常压室温等离子体诱变高效利用木糖产丁二酸菌株
2013-11-12万青曹伟佳张常青刘嵘明梁丽亚陈可泉马江锋姜岷
万青,曹伟佳,张常青,刘嵘明,梁丽亚,陈可泉,马江锋,2,姜岷
1 南京工业大学 生物与制药工程学院 材料化学工程与国家重点实验室,江苏 南京 211816
2 中国石化扬子石油化工有限公司南京研究院,江苏 南京 210048
大肠杆菌Escherichia coli AFP111是NZN111(△pflAB△ldhA)的ptsG自发突变株,其恢复了在厌氧条件下代谢葡萄糖的能力[1-2]。在AFP111中,转化1 mol木糖合成丁二酸的过程中净产生1.67 mol的ATP,但是转化1 mol的木糖合成丁二酸的过程中实际需要2.67 mol的ATP[3-4]。因此,ATP供给不足导致AFP111不能代谢木糖并合成丁二酸[5]。本课题组为了获得可以在厌氧条件下代谢木糖的菌株,尝试用诱变的方法来选育目的菌株。
常温常压等离子体诱变系统 (ARTP)由清华大学研发,通过 ARTP所产生的射线和活性粒子束对菌株的遗传物质造成损伤,从而引起菌株的突变[6-7]。ARTP具有射流温度低、产生的等离子体均匀、无需真空装置、操作简易、成本低、与生物大分子和细胞作用明显等优点[8],已成为快速突变微生物基因组的有效方法。本研究通过利用ARTP常温常压等离子体诱变系统对出发菌株AFP111进行诱变,通过以木糖为碳源的固体平板筛选,最终得到一株在厌氧条件下可以代谢木糖并积累丁二酸的突变株。
1 材料与方法
1.1 材料
Escherichia coli strain AFP111 [F+ λrpoS396(Am)rph-1 △(pflAB::Cam)△(ldhA::Kan)△ptsG],由David P. Clark 教授 (Southern Illinois University)惠赠。
1.2 方法
1.2.1 ARTP诱变
利用常压室温等离子体育种机 (ARTP)进行诱变[9]。以氦气为气体,ARTP射频等离子体的气体流量为 10 L/min;作用距离 2 mm;作用功率120 W。将培养好的种子用0.85%的生理盐水稀释至OD600=1,取10 μL菌液均匀涂布于无菌金属平板上,利用无菌风将菌液吹干;ARTP处理适当的时间,将诱变后的菌体置于装有1 mL无菌生理盐水的离心管中;将菌液稀释 1×104涂布于固体平板上,37 ℃厌氧培养。
1.2.2 摇瓶培养
用接种环从平板上挑取单菌落,接种到装液量为5 mL LB的血清瓶中,加入2~3 g/L木糖,通入无菌过滤的CO2气体2 min,37 ℃、200 r/min培养12 h作为一级种子。将一级种子按10%的接种量接种到装液量为30 mL M9培养基的血清瓶中,加入20 g/L木糖,通入无菌过滤的CO2气体2 min,37 ℃、200 r/min培养72 h。
1.2.3 分析方法和酶活分析
细胞生长用紫外可见分光光度计于波长600 nm处测定吸光度值。有机酸、木糖用高效液相色谱法 (HPLC)检测[10]。磷酸烯醇式丙酮酸羧激酶(PCK)和磷酸烯醇式丙酮酸羧化酶 (PPC)酶活的测定方法见参考文献[11]。
2 结果与分析
2.1 ARTP诱变致死率曲线
由图1可以看出,等离子体对AFP111的杀伤力比较强,ARTP处理6 s会杀死80%的菌体;处理10 s以后致死率达到90%以上;处理30 s致死率达到100%。据文献报道,当存活率为1%~10%时,外界处理对细胞的诱变效应较强[12-13],本实验选取15 s对细胞进行处理。
2.2 厌氧菌株的选育
以平板上菌落的大小作为初选条件,并辅以HPLC精确检测突变菌株产酸性能,最后筛选到4株性能良好的菌株,将此 4株菌进行厌氧摇瓶发酵验证,其发酵结果见表1。
由表 1可以看出,在厌氧条件下,出发菌株根本无法代谢木糖,而突变株可以在厌氧条件代谢木糖并积累丁二酸,其中突变株DC111生长及产酸的性能最好,发酵 72 h菌体 OD600达到了2.26,丁二酸产量为 6.46 g/L,丁二酸得率为0.78 mol/mol,并且其在厌氧条件下的生长和耗糖曲线,如图2所示。
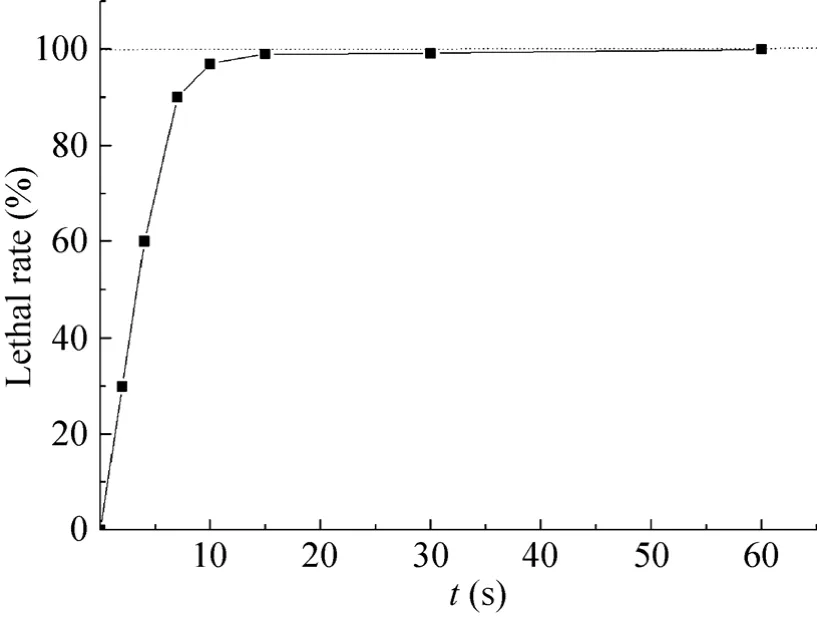
图1 大肠杆菌AFP111的致死率曲线Fig. 1 Variaton of the lethal rate of AFP111 with the ARTP treatment time.
2.3 PCK和PPC酶活测定
在 AFP111厌氧发酵途径中, 磷酸烯醇式丙酮酸羧化酶 (PPC)和磷酸烯醇式丙酮酸羧化激酶 (PCK)皆可催化由磷酸烯醇式丙酮酸 (PEP)到草酰乙酸 (OAA)的反应[14]。而由PCK催化的反应伴有ATP的生成[15-16],对突变株和出发菌株的PPC和PCK比酶活测定,结果如表2所示。
由表2可以看出,突变株DC111中的PCK的比酶活提高了19.33倍,而PPC的比酶活降低了2.32倍。因此,在 DC111中,PEP转化为 OAA的反应PCK起到了主导作用,而在PCK催化过程中伴有1 molATP的生成[7],从而使DC111在厌氧条件下能够有足够的ATP供给来代谢木糖并积累丁二酸。

表1 AFP111和突变株纯厌氧发酵性能比较Table 1 Comparation between AFP111 and the mutants

图2 DC111的生长和耗糖曲线Fig. 2 Anaerobic growth and xylose consumption of DC111.

表2 DC111和AFP111的比酶活Table 2 Specific activities of PPC and PCK in crude extractsof the strain DC111 and AFP111
[1]Chatterjee R, Millard CS, Champion K, et al. Mutation of the ptsG gene results in increased production ofsuccinate in fermentation of glucose by Escherichia coli. Appl Environ Microbiol, 2001, 67: 148–154.
[2]Donnelly MI, Millard CS, Clark DP, et al. A novel fermentation pathway in an Escherichia coli mutant producing succinic acid, acetic acid, and ethanol. Appl Biochem Biotechnol, 1988, 70(72): 187–198.
[3]Hasona A, Kim Y, Healy FG, et al. Pyruvate formate lyase and acetate kinase are essential for anaerobicgrowth of Escherichia coli on xylose. J Bacteriol, 2004, 186: 7593–7600.
[4]Andersson C, Hodge D, Berglund KA, et al. Effect of differentcarbon sources on the production of succinic acid using metabolicallyengineered Escherichia coli. Biotechnol Prog, 2007, 23: 381–388.
[5]Gonzalez R, Tao H, Shanmugam KT, et al. Global gene expression differences associated with changes inglycolytic flux and growth rate in Escherichia coli during thefermentation of glucose and xylose. Biotechnol Prog, 2002, 18: 6–20.
[6]Lu Y, Wang LY, Ma K, et al. Characteristics of hydrogen production of an Enterobacter aerogenes mutantgenerated by a new atmospheric and room temperature plasma(ARTP). Biochem Engin J, 2011, 55: 17–22.
[7]Li G, Li HP, Wang YL, et al. Genetic effects of radio-frequency, atmospheric-pressureglow discharges with helium. Appl Phys Lett, 2008, 92: 221504.
[8]Li HP, Li G, Sun WT, et al. Radio-frequency,atmospheirc-pressureglow discharges: producingmethods,characteristics and applications in bio-medicalfields.Complex Systems, 2008, 982: 584-591.
[9]Guo T, Tang Y, Xi YL, et al. Clostridium beijerinckii mutant obtained by atmosphericpressure glow discharge producing high proportionsof butanol and solvent yields.Biotechnol Lett, 2011, 33: 2379–2383.
[10]Liu RM, Liang LY, Chen KQ, et al. Fermentation of xylose to succinate by enhancement of ATPsupply in metabolically engineered Escherichia coli. Appl Microbiol Biotechnol,2012, 94: 959–968.
[11]Wu H, Min LZ, Zhou L, et al. Improved succinic acidproduction in the anaerobic culture of an Escherichia coli pflB ldhA double mutant as a result of enhanced anapleroticactivities in the preceding aerobic culture. Appl Environ Microbiol, 2007, 73: 7837–7843.
[12]Hua XF, Wang J, Wu ZJ, et al. A salt tolerant Enterobacter cloacae mutant for bioaugmentation of petroleum and salt-contaminated soil. Biochem Eng J, 2010, 49: 201–206.
[13]Laroussi M. Low temperature plasma-based sterilization:overview and state-of-the-art. Plasma Proc Polym, 2005, 2:391–400.
[14]Millard CS, Chao YP, Liao JC, et al. Enhanced productionof succinic acid by overexpression of phosphoenolpyruvate carboxylase in Escherichia coli. Appl Environ Microbiol, 1996, 62: 1808–1810.
[15]Kim P, Laivenieks M, Vieille C, et al. Effect of overexpression of Actinobacillus succinogenes phosphoenolpyruvate carboxykinase on succinate production in Escherichia coli. Appl Environ Microbiol,2004, 70: 1238–1241.
[16]Laivenieks M, Vieille C, Zeikus JG, et al. Cloning,sequencing and overexpression of the Anaerobiospirillum succiniciproducens phosphoenolpyruvate carboxykinase(pckA)gene. Appl Environ Microbiol, 1997, 63:2273–2280.
