CC10蛋白功能及在疾病中的作用研究进展
2011-12-01魏颖徐玉东尹磊淼单纯筱冉君王宇刘晓燕杨永清
魏颖,徐玉东,尹磊淼,单纯筱,冉君,王宇,刘晓燕,杨永清
Clara 细胞是一种分布于哺乳动物肺脏末端细支气管的非纤毛、非浆液分泌上皮细胞,具有多种生物学功能。在气管支气管树分叉处的Clara 细胞是支气管上皮细胞的祖细胞,有助于损伤后上皮再生[1-2]。Clara 细胞 10-kD蛋白(clara cell 10-kD protein,CC10)是Clara 细胞分泌的一种分子量为10-kD的分泌蛋白。其通过抑制磷脂酶 A2(phospholipase A2,PLA2)活性发挥抗炎和免疫调节活性[3-4]。CC10 最初因出现在兔生殖管道着床期,被称为子宫球蛋白(uteroglobin,blastokinin)。除此之外,CC10还有多种别名,如 CCSP(Clara cell secretory protein)、CC16(Clara cell 16-kD protein)、UP-1(urine protein 1)、CCPBP(Clara cell phospholipid-binding protein)、SCGB1A1(Secretoglobin family 1A member 1)等。
1 生物学特性
CC10 由两个 70~77个氨基酸多肽反向平行构成的二聚体形成。其分子量由氨基酸序列决定,在SDS 聚丙烯酰胺凝胶中具有高迁移率,分子量为10~12 kD。因此在最初的描述中,该蛋白被称为CC10蛋白。CC10 单体包含4个 α 螺旋和一个 β 折叠,β 折叠将 4个 α 螺旋隔开。CC10 二聚体的8个 α 螺旋共同构成一个疏水口袋,位于疏水口袋内部的氨基酸残基高度保守。纯化人CC10蛋白 X 射线结晶衍射显示类脂占据疏水腔内部。气相色谱法分析该蛋白疏水腔内类脂抽提物显示该蛋白对磷脂酰胆碱(phosphatidylcholine,PC)和磷脂酰肌醇(phosphatidylinositol,PI)的结合量几乎相等[5]。CC10结合 PC和PI 活性可能与表面活性脂质和肺部其他疏水分子转运有关。CC10 通过结合 PC和PI 抑制 PLA2 活性,发挥抗炎功能[6]。
人 CC10 基因位于染色体 11q12.2-13.1 区域,该区域也编码其他具有抗炎活性蛋白的基因[7]。CC10能够结合钙离子,其某些功能可能由该蛋白钙离子螯合活性介导。CC10钙离子结合位点形成钙离子配体的原子空间结构与分泌性PLA2的主要钙离子结合位点空间结构相似[8]。
2 组织细胞分布
人 CC10蛋白存在于支气管树非纤毛、非浆液、非黏膜柱状细胞的分泌颗粒中。该蛋白出现于人类肺脏的形成过程中,可通过免疫组化验查在妊娠期 21周或者羊水中出现[9]。CC10 主要产生部位在呼吸系统黏膜,提示其主要参与呼吸道炎症调节。CC10 在恒河猴气道的分布研究显示,其遍布于气道上皮非纤毛细胞,上气道和中气道表达量最多,末梢气道较少,末端细支气管几乎不表达 CC10[10]。CC10 也表达于胸腺、垂体腺、胃肠道、胰腺、前列腺以及精囊[11]。Cecchini等[12]研究了 CC10 在唾液腺的表达分布情况,发现 CC10 在舌下腺管道部分表达,在腮腺和颌下腺没有表达。
3 临床应用
3.1 对肿瘤细胞作用
人肺部肿瘤测定表明 10%~30% 肺部腺癌显示出病灶局部 CC10 染色,且恶性肿瘤细胞染色较正常 Clara 细胞染色浅[13]。仓鼠肿瘤模型显示 CC10 染色仅在肿瘤早期出现,这些肿瘤细胞进一步分化为不表达 CC10的鳞状细胞癌[14-15]。B16F10 黑素瘤细胞尾静脉注射 CC10 基因敲除大鼠,发现与野生型同窝出生仔鼠相比,肺部肿瘤集落明显增多。研究进一步发现 CC10 基因敲除大鼠肺脏过表达两种钙结合蛋白 S100A8和S100A9,B16F10 表达晚期糖基化终产物受体(advanced glycosylation end product-specific receptor,RAGE),S100A8和S100A9 既是表达 RAGE的B16F10 细胞的趋化剂,也作为配体与该受体结合。用RAGE 抗体预处理 B16F10 细胞能够抑制肿瘤迁移和侵润。研究还发现 CC10 基因敲除大鼠肺脏 S100A8/S100A9浓度最高,尾静脉浓度最低,提示 B16F10 细胞迁移至肺脏,表明 CC10 抗炎蛋白缺乏可引起肺脏黑素瘤细胞集落增加[16]。此外,与正常上皮细胞相比,前列腺癌细胞 CC10降低,表明该蛋白具有抗肿瘤增长活性。体外研究显示,CC10能够抑制肿瘤细胞穿透基质,提示其可能成为恶性肿瘤抑制剂[17-18]。Harvilchuck和Carlson[19]研究显示苯乙烯及其代谢产物(R-styrene oxide,R-SO)能够显著降低 CC10水平。Clara 细胞是苯乙烯肺毒性的主要靶标,认为逐渐降低的CC10 水平可能引起 Clara 细胞氧化应激增加,提示CC10 可能是肺部肿瘤的早期指示剂。
3.2 对炎症作用
动物模型研究表明 CC10 对炎症和纤维变性肺疾病具有保护作用[20-21],人肺部 CC10 表达降低与肺部炎症相关[14]。对大鼠研究表明细菌脂多糖(lipopolysaccharides,LPS)诱发的急性肺部炎症中 CC10 合成与分泌显著降低,CC10基因缺失与肺部病毒感染易感性增强和氧化应激相关[22-23]。对具有急性呼吸窘迫综合征(acute respiratory distress syndrome,ARDS)危险因素受试者支气管肺泡灌洗液(bronchoalveolar lavage fluid,BALF)研究表明 CC10 浓度与弹性蛋白酶量呈逆相关,CC10蛋白浓度降低与中性粒细胞介导的肺损伤相关,提示 CC10 可能是天然中性粒细胞抑制剂[24],CC10 浓度降低可能有助于外周呼吸道炎性发生,在导致气道重塑和不可逆性气道阻塞的一系列事件中起作用。
研究显示 CC10能保护呼吸道免受氧化应激和炎症损害[25]。吸烟者和慢性阻塞性肺病(chronic obstructive pulmonary disease,COPD)患者 BALF 中 CC10表达显著下调,重度 COPD 患者 CC10 水平较中度患者明显降低,表明在COPD 不同阶段,CC10 表达差异显著[26]。
CC10 具有抗细胞因子活性,其可抑制 IFN-γ、 IL-1和TNF-α 产生。CC10显示出对 IFN-γ 引起的抗病毒活性以及吞噬增强活性的抑制作用,CC10 抑制白细胞趋化活性[27]。研究显示肺部上皮穿透性完整时,血浆 CC10 水平与BALF一致[28],婴儿血浆 CC10 水平增高可能与呼吸道感染引起上皮损伤有关。CC10 与中性粒细胞相互关系研究显示,具有肺部炎症的马支气管肺泡灌洗液中性粒细胞含有丰富的CC10,且免疫电子显微镜定位显示主要存在于胞质和细胞核中。CC10 与马血中性粒细胞孵育显著降低中性粒细胞氧化爆发活性,增加其吞噬作用,提示该蛋白能够进入马肺部炎症中性粒细胞并改变其功能[29]。研究显示早期重组人 CC10蛋白(recombinant human CC10,rhCC10)干预能够增强表面活性剂疗法对呼吸窘迫综合征气体交换和肺稳定性的改善作用[30]。
3.3 对过敏性疾病作用
SELDI-TOF-MS 质谱技术分析小鼠哮喘模型肺组织显示 CC10是哮喘病理生理机制潜在靶蛋白之一[31]。研究显示人体 CC10 水平降低与哮喘相关[32]。抗原致敏大鼠体外实验研究发现,CC10 剂量依赖的显著抑制 Th2 细胞因子表达,但对 IFN-γ 没有抑制作用。同样发现其对 CD4+ Th2细胞具有相似的抑制作用,但是对初始 CD4+ T 细胞没有抑制作用。相反,CC10能够诱导初始 CD4+ T 细胞表达IFN-γ,对极化的Th2 细胞却没有此作用。此外,CC10 对Th2 细胞因子表达的抑制作用与一种重要的转录因子GATA-3 表达降低共存。体内研究表明,抗原激发后CC10缺失大鼠基因重组引起 Th2 细胞因子水平显著降低,伴随GATA-3 表达显著降低,伴随肺部嗜酸性粒细胞水平降低。CC10 对于调节 T 细胞反应具有直接作用,可能通过抑制GATA-3 依赖性的Th2 细胞因子产生抑制 Th2 细胞的功能[33]。研究显示与野生型大鼠相比,致敏和OVA激发后CC10 缺失大鼠 IL-4、 IL-5和IL-13 表达显著增加[34]。此外,CC10 缺失大鼠肺部炎症反应明显增强,将 CC10 加入大鼠 Th2 细胞株显示出下调 IL-4、IL-5和IL-13 表达的作用[33]。
研究显示 CC10 通过 PLA2发挥抗炎活性[35-36]。CC10通过抑制 PLA2和前列素 D2(prostaglandin D2,PGD2)直接发挥抗炎作用。PLA2和PGD2均参与花生四烯酸炎症级联反应[37-38]。PGD2主要由存在于活化肥大细胞内的环氧合酶(COX)产生,具有多种生物活性,包括血管舒张,支气管收缩以及抑制血小板聚集[39]。王秀芳等[40]研究发现5 岁以下哮喘儿童血清 CC10 水平降低,具有遗传性过敏症高风险的哮喘患儿 CC10 水平下降更明显,血清 IgE 水平显著增高,认为具有遗传性过敏症高风险的哮喘患儿血清CC10 水平与IgE 水平存在负相关关系。
过敏性鼻炎儿童鼻灌洗液中 CC10 水平较同年龄正常儿童低[41]。有人对骨桥蛋白在过敏性鼻炎中的作用及其与CC10的关系进行了研究,显示骨桥蛋白在小鼠过敏性鼻炎模型中表达上调,同时 CC10 表达下调;CC10 基因敲除小鼠过敏性鼻炎模型中骨桥蛋白表达上调更为明显。致敏和激发期间给予重组CC10蛋白显著改善 Th2 炎症和减少鼻黏膜骨桥蛋白表达[42]。人 CC10 基因非编码区遗传变异会影响血浆 CC10 水平。38A 基因序列与CC10 水平降低相关,CC10 水平偏低人群具有高哮喘易感性。CC10可能通过抗炎活性发挥抗哮喘作用[43]。除了肺部上皮细胞,CC10也表达于鼠上气道上皮细胞。嗜酸性慢性鼻窦炎(eosinophilic chronic rhinosinusitis,ECRS)发展过程中,CC10 表达逐渐下调。ECRS 形成后,CC10 基因敲除大鼠出现同样的嗜酸性 Th2 反应加重现象,而重组CC10 可以抑制该现象。CC10 缺乏会引起更严重的组织重塑,而组织重塑是慢性气道炎症的基本特点之一。基因芯片研究发现几丁质酶 3 样 1(chitinase 3-like 1,CHI3L1)是 ECRS 发病机制中表达上调最明显的基因之一,其在哮喘病人肺组织和血清中表达增高并与哮喘严重程度相关[44-45],重组CC10可以显著抑制 CHI3L1 表达。CC10 可能通过调节 CHI3L1表达发挥对气道炎症的调节作用[46]。Widegren等[47]研究发现 rhCC10 以 0.56 mg 鼻腔给药对季节性过敏性鼻炎没有抗过敏效应。Liu等[48]研究发现慢性鼻窦炎患者鼻窦黏膜CC10 水平显著降低,同时伴随鼻息肉和哮喘的慢性鼻窦炎患者 CC10 水平降低更明显。CC10 水平与鼻息肉术前 CT分值以及术后内窥镜检查和症状分值呈负相关关系。TNF-α、IL-1β和IL-4 抑制鼻黏膜 CC10 产生,INF-γ和IL-10 促进 CC10 产生。认为CC10 参与慢性鼻窦炎病理生理机制,且与疾病严重程度相关。Zhang等[49]研究了中国汉族人群 CC10 基因变异与慢性鼻窦炎发病的关系,认为CC10 A + 38G 单核苷酸多态性对汉族人群慢性鼻窦炎发病可能不存在实质作用。
3.4 对泌尿生殖系统疾病的作用
CC10 作为分泌蛋白存在于人输卵管,盆腔炎症性疾病和输卵管异位妊娠患者输卵管 CC10 表达上调,提示其参与输卵管免疫抑制反应[50]。
IgA 肾病(IgA-nephropathy,IgAN)患者循环 IgA-Fn(纤连蛋白)复合体水平增高,使 IgA 与Fn 在肾小球异常沉积进而引起肾衰。CC10能够竞争性与存在于Fn 上的肝素结合位点结合,抑制 Fn-IgA 异聚化,降低 IgA 肾病患者循环 IgA- Fn 复合体水平[51]。
4 与其他蛋白分子的相互作用
纤连蛋白在内皮细胞迁移和血管生成中起重要作用。rhCC10能够通过转谷酰胺酶(transglutaminase,TGase)共价结合人原代微血管内皮细胞(HMVEC)表面内源和外源 Fn。在rhCC10 与Fn 相互作用的固相期,rhCC10 显著抑制 HMVEC 对 Fn的依附作用,而对其他胞外基质蛋白没有此作用。此外,体外研究表明 rhCC10 剂量依赖性抑制细胞迁移,阻止创伤愈合[52]。CC10能够抑制某些肿瘤细胞迁移入侵,研究显示其能够与HTB-81、HTB-30和HTB-174等肿瘤细胞株表面的脂钙蛋白-1受体(Lip-1R)相互作用,发挥对肿瘤细胞迁移和入侵的抑制作用[53]。
CC10能够抑制 NF-кB 核转位,抑制 NF-кB DNA结合活性及转录活性[54]。
5 作为疾病生物标记
研究显示 CC10 对 ALI/ARDS 患者具有很好的诊断价值。血浆 CC10 水平快速增高 30% 或更高对诊断ALI/ARDS 有 90% 敏感性和92% 特异性,且CC10 水平在ALI/ARDS 诊断前 2 天增高,提示血浆 CC10 可作为呼吸机相关肺炎 ALI/ARDS 患者潜在的生物标记[55]。与心源性肺水肿相比,CC10 水平在ALI/ARDS 患者血浆及肺部水肿液中明显降低,在帮助鉴别诊断 ALI/ARDS和心源性肺水肿方面是有前景的生物标记物[56]。哮喘儿童血清CC10 水平明显降低,提示其可能作为儿童哮喘的有效诊断指标[57]。Hasegawa等[58]研究发现系统性硬化患者血清CC10 水平较系统性红斑狼疮患者和正常受试者高,但差异不具有统计学意义。CC10 水平增高与合并肺纤维化有关。操作性特征曲线分析显示,CC10 水平增高对于检测系统性硬化患者是否合并肺纤维化与SP-D 具有相同的意义。认为CC10 水平可作为诊断系统性硬化患者合并肺纤维化的潜在血清标记物。
6 展望
大量研究结果显示 CC10 具有抗炎、免疫调节以及抗肿瘤活性,但是相关作用机制仍不明确。STRING8.3是已知和预测蛋白相互作用的数据库,目前整合了 630 种生物体近 2, 590, 259 种蛋白质。STRING8.3 中相互作用分为直接(生理)和间接(功能)关联,蛋白质数据来源包括基因组、高通量实验、共表达以及先前知识。通过 STRING8.3 数据库分析(图 1)表明:CC10 与CUBN、LRP2、LRPAP1等膜受体存在直接或间接关联,与CEBPD、FOXA1、SPDEF、TFF3、FOXA2、TITF1、SFTP1、RB1等转录因子具有相关关系,与IL5、IL13等 Th2 细胞因子具有直接相关关系。推测 CC10 可能通过膜受体将信号传入细胞内,通过转录因子低表达相应炎性介质介导抗炎、抗肿瘤效应。在今后的研究中,应充分利用现代生物信息学、分子生物学以及细胞生物学技术深入研究 CC10 在疾病发病机制中的作用,明确相关分子相互作用关系,揭示相关生物信号传导途径。
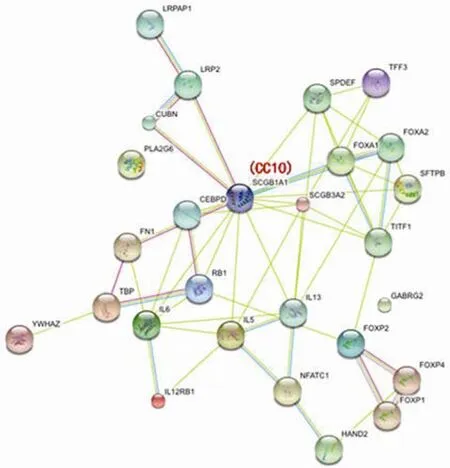
图1 CC10 与其他蛋白相互关系图
[1]Hermans C, Knoops B, Wiedig M, et al.Clara cell protein as a marker of Clara cell damage and bronchoalveolar blood barrier permeability.Eur Respir J, 1999, 13(5):1014-1021.
[2]Pilette C, Godding V, Kiss R, et al.Reduced epithelial expression of secretory component in small airways correlates with airflow obstruction in chronic obstructive pulmonary disease.Am J Respir Crit Care Med, 2001, 163(1):185-194.
[3]Miele L, Cordella-Miele E, Mukherjee AB.Uteroglobin: structure,molecular biology, and new perspectives on its function as a phospholipase A2 inhibitor.Endocr Rev, 1987, 8(4):474-490.
[4]Levin SW, Butler JD, Schumacher UK, et al.Uteroglobin inhibits phospholipase A2 activity.Life Sci, 1986, 38(20):1813-1819.
[5]Umland TC, Swaminathan S, Singh G, et al.Structure of a human Clara cell phospholipid-binding protein-ligand complex at 1.9 A resolution.Nat Struct Biol, 1994, 1(8):538-545.
[6]Andersson O, Nordlund-Möller L, Brönnegård M, et al.Purification and level of expression in bronchoalveolar lavage of a human polychlorinated biphenyl (PCB)-binding protein: evidence for a structural and functional kinship to the multihormonally regulated protein uteroglobin.Am J Respir Cell Mol Biol, 1991, 5(1):6-12.
[7]Hay JG, Danel C, Chu CS, et al.Human CC10 gene expression in airway epithelium and subchromosomal locus suggest linkage to airway disease.Am J Physiol, 1995, 268(4 Pt 1):L565- L575.
[8]Barnes HJ, Nordlund-Möller L, Nord M, et al.Structural basis for calcium binding by uteroglobins.J Mol Biol, 1996, 256(2):392-404.
[9]Singh G, Katyal SL.Clara cell proteins.Ann N Y Acad Sci, 2000,923:43-58.
[10]Coppens JT, Van Winkle LS, Pinkerton K, et al.Distribution of Clara cell secretory protein expression in the tracheobronchial airways of rhesus monkeys.Am J Physiol Lung Cell Mol Physiol, 2007,292(5):L1155- L1162.
[11]Peri A, Cordella-Miele E, Miele L, et al.Tissue-specific expression of the gene coding for human Clara cell 10-kD protein, a phospholipase A2-inhibitory protein.J Clin Invest, 1993, 92(5):2099-2109.
[12]Cecchini MP, Merigo F, Cristofoletti M, et al.Immunohistochemical localization of Clara cell secretory proteins (CC10-CC26) and Annexin-1 protein in rat major salivary glands.J Anat, 2009, 214(5):752-758.
[13]Singh G, Singh J, Katyal SL, et al.Identification, cellular localization,isolation, and characterization of human Clara cell-specific 10 KD protein.J Histochem Cytochem, 1988, 36(1):73-80.
[14]Broers JL, Jensen SM, Travis WD, et al.Expression of surfactant associated protein-A and Clara cell 10 kilodalton mRNA in neoplastic and non-neoplastic human lung tissue as detected by in situ hybridization.Lab Invest, 1992, 66(3):337-346.
[15]Herbert RA, Stegelmeier BS, Gillett NA, et al.Plutonium-induced proliferative lesions and pulmonary epithelial neoplasms in the rat:immunohistochemical and ultrastructural evidence for their origin from type II pneumocytes.Vet Pathol, 1994, 31(3):366-374.
[16]Saha A, Lee YC, Zhang Z, et al.Lack of an endogenous anti-inflammatory protein in mice enhances colonization of B16F10 melanoma cells in the lungs.J Biol Chem, 2010, 285(14):10822-10831.
[17]Kundu GC, Mantile G, Miele L, et al.Recombinant human uteroglobin suppresses cellular invasiveness via a novel class of high-affinity cell surface binding site.Proc Natl Acad Sci U S A, 1996,93(7):2915-2919.
[18]Weeraratna AT, Cajigas JA, Schwartz A, et al.Loss of uteroglobin expression in prostate cancer: relationship to advancing grade.Clin Cancer Res, 1997, 3(12 Pt 1):2295-2300.
[19]Harvilchuck JA, Carlson GP.Effect of multiple doses of styrene and R-styrene oxide on CC10, bax, and bcl-2 expression in isolated Clara cells of CD-1 mice.Toxicology, 2009, 259(3):149-152.
[20]Chen LC, Zhang Z, Myers AC, et al.Cutting edge: altered pulmonary eosinophilic inflammation in mice deficient for Clara cell secretory 10-kDa protein.J Immunol, 2001, 167(6):3025-3028.
[21]Lee YC, Zhang Z, Mukherjee AB.Mice lacking uteroglobin are highly susceptible to developing pulmonary fibrosis.FEBS Lett, 2006,580(18):4515-4520.
[22]Arsalane K, Broeckaert F, Knoops B, et al.Clara cell specific protein(CC16) expression after acute lung inflammation induced by intratracheal lipopolysaccharide administration.Am J Respir Crit Care Med, 2000, 161(5):1624-1630.
[23]Geerts L, Jorens PG, Willems J, et al.Natural inhibitors of neutrophil function in acute respiratory distress syndrome.Crit Care Med, 2001,29(10):1920-1924.
[24]Hawgood S, Clements JA.Pulmonary surfactant and its apoproteins.J Clin Invest, 1990, 86(1):1-6.
[25]Broeckaert F, Clippe A, Wattiez R, et al.Lung hyperpermeability,Clara-cell secretory potein (CC16), and susceptibility to ozone of five inbred strains of mice.Inhal Toxicol, 2003, 15(12):1209-1230.
[26]Braido F, Riccio AM, Guerra L, et al.Clara cell 16 protein in COPD sputum: a marker of small airways damage? Respir Med, 2007,101(10):2119-2124.
[27]Mukherjee AB, Kundu GC, Mantile-Selvaggi G, et al.Uteroglobin: a novel cytokine? Cell Mol Life Sci, 1999, 55(5):771-787.
[28]Broeckaert F, Arsalane K, Hermans C, et al.Serum clara cell protein:a sensitive biomarker of increased lung epithelium permeability caused by ambient ozone.Environ Health Perspect, 2000, 108(6):533-537.
[29]Katavolos P, Ackerley CA, Clark ME, et al.Clara cell secretory protein increases phagocytic and decreases oxidative activity of neutrophils.Vet Immunol Immunopathol, 2011, 139(1):1-9.
[30]Shashikant BN, Miller TL, Welch RW, et al.Dose response to rhCC10-augmented surfactant therapy in a lamb model of infant respiratory distress syndrome: physiological, inflammatory, and kinetic profiles.J Appl Physiol, 2005, 99(6):2204-2211.
[31]Calvo FQ, Fillet M, de Seny D, et al.Biomarker discovery in asthma-related inflammation and remodeling.Proteomics, 2009, 9(8):2163-2170.
[32]Shijubo N, Itoh Y, Yamaguchi T, et al.Clara cell protein-positive epithelial cells are reduced in small airways of asthmatics.Am J Respir Crit Care Med, 1999, 160(3):930-933.
[33]Hung CH, Chen LC, Zhang Z, et al.Regulation of TH2 responses by the pulmonary Clara cell secretory 10-kd protein.J Allergy Clin Immunol, 2004, 114(3):664-670.
[34]Wang SZ, Rosenberger CL, Espindola TM, et al.CCSP modulates airway dysfunction and host responses in an Ova-challenged mouse model.Am J Physiol Lung Cell Mol Physiol, 2001, 281(5):L1303-L1311.
[35]Mantile G, Miele L, Cordella-Miele E, et al.Human Clara cell 10-kDa protein is the counterpart of rabbit uteroglobin.J Biol Chem, 1993,268(27):20343-20351.
[36]Andersson O, Nordlund-Möller L, Barnes HJ, et al.Heterologous expression of human uteroglobin/polychlorinated biphenyl-binding protein.Determination of ligand binding parameters and mechanism of phospholipase A2 inhibition in vitro.J Biol Chem, 1994, 269(29):19081-19087.
[37]Mandal AK, Zhang Z, Ray R, et al.Uteroglobin represses allergen-induced inflammatory response by blocking PGD2 receptor-mediated functions.J Exp Med, 2004, 199(10):1317-1330.
[38]Sohn J, Kim TI, Yoon YH, et al.Novel transglutaminase inhibitors reverse the inflammation of allergic conjunctivitis.J Clin Invest, 2003,111(1):121-128.
[39]Luster AD, Tager AM.T-cell trafficking in asthma: lipid mediators grease the way.Nat Rev Immunol, 2004, 4(9):711-724.
[40]Wang XF, Hou YY, Bi D.Expression of serum Clara cell secretory protein 10 in wheezing children under 5 years old.Chin J Contemp,2011, 13(3):199-201.(in Chinese)王秀芳, 侯颖莹, 毕丹.Clara细胞分泌蛋白10在5岁以下喘息儿童外周血中的表达.中国当代儿科杂志, 2011, 13(3):199-201.
[41]Johansson S, Keen C, Ståhl A, et al.Low levels of CC16 in nasal fluid of children with birch pollen-induced rhinitis.Allergy, 2005, 60(5):638-642.
[42]Liu Y, Lu X, Yu HJ, et al.The expression of osteopontin and its association with Clara cell 10 kDa protein in allergic rhinitis.Clin Exp Allergy, 2010, 40(11):1632-1641.
[43]Laing IA, Hermans C, Bernard A, et al.Association between plasma CC16 levels, the A38G polymorphism, and asthma.Am J Respir Crit Care Med, 2000, 161(1):124-127.
[44]Chupp GL, Lee CG, Jarjour N, et al.A chitinase-like protein in the lung and circulation of patients with severe asthma.N Engl J Med,2007, 357(20):2016-2027.
[45]Ober C, Tan Z, Sun Y, et al.Effect of variation in CHI3L1 on serum YKL-40 level, risk of asthma, and lung function.N Engl J Med, 2008,358(16):1682-1691.
[46]Wang H, Long XB, Cao PP, et al.Clara cell 10-kD protein suppresses chitinase 3-like 1 expression associated with eosinophilic chronic rhinosinusitis.Am J Respir Crit Care Med, 2010, 181(9):908-916.
[47]Widegren H, Andersson M, Greiff L.Effects of Clara cell 10 (CC10)protein on symptoms and signs of allergic rhinitis.Ann Allergy Asthma Immunol, 2009, 102(1):51-56.
[48]Liu Z, Lu X, Zhang XH, et al.Clara cell 10-kDa protein expression in chronic rhinosinusitis and its cytokine-driven regulation in sinonasal mucosa.Allergy, 2009, 64(1):149-157.
[49]Zhang F, Xiong ZG, Cao PP, et al.Lack of association of Clara cell 10-kDa protein gene variant with chronic rhinosinusitis in a Chinese Han population.Am J Rhinol, 2008, 22(4):376-380.
[50]Quintar AA, Mukdsi JH, del Valle Bonaterra M, et al.Increased expression of uteroglobin associated with tubal inflammation and ectopic pregnancy.Fertil Steril, 2008, 89(6):1613-1617.
[51]Chowdhury B, Zhang Z, Mukherjee AB.Uteroglobin interacts with the heparin-binding site of fibronectin and prevents fibronectin-IgA complex formation found in IgA-nephropathy.FEBS Lett, 2008,582(5):611-615.
[52]Antico G, Lingen MW, Sassano A, et al.Recombinant human uteroglobin/CC10 inhibits the adhesion and migration of primary human endothelial cells via specific and saturable binding to fibronectin.J Cell Physiol, 2006, 207(2):553-561.
[53]Zhang Z, Kim SJ, Chowdhury B, et al.Interaction of uteroglobin with lipocalin-1 receptor suppresses cancer cell motility and invasion.Gene,2006, 369:66-71.
[54]Yoon JM, Lim JJ, Yoo CG, et al.Adenovirus-uteroglobin suppresses COX-2 expression via inhibition of NF-kappaB activity in lung cancer cells.Lung Cancer, 2005, 48(2):201-209.
[55]Determann RM, Millo JL, Waddy S, et al.Plasma CC16 levels are associated with development of ALI/ARDS in patients with ventilator-associated pneumonia: a retrospective observational study.BMC Pulm Med, 2009, 9:49.
[56]Kropski JA, Fremont RD, Calfee CS, et al.Clara cell protein (CC16),a marker of lung epithelial injury, is decreased in plasma and pulmonary edema fluid from patients with acute lung injury.Chest,2009, 135(6):1440-1447.
[57]Gioldassi XM, Papadimitriou H, Mikraki, et al.Clara cell secretory protein: determination of serum levels by an enzyme immunoassay and its importance as an indicator of bronchial asthma in children.J Pharm Biomed Anal, 2004, 34(4):823-826.
[58]Hasegawa M, Fujimoto M, Hamaguchi Y, et al.Use of serum clara cell 16-kDa (CC16) levels as a potential indicator of active pulmonary fibrosis in systemic sclerosis.J Rheumatol, 2011, 38(5):877-884.
