肝素微生物生产展望
2012-01-06陈祥娥凌沛学
陈祥娥,凌沛学
(1.山东大学 药学院,山东 济南 250012;2.山东省生物药物研究院 博士后科研工作站,山东 济南250101)
肝素微生物生产展望
陈祥娥1,2,凌沛学1,2
(1.山东大学 药学院,山东 济南 250012;2.山东省生物药物研究院 博士后科研工作站,山东 济南250101)
肝素作为重要的抗凝血药物在临床中广泛应用。近年来,动物来源肝素的质量问题成为人们关注的焦点,研究开发非动物源性肝素的大规模生产方式成为一大热点。采用与肝素生物合成前体具有相似结构的细菌荚膜多糖heparosan作为底物,已成功合成肝素、硫酸乙酰肝素,并获得多种具有肝素相关功能的肝素衍生物,是一种较有希望的非动物源性肝素生产方式。本文对此进行综述。
heparosan;肝素;微生物发酵
肝素(HP)被发现已90余年,作为抗凝血和抗血栓药物广为应用。HP还具有降血脂、抗炎、抗过敏、抗动脉粥样硬化、抗肿瘤、抑制细菌黏附等作用[1-2],是重要的糖胺聚糖(GAG)类生化药物之一。目前,商品化HP主要是从肠黏膜等组织中提取。2008年在HP使用中,曾因杂质过量出现严重的过敏反应甚至死亡[3-4]。动物源性HP的质量成为广泛关注的焦点,同时也引发了对非动物来源HP的需求。
微生物多糖的生产以其潜在的生物工程应用,得到广泛关注。已知有多种细菌可以产生含GAG样聚合物的荚膜多糖[5]。目前,透明质酸(HA)已实现细菌发酵生产,而HP等其他重要GAG的大规模微生物发酵仍处在探索阶段。
动物体内HP的二糖单位主要为葡糖胺(GlcN)和艾杜糖醛酸(IdoUA),GlcN的氨基氮和C6位均带有硫酸基。HP合成时二糖中是葡糖醛酸(GlcUA),然后差向异构化为IdoUA,并随之进行C2位硫酸化。脊椎动物合成HP的过程是先催化合成多糖heparosan作为前体,然后进行一系列的硫酸化及异构化修饰[6]。细菌heparosan的发现,引起了广泛关注。因为它使通过细菌合成,化学或生物修饰生产HP、硫酸乙酰肝素(HS)成为可能。本文就以heparosan为前体的HP类微生物发酵生产作一综述。
1 heparosan
肝素前体heparosan又称 N-acetylheparosan,(-GlcUA-1,4-GlcNAc-1,4-)n(其中GlcUA代表葡糖醛酸;GlcNAc代表乙酰葡糖胺),是某些细菌荚膜中多糖骨架的二糖重复单位,同时也作为HP和HS的生物合成前体,在水螅和脊椎动物中都有发现[7]。其二糖骨架结构虽与脊椎动物中的HP类似,但未硫酸化,也没有将GlcUA异构化为IdoUA。

图1 heparosan二糖单位的结构(A)和HP的主要和可变重复二糖单位(B)
采用生物工程的方法,以heparosan为前体合成HP提供了一种新的、更为安全且较有希望的非动物源性生产方式[8]。目前发现的荚膜多糖中含heparosan的细菌有大肠杆菌K5(E.coli K5)和多杀性巴氏杆菌D型(P.multocida type D)。其中P.multocida产生的heparosan平均相对分子质量(Mr)较高,为 2.0 ×105~3.0 ×105,而 E.coli K5 heparosan(K5PS)大小与 HP接近,Mr为1.0 ×104~2.0×104。所以,以K5PS为HP、HS生物合成前体的研究开展较多,应用也较为广泛。研究表明,从E.coli K5获得的重均分子质量(Mw)1.0×104以上的heparosan可在酶的作用下转化成类似于HP的具有抗凝血活性的多糖[9-10]。
2 合成方法
脊椎动物HS的生物合成过程是先催化合成GlcUAGlcNAc二糖重复单位组成的heparosan多糖骨架。经N-去乙酰化酶/硫酸转移酶(NDST)作用,将GlcNAc转化成N-磺基葡糖胺(GlcNS)。N-硫酸化后,C5-异构化酶(C5-epi)再将部分GlcUA转化成IdoUA。所得多糖经2-O-硫酸转移酶(2-OST),6-O-硫酸转移酶(6-OST)和3-O-硫酸转移酶(3-OST)的修饰分别将磺基加入IdoUA和GlcUA的C2位以及GlcN的C6位和C3位。虽然,酶修饰不一定遵循上述顺序,但NDST对GlcNAc的作用决定异构化和O-硫酸化的水平[11]。
以heparosan为前体,酶为基础的HP、HS合成是一种较有希望的非动物源性生产方式。与化学合成法相比,化学-酶法合成具有HP、HS生物合成酶的高度区域选择性,在合成硫酸化多糖和较大低聚糖中具有显著优势。HS生物合成途径的拟态化学-酶法或全酶法合成可产生具有特殊生物活性的多种 HS 结构[12]。
研究认为,HP类产品的抗凝血活性主要依赖于其多糖链中能与抗凝血酶(AT)结合的五糖序列。Lindahl等[9]以K5PS为底物,化学法N-去乙酰化和N-硫酸化后,经C5-epi作用,将部分GlcUA转化为IdoUA,所得异构化N-硫酸化K5PS与N,N-二甲基甲酰胺和吡啶三氧化硫反应,实现全部O-硫酸化,二甲亚砜-甲醇(9∶1)选择性去硫酸化后,再进行GlcN 6-O-硫酸化及N-硫酸化,最终获得可与AT结合的特殊“neoheparin”。Kuberan等[13]则采用全酶法合成了ATⅢ结合五糖。通过选择适当的生物合成用酶相组合,也可得到具有HP类似结构及抗凝血活性的硫酸化多糖[14]。
HP、HS生物合成过程中所涉及的特异酶已基本克隆得到,已在E.coli中成功表达16种参与其生物合成的酶(部分见表1)。可实现以 heparosan为前体,经 NDST、C5-epi、2-OST、3-OST、6-OST 等酶修饰,合成 HP 和 HS[15]。

表1 E.coli中表达的部分HS生物合成用酶
以heparosan为前体合成HP及其类似物首要解决的就是heparosan的产量问题。Wang等[16]采用优化培养基,富氧给予并指数补加葡萄糖,在7 L发酵罐中培养E.coli K5菌株制备 heparosan,产量可达15 g/L,Mr为5.8 ×104,Mw为8.4×104。此外,还可联合应用发酵工程、基因工程和代谢工程等,以获得具有理想结构特性及较高产量的heparosan,作为生物工程HP生产前体[17]。
3 应用
以heparosan为前体可合成HP、HS及多种衍生物,heparosan及其衍生物具有与HP类似的结构,其硫酸化衍生物亦有许多与HP类似的生物活性,主要有抗凝血、调节细胞因子及抗病毒作用等。以K5PS为前体合成的HP类衍生物见图2。
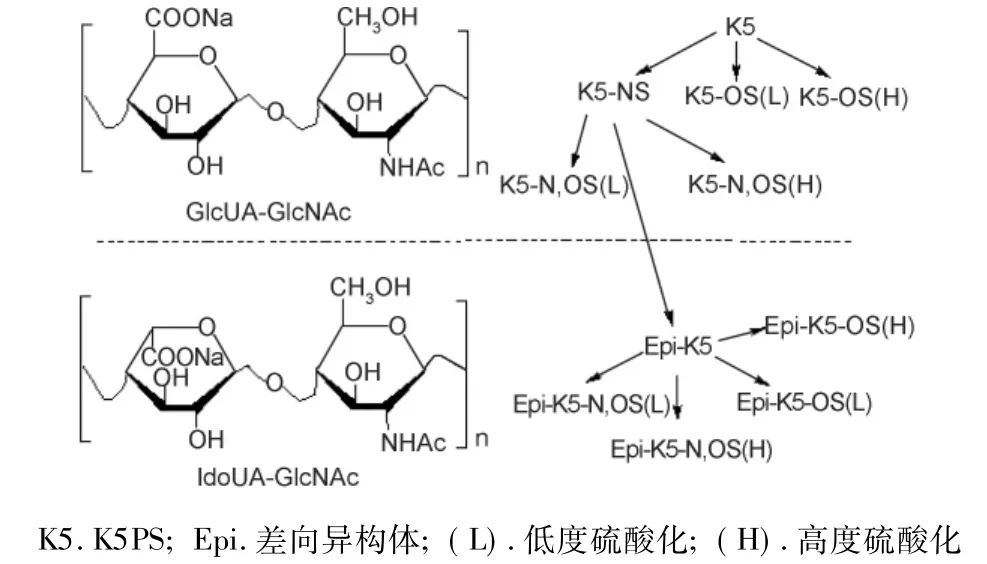
图2 K5荚膜多糖及其衍生物的化学结构
研究发现,未经修饰的heparosan可抑制人乳头状瘤病毒16(HPV-16)假病毒颗粒与固定化HP间的相互作用,高度O-硫酸化、N,O-硫酸化的多糖衍生物则具有抗成纤维细胞生长因子2(FGF-2)活性,抑制人类免疫缺陷病毒(HIV)、单纯疱疹病毒(HSV)和HPV的感染作用等。磺胺heparosan则具有与HP类似的抗凝血和抗蛋白酶活性,能释放组织因子通道抑制剂,抑制组织因子介导的血小板激活作用[18]。
Munoz等[19]将生物素酰化的K5PS固定在传感器芯片上,用参与HP生物合成的酶选择性修饰获得与HP结构类似的多糖,通过检测这些多糖与ATⅢ的表面等离子体共振确定其相互作用,实现HP-蛋白质相互作用的快速筛选,并可提供参与结合的关键硫酸化基团的结构信息。
4 展望
近年来,HP的非动物源性生产研究较多,有化学法[20]、中国仓鼠卵巢(CHO)细胞代谢工程[21-22]等多种方法,其中以heparosan为前体,酶修饰为基础的HP合成方式是一种较有希望的替代方法,可产生具有特殊生物活性的多种HP产品。相信随着研究的不断深入,可实现HP的大规模产业化微生物生产。
[1]Kreuger J,Spillmann D,Li J P,et al.Interactions between heparan sulfate and proteins:the concept of specificity[J].J Cell Biol,2006,174(3):323-327.
[2]Gu L,Wang H,Guo Y L,et al.Heparin blocks the adhesive of E.coli O157:H7 to human colonic epithelial cells[J].Food Control,2008,19(12):1119-1125.
[3]Liu H,Zhang Z,Linhardt R J.Lessons learned from the contamination of heparin[J].Nat Prod Rep,2009,26:313-321.
[4]Guerrini M,Beccati D,Shriver Z,et al.Oversulfated chondroitin sulfate is a contaminant in heparin associated with adverse clinical events[J].Nat Biotechnol,2008,26(6):669-675.
[5]DeAngelis P L.Microbial glycosaminoglycan glycosyltransferases[J].Glycobiology,2002,12(1):9R-16R.
[6]Busse M,Feta A,Presto J,et al.Contribution of EXT1,EXT2,and EXT3 to heparan sulfate chain elongation[J].JBiol Chem,2007,282(45):32802-32810.
[7]陈祥娥,凌沛学,张天民.细菌肝素前体heparosan合酶[J].中国生化药物杂志,2008,29(6):422-424.
[8]Laremore TN,Zhang F,Dordick JS,et al.Recent progress and applications in glycosaminoglycan and heparin research[J].Curr Opin Chem Biol,2009,13(5-6):633-640.
[9]Lindahl U,Li JP,Kusche-Gullberg M,et al.Generation of“Neoheparin”from E.coli K5 capsular polysaccharide[J].J Med Chem,2005,48(2):349-352.
[10] Zhang Z Q,McCallum SA,Xie J,et al.Solution structures of chemoenzymatically synthesized heparin and its precursors[J].J Am Chem Soc,2008,130(39):12998-13007.
[11] Kusche M,Hannesson H H,Lindahl U.Biosynthesis of heparin.Use of Escherichia coli K5 capsular polysaccharide as a model substrate in enzymic polymer-modification reactions[J].Biochem J,1991,275(Pt 1):151-158.
[12] Chen J,Avci F Y,Munoz E M,et al.Enzymatic redesigning of biologically active heparan sulfate[J].JBiol Chem,2005,280(52):42817-42825.
[13] Kuberan B,Lech M Z,Beeler D L,et al.Enzymatic synthesis of antithrombin III binding heparan sulfate pentasaccharide[J].Nat Biotechnol,2003,21(11):1343-1346.
[14] Chen J,Jones C L,Liu J.Using an enzymatic combinatorial approach to identify anticoagulant heparan sulfate structures[J].Chem Biol,2007,14(9):986-993.
[15] Peterson S,Frick A,Liu J.Design of biologically active heparan sulfate and heparin using an enzyme-based approach[J].Nat Prod Rep,2009,26(5):610-627.
[16] Wang Z Y,Ly M,Zhang F,et al.E.coli K5 fermentation and the preparation of heparosan,a bioengineered heparin precursor[J].Biotechnol Bioeng,2010,107(6):964-973.
[17] Wang Z,Dordick J S,Linhardt R J.Escherichia coli K5 heparosan fermentation and improvement by genetic engineering[J].Bioeng Bugs,2011,2(1):63-67.
[18]陈祥娥,凌沛学.大肠杆菌K5荚膜多糖硫酸化衍生物的应用[J].中国生化药物杂志,2009,30(6):427-429.
[19] Munoz E,Xu D,Avci F,et al.Enzymatic synthesis of heparin related polysaccharides on sensor chips:rapid screening of heparin-protein interactions[J].Biochem Biophys Res Commun,2006,339(2):597-602.
[20] Petitou M,van Boeckel C A.A synthetic antithrombin Ⅲ binding pentasaccharide is now a drug!What comes next? [J].Angew Chem Int Ed Engl,2004,43(24):3118-3133.
[21] Bame K J,Lidholt K,Lindahl U,et al.Biosynthesis of heparan sulfate.coordination of polymermodification reactions in a Chinese hamster ovary cell mutant defective in N-sulfotransferase[J].JBiol Chem,1991,266(16):10287-10293.
[22] Zhang L,Lawrence R,Frazier B A,et al.CHO glycosylation mutants:proteoglycans[J].Methods Enzymol,2006,416:205-221.
Progress on microbial production of heparin
CHEN Xiang-e1,2,LING Pei-xue1,2
(1.School of Pharmaceutical Science,Shandong University,Jinan 250012,China;2.Postdoctoral Scientific Research Workstation,Institute of Biopharmaceuticals of Shandong Province,Jinan 250101,China)
TQ929
A
1005-1678(2012)04-0502-03
2011-07-19
陈祥娥,女,博士研究生,微生物与生化药学专业;凌沛学,通信作者,研究员,博士生导师,Tel:0531-81213003,E-mail:peixue.ling@bausch.com。
