带Myc、His标签的SPAG4L真核表达载体的构建与表达
2013-11-12陈颜亮郑治王剑龙周晓哲李艳杨梦黄利华邢晓为
陈颜亮,郑治,王剑龙,周晓哲,李艳,杨梦,黄利华,邢晓为
1 中南大学湘雅三医院骨科,湖南 长沙 410013
2 中南大学湘雅三医院医学实验中心,湖南 长沙 410013
精子发生障碍是引起男性不育的主要原因之一,在精子发生过程中,生殖细胞先后经历精原细胞有丝分裂阶段、精母细胞的减数分裂阶段和精子细胞的变态成形阶段,最后发育为成熟的精子[1-3]。在精子发生过程中的每个阶段都会有一些特异性的基因发挥特定作用,维持精子发生这一重要生殖生理活动[3]。
当前国内外的研究认为,Y染色体上的AZF基因微缺失与男性不育有关[4–5]。除了AZF基因外,还有许多常染色体上的基因也参与精子发生过程[6–7]。近年来研究发现,一个新的包含SUN结构域的蛋白家族参与精子发生过程。该蛋白家族最为明显的特征是 C端有一个保守的SUN结构域,靠近N端有跨膜区(TM),TM和SUN结构域之间有coiled-coil区[8-10]。目前已经发现在哺乳动物中至少存在5种以上SUN结构域蛋白,已报道的有 SUN1、SUN2、SUN3、SPAG4 (又称 SUN4)和 SPAG4L(又称 SUN5 或TSARG4)等[10]。
在前期研究中,我们从人类精子尾部基因SPAG4出发,克隆了一个与该基因同处染色体20q11.21区域的人类睾丸新基因 SPAG4L(或SUN5) (GenBank Accession No. AF401350)[11-13]。通过对不同发育阶段的小鼠研究发现,Spag4L(或称 SRG4)基因是一个小鼠睾丸特异性表达基因,其表达严格受生长发育调控[14]。近年来研究发现,小鼠Spag4L蛋白主要在精母细胞、圆形精子细胞及成熟精子细胞中表达,在精原细胞中不表达[13-15]。通过对 SPAG4L及其突变体的亚细胞定位发现,SPAG4L在细胞的内质网和核膜表达,跨膜区 (TM)和 coiled-coil区对于 SPAG4L在核膜上的定位是必需的[13]。Frohnert等[15]研究发现,Spag4L在小鼠睾丸中有两个转录本,该蛋白在精子中介导核膜和顶体的连接。我们研究发现,Spag4L蛋白在分离的小鼠精母细胞中能够与内质网标志物 PDI和内核膜标志物Lamin B1共定位。通过对减数分裂中Spag4L的表达变化进行观察,发现Spag4L 在减数分裂过程中参与胞核的迁移和核膜重塑[13]。
为了获得 SPAG4L蛋白,前期研究中我们成功地用PQE-30对SPAG4L的C末端进行了原核表达[16],该蛋白可用于研究SPAG4L的C端SUN结构域区的功能,但是不能用于研究N端SPAG4L的功能。SPAG4L是通过N端与核膜上的分子相互作用,从而滞留在核膜上。我们试用了几种原核表达载体对SPAG4L N端进行表达,均未成功。为了获得带有His、Myc 双标签的 SPAG4L全长蛋白,本研究从人睾丸cDNA中扩增 SPAG4L开放阅读框,构建到pcDNA3.1/myc-His(-)A 真核表达载体的 CMV启动子下游,获得 pcDNA3.1/myc-His(-)A/SPAG4L重组质粒。随后,将该重组质粒导入到HeLa细胞,经G418筛选后,挑取单克隆建立稳定转染 SPAG4L的细胞系,该细胞系能够表达带有 Myc 和 6×His标签的 SPAG4L蛋白。通过本研究,建立稳定表达 SPAG4L全长蛋白的细胞系,将为下一步进行免疫共沉淀和pull-down实验提供了理想的蛋白,为深入研究SPAG4L蛋白定位在核膜上的分子机制奠定了基础。
1 材料与方法
1.1 材料
人睾丸cDNA、感受态细菌JMl09及HeLa细胞由本实验中心保存;Easy-A高保真PCR扩增酶购自 Agilent公司;SPAG4L抗体购自ABGAB公司(用于免疫荧光)和 Proteintech 公司(用于Western blotting);PDI 抗体购自Santa Cruz公司;G418购自Amresco公司;限制性内切酶购自 Fermentas公司;pcDNA3.1/myc-His(-)A载体购自Invitrogen 公司;His 单克隆抗体、Myc 单克隆抗体、GAPDH 单克隆抗体、FITC标记山羊抗兔IgG(H+L)、Cy3标记山羊抗小鼠IgG(H+L)均购自碧云天生物技术研究所;高糖DMEM培养基、LipofectamineTM2000、胎牛血清购自Gibco公司等。
1.2 方法
1.2.1 PCR扩增SPAG4L基因开放阅读框
根据我们提交国际GenBank登录的序列,用 PCRDESN软件设计引物,在上游引物中引入酶切位点NheⅠ,在下游引物中引入酶切位点KpnⅠ,并分别在上游和下游引物的5'端加上保护碱基。10 µL PCR扩增体系如下:10×Easy-A反应缓冲液 1 µL,2.5 mmol/L dNTPs 1 µL,上、下游引物各0.1 µL,Easy-A高保真PCR扩增酶0.2 µL,睾丸cDNA模板0.8 µL,加双蒸水补充至10 µL。在PE9700型PCR仪上进行PCR扩增,PCR反应条件为:95 ℃预变性2 min 30 s;94 ℃变性30 s,退火30 s,72 ℃延伸1 min,扩增35个循环;72 ℃延伸10 min,最后降至4 ℃保温。扩增产物经15 g/L琼脂糖电泳进行检测。
1.2.2 pcDNA3.1/myc-His(-)A/SPAG4L重组质粒的构建
按照参考文献[10]所述方法将 PCR扩增的SPAG4L基因片段插入到pUCm-T载体,转化感受态JM109后,在含有氨苄青霉素的LB固体培养基上培养。所筛选阳性克隆经PCR扩增鉴定后抽提质粒进行双酶切,回收目的片段,亚克隆到pcDNA3.1/myc-His(-)A表达载体中。所筛选的克隆经PCR鉴定后,抽提质粒进行酶切鉴定和测序。
1.2.3 稳定转染 pcDNA3.1/myc-His(-)A/SPAG4L细胞系的建立
将消化好的HeLa细胞接种6孔板,每孔接种约1×105个细胞,待细胞融合至80% 左右,按照 LipofectamineTM2000操作说明书提供的方法将 pcDNA3.1/myc-His(-)A 空质粒和pcDNA3.1/myc-His(-)A/SPAG4L分别转染HeLa细胞,24 h后,更换含有G418(800 mg/L)的选择性培养基进行筛选,每 3天换液一次。筛选35~40 d后,获得G418抗性细胞克隆。检测阳性细胞克隆,用有限稀释法进行亚克隆,用含G418的培养基进行培养,观察克隆形成情况,待克隆形成后挑单克隆进行扩大培养,检测SPAG4L的表达,最终获得单克隆阳性表达SPAG4L的转染细胞系。
1.2.4 Western blotting检测稳定转染细胞系中SPAG4L蛋白的表达
按照RIPA试剂盒说明书所述方法分别提取稳定转染 pcDNA3.1/myc-His(-)A空质粒和pcDNA3.1/myc-His(-)A/SPAG4L的细胞总蛋白,测定浓度,经过SDS-PAGE电泳后转PVDF膜。Western blotting检测按照常规方法进行。
1.2.5 免疫荧光技术检测SPAG4L表达
在 6孔板底预先放置灭菌好的洁净盖玻片,接种细胞,放置到在37 ℃、50 mL/L CO2培养箱中培养 24 h,取出后用D-Hanks 液洗脱3次,经4 %多聚甲醛固定。参照我们已建立的方法进行免疫荧光实验,检测SPAG4L蛋白、标签Myc,标签His以及内质网标志蛋白PDI的表达[13]。
2 结果
2.1 PCR扩增SPAG4L基因结果
PCR扩增 SPAG4L基因开放阅读框,扩增产物经1.5%琼脂糖凝胶电泳检测,发现SPAG4L基因扩增条带大小符合实验设计。
2.2 pcDNA3.1/myc-His(-)A/SPAG4L重组质粒的鉴定
挑取转化了 pUCm-T-SPAG4L重组质粒的阳性克隆,在含氨苄青霉素的液体LB培养基中培养至合适的浓度,以菌液为模板,PCR扩增SPAG4L基因,筛选阳性克隆。分别将6号和2号阳性克隆抽提质粒DNA后测序,将测序正确的6号质粒用NheⅠ和KpnⅠ双酶切,回收目的基因片段,插入到同样经过 NheⅠ和 KpnⅠ双酶切的pcDNA3.1/myc-His(-)A表达质粒中,筛选阳性克隆,用上述SPAG4L基因引物进行PCR检测,发现所挑选的阳性克隆能够扩增出SPAG4L基因(图 1),随后,抽提质粒,用限制性内切酶NheⅠ和KpnⅠ进行双酶切分析,结果发现,重组质粒中所插入的SPAG4L基因片段大小正确(图 2)。对 pcDNA3.1/myc-His(-)A/SPAG4L进行双向测序,结果表明,所插入的SPAG4L基因序列没有发生点突变和移码突变,说明重组质粒 pcDNA3.1/myc-His(-)A/SPAG4L构建成功。
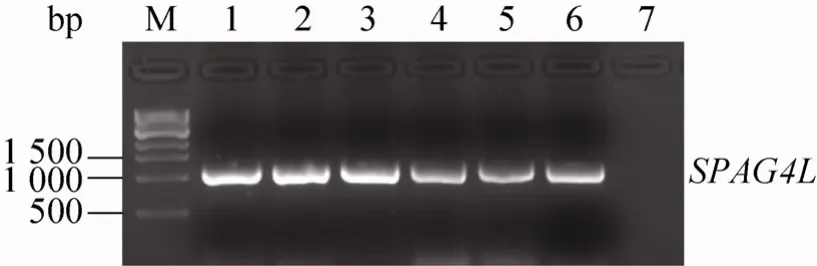
图1 PCR鉴定pcDNA3.1/myc-His(-)A/SPAG4L阳性克隆Fig. 1 Positive clones of pcDNA3.1/myc-His(-)A/SPAG4L were identified by PCR. M: 1 kb ladder; 1–5:positive clones; 6: positive control; 7: H2O.
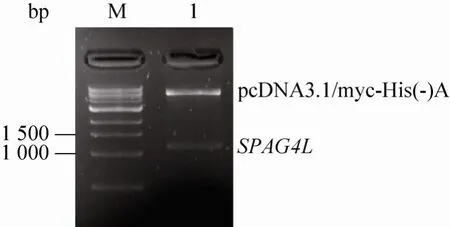
图2 酶切鉴定pcDNA3.1/myc-His(-)A/SPAG4L重组质粒Fig. 2 Identification of recombinant plasmid pcDNA3.1/myc-His(-)A/SPAG4L by enzyme digestion.M: 1 kb ladder; 1: pcDNA3.1/myc-His(-)A/SPAG4L plasmid.
2.3 Western blotting检测稳定转染细胞系中SPAG4L蛋白表达
我们将筛选得到的亚克隆进行扩大培养,提取蛋白,应用Western blotting技术检测稳定转染细胞系中 SPAG4L及其标签蛋白表达。结果发现,在稳定转染 pcDNA3.1/myc-His(-)A/SPAG4L的细胞系中,带有Myc和His标签的SPAG4L蛋白能够正确表达,蛋白大小在43 kDa左右,蛋白大小与预期相符。而稳定转染了空白质粒pcDNA3.1/myc-His(-)A的细胞系中,用SPAG4L、Myc、His抗体,均未在43 kDa左右处检测到目的条带,而内参GAPDH蛋白在转染pcDNA3.1/myc-His(-)A/SPAG4L组和空白质粒组均有表达(图3)。
2.4 免疫荧光检测结果
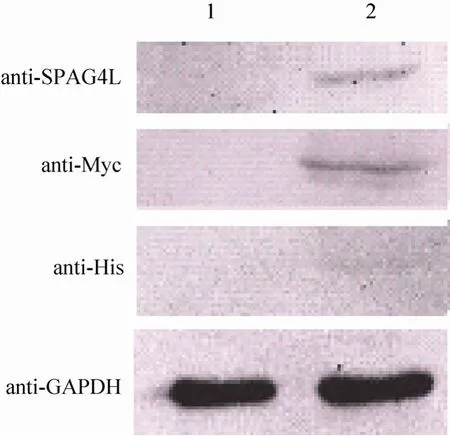
图3 Western blotting检测新建细胞系中SPAG4L及其标签蛋白的表达Fig. 3 Expression of SPAG4L and its tags was detected in stable transfected cell lines. The protein samples were obtained from HeLa cell lines stably transfected with pcDNA3.1/myc-His(-)A (Lane 1)and pcDNA3.1/myc-His(-)A/SPAG4L (Lane 2).
为了观察 SPAG4L稳定转染细胞系中SPAG4L及其标签蛋白是否表达,应用兔抗人SPAG4L抗体和Myc单克隆(小鼠来源)抗体对稳定转染SPAG4L的细胞系进行免疫荧光检测。结果发现,用兔抗人 SPAG4L一抗杂交后,用FITC标记山羊抗兔IgG(H+L)进行检测,可见细胞核膜及其周围出现绿色荧光信号;用Myc单克隆抗体作为一抗进行免疫荧光实验,用 Cy3标记山羊抗小鼠IgG(H+L)进行检测,可见细胞核膜及其周围出现红色荧光信号(图4)。由此可见,SPAG4L及其标签蛋白在稳定转染细胞系中能够正确表达,即主要分布在细胞核膜及其周围。
PDI是一种主要存在于内质网腔,大小为55 kDa的多功能蛋白,其功能是在内质网腔中催化二硫键的形成,促使新生肽链成熟成有生物活性的蛋白质。本研究中,我们将 PDI作为内质网的标志蛋白与 SPAG4L在稳定转染SPAG4L的细胞系中进行免疫共定位实验,结果发现,标记为绿色的 SPAG4L蛋白能够和标记为红色的PDI蛋白共定位,说明SPAG4L除了在核膜表达,而且还在内质网表达,该结果与前期本课题组研究的 SPAG4L蛋白亚细胞定位结果一致(图 5)。PDI是一个多功能蛋白,SPAG4L是否与PDI相互作用,还有待于进一步研究。
3 讨论
精子发生是一个复杂的过程[1-2]。前期的研究发现,SUN 蛋白(SUN1、SUN2、SUN3、SPAG4和 SPAG4L)是一类新发现的核膜蛋白,都参与了精子的发生[9,11,13,17-18]。
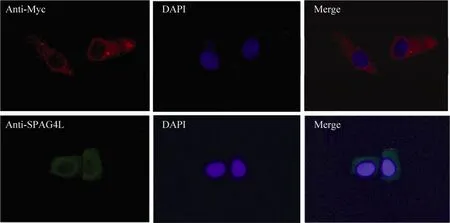
图4 免疫荧光技术检测稳定转染pcDNA3.1/myc-His(-)A/SPAG4L细胞中SPAG4L和myc蛋白表达Fig. 4 Expression of SPAG4L and myc-tag was detected in stable transfected cell lines by immunofluorescence(200×).
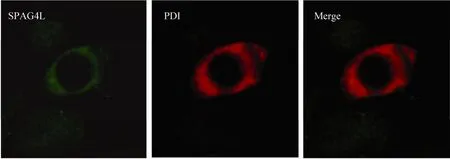
图5 稳定转染pcDNA3.1/myc-His(-)A/SPAG4L的细胞中SPAG4L与内质网蛋白PDI免疫共定位Fig. 5 SPAG4L colocalized with ER marker PDI by immunofluorescence in cells stably transfected with pcDNA3.1/myc-His(-)A/SPAG4L (400×).
SUN1和SUN2在精子发生的整个过程中均有表达,将小鼠中Sun1基因敲除,将会导致小鼠出现雄性不育[17]。SUN3是一个睾丸特异性表达蛋白,能够与Nesprin-1相互作用形成LINC复合物,将分化中的精子核与周围的细胞骨架结构连接在一起,从而确保精子核能够正确的成形和延伸,这对于精子头部的形成至关重要[18]。SPAG4主要在睾丸和胰腺中表达[19],该蛋白依靠亮氨酸拉链与人精子尾部蛋白Odf1相互作用,在精子尾部的形成以及精子运动方面发挥着重要作用[11]。Kracklauer等[20]研究发现,在雄性果蝇中 Spag4对于维持中心粒与细胞核的连接是必需的,如果将 Spag4基因敲除,那么在圆形精子和长形精子中细胞核将不能正确定位,出现核离散的现象,雄性果蝇出现不育。重新将 Spag4基因导入,则生殖功能恢复,其中SUN domain对于生殖功能恢复至关重要。
SPAG4L是我们研究小组首先成功克隆的,生物信息分析结果表明,SPAG4L与SPAG4虽然同属SUN蛋白家族,但是它们不是同一个蛋白,它们的氨基酸序列有45% 同源性。Chalmel F等[21]对精子发生中基因表达变化的研究发现,Spag4L基因是一个减数分裂的标志性基因。我们研究发现,Spag4L基因在小鼠出生后3周开始高表达[14]。郭睿等[22]分离纯化了小鼠的各级生精细胞,对Spag4L进行了荧光定量PCR检测,结果发现,从粗线期精母细胞阶段到长形精子细胞阶段Spag4L表达下调,在圆形和长形精子细胞中其表达量分别下降了33%和45%。这些结果说明,在小鼠减数分裂中Spag4L蛋白发挥着重要的功能。通过对 SPAG4L及其突变体的定位发现,SPAG4L主要定位于细胞核膜和内质网。根据扩散-滞留模型(“Diffusion-Retention” model)理论[10],从理论上分析,SPAG4L蛋白必须与内核膜蛋白或核质蛋白相互作用才能保证在核膜上的锚定。
在前期研究中,我们虽然成功对 SPAG4L的 C末端进行了原核表达,但是该融合蛋白不能用于研究SPAG4L的N端功能。生物信息分析结果表明,SPAG4L C端朝向胞质,N端朝向核质,靠近N端的地方有跨膜区,前期研究中,我们尝试用不同的原核表达载体对 SPAG4L全长进行原核表达,结果均未成功。因此,我们尝试将 SPAG4L放到 CMV启动子下游进行表达,同时,希望所表达的融合蛋白带有Myc和His双标签,以利于后续的实验研究。本研究中,我们选择了真核表达载体 pcDNA3.1/myc-His(-)A,成功构建了 pcDNA3.1/myc-His(-)A/SPAG4L真核表达质粒,通过体外转染细胞,筛选获得稳定转染细胞系,这些细胞系能够成功表达带Myc、His标签的SPAG4L全长蛋白。这样,我们下一步可以通过6×His标签对SPAG4L融合蛋白进行纯化,通过pull-down实验研究筛选SPAG4L相互作用蛋白。此外,Myc标签还可以用于免疫共沉淀实验,为研究 SPAG4L蛋白相互作用及信号通路提供了有力的工具。
总之,我们成功构建了带有His、Myc双标签的 pcDNA3.1/myc-His(-)A/SPAG4L真核表达质粒,并成功获得了稳定转染 pcDNA3.1/myc-His(-)A/SPAG4L的细胞系,为深入研究SPAG4L蛋白的功能及其相互作用蛋白奠定了基础。
[1]Chu DS, Shakes DC. Spermatogenesis. Adv Exp Med Biol, 2013, 757: 171–203.
[2]Chocu S, Calvel P, Rolland AD, et al.Spermatogenesis in mammals: proteomic insights.Syst Biol Reprod Med, 2012, 58(4): 179–190.
[3]Aston KI, Conrad DF. A review of genome-wide approaches to study the genetic basis for spermatogenic defects. Methods Mol Biol, 2013,927: 397–410.
[4]Vogt PH, Bender U. Human Y chromosome microdeletion analysis by PCR multiplex protocols identifying only clinically relevant AZF microdeletions.Methods Mol Biol, 2013, 927: 187–204.
[5]Behulova R, Strhakova L, Boronova I, et al. DNA analysis of Y chromosomal AZF region in Slovak population with fertility disorders. Bratisl Lek Listy, 2011, 112(4): 183–187.
[6]Yang S, Wang W, Lei C, et al. Localization and characterization of rat transmembrane protein 225 specifically expressed in testis. DNA Cell Biol,2011, 30(1): 9–16.
[7]Teng YN, Liao MH, Lin YB, et al. Expression of lrwd1 in mouse testis and its centrosomal localization. Int J Androl, 2010, 33(6): 832–840.
[8]Yu J, Lei K, Zhou M, et al. KASH protein Syne-2/Nesprin-2 and SUN proteins SUN1/2 mediate nuclear migration during mammalian retinal development. Hum Mol Genet, 2011, 20(6):1061–1073.
[9]Morimoto A, Shibuya H, Zhu X, et al. A conserved KASH domain protein associates with telomeres, SUN1, and dynactin during mammalian meiosis. J Cell Biol, 2012, 198(2): 165–172.
[10]Hasan S, Güttinger S, Mühlhäusser P, et al.Nuclear envelope localization of human UNC84A does not require nuclear lamins. FEBS Lett, 2006,580(5): 1263–1268.
[11]Shao X, Tarnasky HA, Lee JP, et al. Spag4, a novel sperm protein, binds outer dense-fiber protein Odf1 and localizes to microtubules of manchette and axoneme. Dev Biol, 1999, 211(1):109–123.
[12]Xing XW, Li LY, Fu JJ, et al. Cloning of cDNA of TSARG4, a human spermatogenesis related gene.Acta Biochim Biophys Sin, 2003, 35(3): 283–288(in Chinese).邢晓为, 李麓芸, 傅俊江, 等. 人类生精相关新基因TSARG4的cDNA 克隆. 生物化学与生物物理学报, 2003, 35(3): 283–288.
[13]XZ Jiang, MG Yang, LH Huang,et al. SPAG4L, a novel nuclear envelope protein involved in nuclear migration in meiotic stage of spermatogenesis.DNA Cell Biol, 2011, 30(11): 875–882.
[14]Xing XW, Li LY, Liu G, et al. Identification of a novel gene SRG4, expressed at specific stages of mouse spermatogenesis. Acta Biochim Biophys Sin, 2004, 36(5): 351–359.
[15]Frohnert C, Schweizer S, Hoyer-Fender S.SPAG4L/SPAG4L-2 are testis-specific SUN domain proteins restricted to the apical nuclear envelope of round spermatids facing the acrosome.Mol Hum Reprod, 2011, 17(4): 207–218.
[16]Jiang XZ, Yang MG, Xing XW. Prokaryotic expression and purification of SPAG4L, a novel human testis gene. J Southern Med Univ, 2010,30(9): 2047–2050 (in Chinese).蒋先镇, 杨明刚, 邢晓为. 人睾丸新基因SPAG4L的原核表达与纯化. 南方医科大学学报,2010, 30(9): 2047–2050.
[17]Ding X, Xu R, Yu J, et al. SUN1 is required for telomere attachment to nuclear envelope and gametogenesis in mice. Dev Cell, 2007, 12(6):863–872.
[18]Göb E, Schmitt J, Benavente R, et al. Mammalian sperm head formation involves different polarization of two novel LINC complexes. PLoS ONE, 2010, 5(8): e12072.
[19]Kennedy C, Sebire K, de Kretser DM, O'Bryan MK. Human sperm associated antigen 4 (SPAG4)is a potential cancer marker. Cell Tissue Res, 2004,315(2): 279–283.
[20]Kracklauer MP, Wiora HM, Deery WJ, et al. The Drosophila SUN protein Spag4 cooperates with the coiled-coil protein Yuri Gagarin to maintain association of the basal body and spermatid nucleus. J Cell Sci, 2010, 123(Pt 16): 2763–2772.
[21]Chalmel F, Rolland AD, Niederhauser-Wiederkehr C, et al. The conserved transcriptome in human and rodent male gametogenesis. Proc Natl Acad Sci USA, 2007, 104(20): 8346–8351.
[22]Guo R, Li XX, Wang HZ. Stage-specific expression analysis of mouse testis-specific genes in spermatogenic cells. Chin J Zool, 2009, 44(1):39–46 (in Chinese).郭睿, 李喜霞, 王惠珍. 小鼠睾丸特异基因在生精细胞中阶段性表达的定量分析. 动物学杂志,2009, 44(1): 39–46.
[23]Tapley EC, Ly N, Starr DA. Multiple mechanisms actively target the SUN protein UNC-84 to the inner nuclear membrane. Mol Biol Cell, 2011,22(10): 1739–1752.
