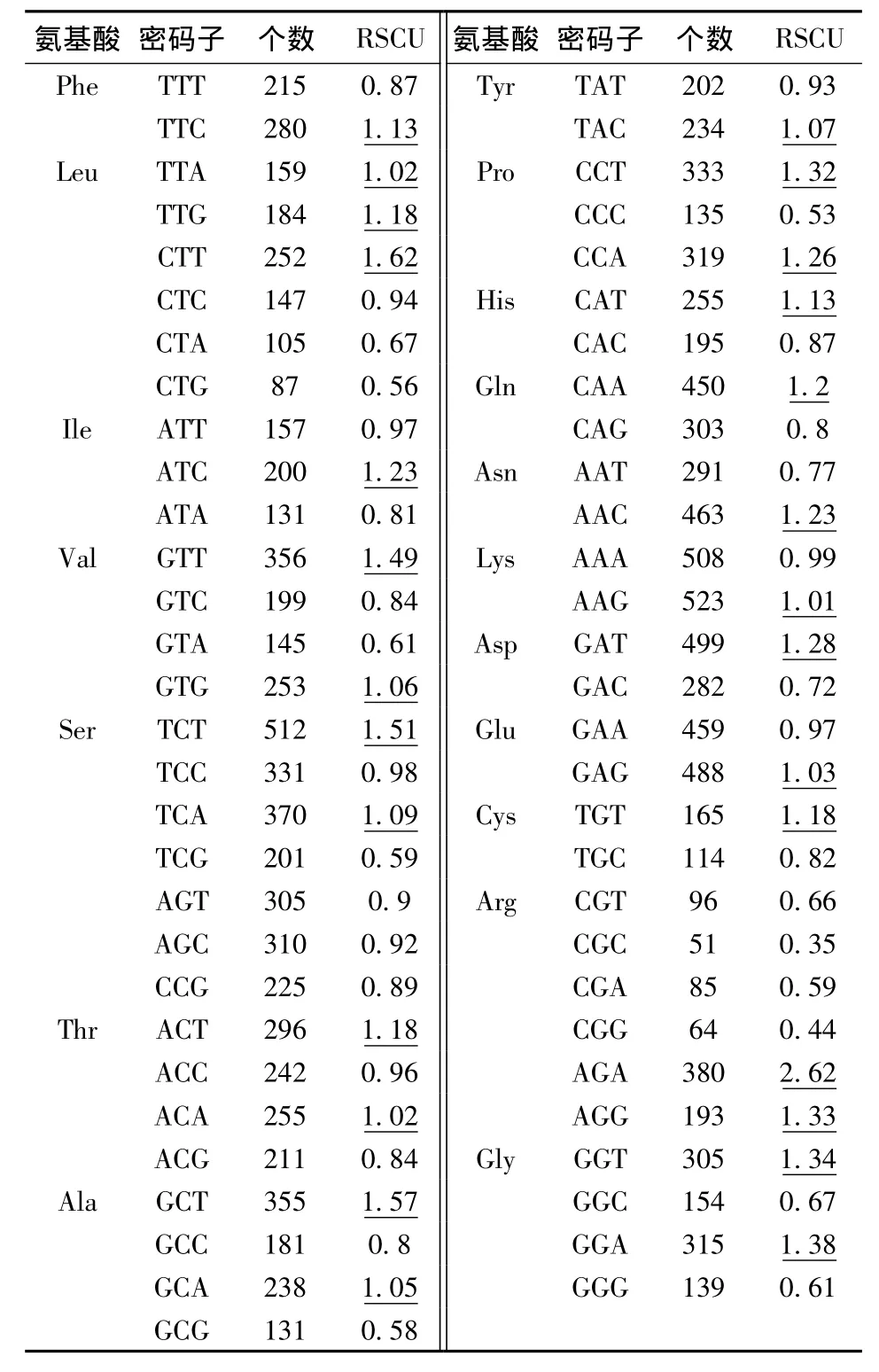
表3 甘蓝型油菜WRKY基因家族相对同义密码子使用度Table 3 Average relative frequence of 59 synonymous codons genes of WRKY families of rapseed
3 讨论
在对BnWRKY基因家族碱基组成的分析中发现,G+C含量总体均小于0.5,表明G+C的相对含量偏低(表1),即A+T的相对含量稍高;同时,在对密码子相对使用度(RSCU)的分析时发现,该家族偏爱的密码子多以A或T结尾(表3)。以上分析说明该基因家族的碱基组成对密码子偏爱的选择有一定的影响。这与我们做的相关性分析(G+C含量与多个参数呈显著或极显著相关,见表2)的结果相一致。ENC-plot曲线分析进一步发现,除了碱基组成外,自然选择等其它因素也对BnWRKY基因家族密码子的偏好性有影响,其影响程度有待进一步研究。
BnWRKY基因家族各成员的基因表达水平的相关参数(CAI、ENc和Fop等)的值均偏小(表1),根据这些数据我们推测,该基因家族在甘蓝型油菜中的表达量较低。在拟南芥的研究表明,AtWRKYs基因参与多种胁迫反应,在受到胁迫时进行诱导表达[22]。根据报道的实验数据显示,当甘蓝型油菜受到S.sclerotiorum、A.raphani等病原菌及各种植物激素处理时,测试的大部分Bn-WRKYs均被显著诱导,这些结果说明WRKY基因家族成员多为胁迫诱导型基因。
本研究调查了BnWRKY基因家族的密码子偏好性及其偏好程度,这些分析可为该基因家族新成员的鉴定及其表达分析预测提供线索;同时既可为该基因家族实现异源表达选择合适的宿主表达系统,也可为通过优化密码子提高基因的表达量提供依据。
[1]Fuglsang A.The'effective number of codons'revisited[J].Biochem Biophys Res Commun,2004,317(3):957-964.
[2]吴宪明,吴松锋,任大明,等.密码子偏性的分析方法及相关研究进展[J]. 遗传,2007,29(4):420-426.
[3]Moriyama E N,Powell J R.Codon usage bias and tRNA abundance in Drosophila[J].J Mol Evol,1997,45(5):514-523.
[4]Olejniczak M,Uhlenbeck O C.tRNA residues that have coevolved with their anticodon to ensure uniform and accurate codon recognition[J].Biochimie,2006,88(8):943-950.
[5]Xia X.Mutation and selection on the anticodon of tRNA genes in vertebrate mitochondrial genomes[J].Gene,2005,345(1):13-20.
[6]Knight R D,Freeland S J,Landweber L F.A simple model based on mutation and selection explains trends in codon and amino-acid usage and GC composition within and across genomes[J].Genome Biol,2001,2(4):H10.
[7]石秀凡,黄京飞,柳树群,等.人类基因同义密码子偏好的特征以及与基因GC含量的关系[J].生物化学与生物物理进展,2002,29(3):411-414.
[8]Carlini D B,Chen Y,Stephan W.The relationship between third-codon position nucleotide content,codon bias,mRNA secondary structure and gene expression in the drosophilid alcohol dehydrogenase genes Adh and Adhr[J].Genetics,2001,159(2):623-633.
[9]顾万君,马建民,周 童,等.不同折叠类型蛋白编码基因的密码子使用[J].东南大学学报(自然科学版),2002,32(3):362-366.
[10]Grunewald W,Karimi M,Wieczorek K,et al.A role for At-WRKY23 in feeding site establishment of plant-parasitic nematodes[J].Plant Physiol,2008,148(1):358-368.
[11]Jiang Y,Deyholos M K.Functional characterization of Arabidopsis NaCl-inducible WRKY25 and WRKY33 transcription factors in abiotic stresses[J].Plant Mol Biol,2009,69(1-2):91-105.
[12]De Michele R,Formentin E,Todesco M,et al.Transcriptome analysis of Medicago truncatula leaf senescence:similarities and differences in metabolic and transcriptional regulations as compared with Arabidopsis,nodule senescence and nitric oxide signalling[J].New Phytol,2009,181(3):563-575.
[13]Zhao J,Wang J,An L,et al.Analysis of gene expression profiles in response to Sclerotinia sclerotiorum in Brassica napus[J].Planta,2007,227(1):13-24.
[14]Yang B,Jiang Y,Rahman M H,et al.Identification and expression analysis of WRKY transcription factor genes in canola(Brassica napus L.)in response to fungal pathogens and hormone treatments[J].BMC Plant Biol,2009,9:68.
[15]Zhao J,Buchwaldt L,Rimmer S R,et al.Patterns of differential gene expression in Brassica napus cultivars infected with Sclerotinia sclerotiorum[J].Mol Plant Pathol,2009,10(5):635-649.
[16]沈金雄,傅廷栋.中国油菜生产、改良与食用油供给安全[J].中国农业科技导报,2011,13(1):1-8.
[17]Fuglsang A.The'effective number of codons'revisited[J].Biochem Biophys Res Commun,2004,317(3):957-964.
[18]Liu Q,Feng Y,Xue Q.Analysis of factors shaping codon usage in the mitochondrion genome of Oryza sativa[J].Mitochondrion,2004,4(4):313-320.
[19]Jiang Y,Deng F,Wang H,et al.An extensive analysis on the global codon usage pattern of baculoviruses[J].Arch Virol,2008,153(12):2273-2282.
[20]Wright F.The'effective number of codons'used in a gene[J].Gene,1990,87(1):23-29.
[21]Sharp P M,Li W H.An evolutionary perspective on synonymous codon usage in unicellular organisms[J].J Mol Evol,1986,24(1-2):28-38.
[22]Liu Z Q,Yan L,Wu Z,et al.Cooperation of three WRKY-domain transcription factors WRKY18,WRKY40,and WRKY60 in repressing two ABA-responsive genes ABI4 and ABI5 in Arabidopsis[J].J Exp Bot,2012,63(18):6371-6392.
