水螅基盘的固着行为及机理的初步研究
2013-12-03钱小成杨好强赵凤霞潘红春
钱小成,杨好强,赵凤霞,潘红春
(安徽师范大学生命科学学院重要生物资源保护与利用研究安徽省重点实验室生物环境与生态安全安徽省高校省级重点实验室,芜湖241000)
刺胞动物门水螅纲的淡水水螅是适合进行形态发生和个体发育调控机制研究的重要模式生物[1-3],因为水螅具有简单的单个极性体轴(口区-胃区-足区)[4,5]。极性体轴的一端为由口、垂唇和围绕口边缘着生的数根触手构成的口区,另一端为由柄和位于身体末端的基盘构成的足区,而介于口区和足区之间的区域即为胃区,柄和胃区的交界处为无性生殖的出芽区。在自然状态下,水螅的基盘固着于水中物体的表面、水螅体舒展伸长,其身体另一端即头区的触手自然延长下垂捕获水中快速运动的小型节肢动物溞类、与此同时触手上分布的大量刺细胞释放刺丝囊麻醉或杀死猎物,而后触手迅速收缩把猎物吞进胃循环腔。因此水螅基盘的固着行为是水螅进行正常捕食活动的前提条件。
长期以来,一直认为水螅是靠基盘粘液细胞分泌的粘液粘附于固着物表面[6-8],但有关水螅基盘固着行为及机理的相关信息非常有限,比如基盘吸附面与固着物表面之间吸附力的确切来源、粘液细胞在基盘吸附面上的分布情况等目前知之甚少。本研究组在实验室饲养水螅时发现,有时为了清洁培养水螅的玻璃烧杯、需要把水螅移到另外一个烧杯中,有些固着在烧杯侧壁上的水螅个体与烧杯壁表面间的粘附力很强、以至于用玻璃吸管多次冲水都不能让水螅基盘与烧杯壁分离,直至增大冲水力度导致水螅身体中间断裂时基盘还不能与烧杯壁分离,该现象对基盘与固着物之间粘附力仅来自粘液的观点提出了质疑。基盘粘液细胞是水螅身体上一种特化的终端分化细胞,除了能分泌粘多糖物质到细胞外形成粘液外,它还有一个重要特征是其细胞质内表达一种水螅基盘特异性过氧化物酶,该酶在粘液细胞中高量表达,把水螅活体放入含有ABTS[2,2'-azino-bis-(3-ethylbenzothiazoline-6-sulfonic acid)]和H2O2的反应体系中,基盘中表达过氧化物酶的粘液细胞能被染成紫红色[9]。基于此,本研究除了详细观察了水螅基盘的固着行为、还利用上述的ABTS组织化学方法对粘液细胞在基盘吸附面上的分布模式进行了显示,另外本文还对水螅基盘特异性过氧化物酶基因进行了分子克隆,期望探讨该酶高量表达与基盘固着行为之间是否存在内在联系。
1 材料和方法
1.1 实验材料
实验用水螅为本实验室培育的大乳头水螅(Hydra magnipapillata)中国品系单克隆无性繁殖系,原种采自安徽省芜湖市汀堂公园水塘(31°21'39.2″N,118°22'51.46″E)。水螅每天喂食一次卤虫无节幼体(Artemia sp.),喂食1 h后更换培养液,然后维持在温度22℃ ±0.5℃的光照培养箱中(光照强度=2000 LX,LD:HD=12 h :12 h)。
1.2 水螅基盘固着行为及形态学观察
借助本研究组大乳头水螅规模化培养平台,在喂食后换培养液时挑取一些基盘没有固着的水螅,置于凹玻片、再添加适量培养液,Olympus数码显微镜下进行连续观察和记录。
1.3 基盘固着时基盘吸附面的组织学观察
在水螅培养平台上挑选有水螅在侧壁上固着的水螅培养烧杯,先进行快速微波固定(微波炉频率2450 MHZ,功率800 W)1 min,然后Bouin氏固定液再固定15~20 min,梯度酒精脱水,水螅标本定位后石蜡包埋,做7 μm厚度的切片,苏木精及伊红双染,Olympus数码显微镜下观察和拍照。
1.4 基盘过氧化物酶表达的检测
通过ABTS细胞化学染色法检测水螅基盘粘液细胞分子标志物过氧化物酶的表达,观察水螅基盘粘液细胞在基盘吸附面上分布模式。将水螅置于白色陶瓷比色板上(每孔1只水螅),每孔分别加入350μL新鲜配制的ABTS染色混合液(655 mmol/L柠檬酸、345 mmol/L 柠檬酸盐和 1%ABTS 溶液各 700 μL,70 μL 0.3%H2O2,双蒸水定容至7 mL)[10],反应20 min 后用双蒸水洗涤2次,加入300 μL 2%NaF溶液,然后通过Olympus数码显微镜观察和拍照。
1.5 基盘过氧化物酶基因的分子克隆
1.5.1 大乳头水螅总RNA的提取及mRNA的纯化
取200条大乳头水螅,停止喂食3 d后用 Trizol试剂提取总RNA。具体操作按试剂盒说明书进行。总RNA提取后,溶于无RNA酶的水中,-70℃冰箱备用。取50 μL总 RNA并按 Oligotex mRNA Purification Kit说明书纯化mRNA。
1.5.2 SMART RACE cDNA文库的构建
实验操作基本按SMARTTM cDNA Library Construction Kit说明书进行。取3 μL mRNA样品约0.5 μg,加到0.2 mL的反应管中,按说明书加入相应试剂及引物,反转录合成第一链cDNA;采用LD-PCR(longdistance PCR)技术合成第二链cDNA。
1.5.3 大乳头水螅基盘特异性过氧化物酶全长cDNA序列的扩增及序列分析
根据水螅核基因组计划由DNA数据推导的水螅基盘特异性过氧化物酶cDNA序列(GenBank序列号:NW_002092004)[11]设计了一对引物(HMPPOD-F:5'-tattcatttgtttaatataac-3';HMPPOD-R:5'-tgattagaatataaacttat-cac-3'),引物由上海生工生物工程有限公司合成。以上述RACE cDNA文库为模板,扩增水螅基盘特异性过氧化物酶基因全长cDNA序列。PCR总反应体积为50 μL,其中含10 × buffer 5 μL,25 mmol MgCl23.0 μL,2.5 mmol dNTP 2.0 μL,引物(10 μmol)各 1 μL,Taq DNA聚合酶0.25 μL(Takara,5 U/μL),DNA 模板5 ~10 ng,使用灭菌后去离子水将反应总体积补至50 μL。反应体系95℃预变性5 min;95℃变性30 s,42℃复性30 s,68℃延伸90 s,循环次数为37;72℃延伸8 min。扩增产物经1.0%琼脂糖凝胶电泳,然后在凝胶成像系统上检测、成像。
PCR产物用柱式PCR产物纯化试剂盒经切胶纯化后用T/A Cloning Kit(TaKaRa)连接到克隆载体pMD 18-T vector中,再将重组质粒转化大肠杆菌E.coli JM109感受态细胞,挑取阳性克隆。单克隆菌株经扩大培养后,提取并纯化质粒,采用通用引物M13和M13 Reserve在LI-COR DNA测序仪上进行双向序列测定。
2 结果与分析
2.1 水螅基盘固着行为的动态观察
为描述方便,把水螅基盘固着过程分为3个时期。
2.1.1 固着准备期
水螅身体舒展伸长,基盘端逐渐靠近载玻片玻璃表面,最终紧贴载玻片(图1A-B)。
2.1.2 吸附面形成期
基盘与载玻片接触的吸附面面积逐渐扩大(图1C),吸附面的粘液细胞分泌大量粘液,由于基盘吸附面紧贴载玻片,所以较多的粘液从基盘吸附面与载玻片间被挤压出来(图1D)。
2.1.3 “吸垫”形成期
吸附面形成后约20 min在显微镜下可以清晰观察到仅仅吸附面的圆形周边区域与固着物表面直接接触、而吸附面中央区域不与固着物表面直接接触,即基盘吸附面形成类似“吸垫”的结构(图1E)。对固着时的基盘进行微波快速固定后的组织学研究结果直接证实了基盘吸附面中央区域与固着物表面不直接接触、而是朝胃循环腔方向形成凹陷(图1H)。另外,基盘吸附面形成类似“吸垫”结构后,吸附面的圆形周边区域的外胚层细胞沿着载玻片表面形成许多片状伪足和丝状伪足(图1F-G)。
2.2 水螅基盘粘液细胞分子标志物水螅基盘特异性过氧化物酶的表达模式
通过ABTS组织化学方法显示水螅基盘粘液细胞分子标志物基盘特异性过氧化物酶仅在基盘吸附面的周边区域外胚层细胞中表达(图2)、而吸附面中央区域无表达,这个表达模式表明粘液细胞在基盘吸附面上不是均匀分布的,而仅分布在吸附面的周边区域即吸附面与固着物直接接触的部位。
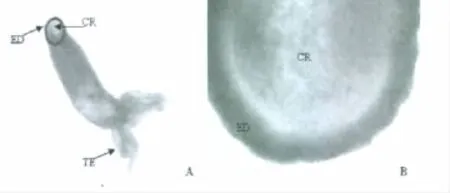
图2 大乳头水螅基盘特异性过氧化物酶的表达模式Fig 2 The expression pattern of foot-specific peroxidase in Hydra magnipapillata
2.3 水螅基盘特异性过氧化物酶基因的克隆和序列分析
水螅基盘特异性过氧化物酶基因扩增产物大小和预期片段大小接近(图3),PCR产物测序后得到的cDNA序列已提交GenBank(GenBank登录号:JF951860),该序列中蛋白质编码区共873 bp、编码290个氨基酸(图4)、其中第246核苷酸位点(C)与水螅核基因组计划由DNA数据推导的水螅基盘特异性过氧化物酶cDNA序列相应位点(T)的碱基不同。在NCBI网站进行在线BLAST分析后发现该酶蛋白氨基酸序列中隐含2个类似fascin蛋白的结构域(图5),它们分别位于基盘特异性过氧化物酶氨基酸序列的38~154位点及117~244位点。



3 讨论
从本研究的诸多结果综合起来看,水螅基盘固着时基盘吸附面与固着物间的吸附力可能有以下3个来源。首先,吸附面的圆形周边区域与固着物表面直接接触、而吸附面中央区域不与固着物表面直接接触(图1E),即基盘吸附面形成类似“吸垫”的结构应是基盘吸附力的主要来源。吸附面的圆形周边区域与固着物表面紧贴、从而起着封固作用,使得吸附面中央区域与固着物表面之间形成的空隙与外界水体隔绝,这个空隙区域相当于是一个负压区或低压区、内部可能没有水或有部分水。其次,基盘粘液细胞分泌的粘液在基盘吸附力的形成中演绎着重要角色,在基盘吸附面“吸垫”结构形成之前,粘液提供了基盘吸附面与固着物间初步的吸附力;在“吸垫”结构形成之后,吸附面的圆形周边区域与固着物表面间的粘液起着封固剂的作用、以保持吸附面中央区域与固着物表面之间空隙中的负压状态。第三,吸附面圆形周边区域外胚层细胞沿着载玻片表面形成的许多伪足增强了基盘吸附面的吸附力。
通过ABTS组织化学方法显示水螅基盘粘液细胞分子标志物基盘特异性过氧化物酶仅在基盘吸附面的周边区域外胚层细胞中表达(图2),而吸附面中央区域无表达,这个表达模式表明粘液细胞在基盘吸附面上不是均匀分布的,而仅分布在吸附面的周边区域即吸附面与固着物直接接触的部位,也说明基盘吸附面的周边区域外胚层细胞是基盘粘液细胞。对基盘特异性过氧化物酶氨基酸序列进行生物信息学分析的结果表明,基盘特异性过氧化物酶氨基酸序列中包含fascin蛋白的2个保守结构域。fascin蛋白是一种小分子球状蛋白,在细胞表面细胞膜突起的形成中起重要作用[12]。在一些小分子蛋白质因子如G-protein Rac等因素的激发下,细胞质中散乱的actin分子逐渐向细胞膜方向转移,同时细胞膜向外突起形成伪足,在fascin蛋白的作用下有序交联起来的actin分子聚合体就是伪足中的支撑物[13-15]。因此,水螅基盘特异性过氧化物酶可能具有的fascin蛋白活性有助于基盘粘液细胞伪足的形成、从而增强基盘对固着物的吸附力。
[1]Kasuminga O.Developmental neurobiology of hydra,a model animal of cnidarians[J].Canadian Journal Zoology,2002,80:1678-1689.
[2]Steele R E.Developmental signaling in hydra:what does it take to build a"simple"animal[J].Dev Biol,2002,248:199-219.
[3]Bosch T C.Ancient signals:peptides and the interpretation of positional information in ancestral metazoans[J].Comp Biochem Physiol Part B Biochem Mol Biol,2003,136:185-196.
[4]Zeretzke S,Perez F,Velden K,et al.Ca2+-ions and pattern control in Hydra[J].Int J Dev Biol,2002,46:705-710.
[5]Kobayakawa Y,Kodama R.Foot formation in hydra:commitment of the basal disk cells in the lower peduncle[J].Dev Growth Differ,2002,44:517-526.
[6]Kepner W A,Thomas W L.Histological features correlated with gas secretion in Hydra oligactis Pallas[J].Biological Bulletin,1928,54(6):529-533.
[7]Kobayakawa Y,Kodama R.Foot formation in Hydra:commitment of the basal disk cells in the lower peduncle[J].Development,Growth& Differentiation,2002,44(6):517 526.
[8]Galliot B,Chera S.The Hydra model:disclosing an apoptosis driven generator of Wnt-based regeneration [J].Trends in Cell Biology,2010,20:514-523.
[9]Ghaskadbi S S,Shetye L,Chiplonkar S,et al.Ultraviolet irradiation initiates ectopic foot formation in regenerating hydra and promotes budding[J].Biosci,2005,30(2):177-182.
[10]徐金金,潘红春,金 灿,等.大乳头水螅基盘再生进程中过氧化物酶的表达[J].中国组织化学与细胞化学杂志,2010,19(3):269-273.
[11]Chapman J A,Kirkness E F,Simakov O,et al.The dynamic genome of hydra[J].Nature,464:592-596.
[12]Adams J C.Roles of fascin in cell adhesion and motility[J].Curr Opin Cell Biol,2004,16:590-596.
[13]Schafer C,Borm B,Born S,et al.One step ahead:role of filopodia in adhesion formation during cell migration of keratinocytes[J].Exp Cell Res,2009,315:1212-1224.
[14]Svitkina T M,Bulanova E A,Chaga O Y,et al.Mechanism of filopodia initiation by reorganization of a dendritic network[J].J Cell Biol,2003,160:409-421.
[15]Khurana S,George S P.The role of actin bundling proteins in the assembly of filopodia in epithelial cells[J].Cell Adh Migr,2011,5:409-420.
