HIV-1 Vif的克隆、表达与活性检测
2010-01-24覃晓琳刘朝奇刘徽婷汪龚泽
覃晓琳,刘朝奇,刘徽婷,汪龚泽
HIV-1 Vif的克隆、表达与活性检测
覃晓琳,刘朝奇,刘徽婷,汪龚泽
目的 克隆 H IV-1 Vif基因,构建原核表达质粒,进行蛋白的表达及其生物学活性的检测。方法PCR扩增Vif基因,并将其克隆到原核表达载体p ET28a(+)中,构建原核表达质粒p ET28a(+)/Vif,进行双酶切及测序鉴定。获得的阳性质粒转化大肠杆菌BL21(DE3)中进行表达,对表达产物进行SDS-PAGE电泳和Western blot检测分析,利用亲和层析方法纯化Vif蛋白。利用Pull-Dow n方法检测 H IV-1 Vif与SH3(HCK)特异性结合活性。结果通过酶切和测序,结果表明重组质粒p ET28a(+)/Vif构建正确。SDS-PAGE和Western blot结果鉴定了原核表达的Vif重组蛋白大小正确。纯化了蛋白Vif、SH3和 GST。GST pull-dow n试验说明Vif和SH3蛋白具有体外特异性结合活性。结论成功地克隆、表达和纯化了Vif蛋白,Vif与SH3蛋白具有结合活性,为进一步研究针对Vif与SH3结合的药物筛选提供实验依据。
H IV-1 Vif;SH3(HCK);结合
H IV-1Vif基因是 H IV病毒编码的6个辅助蛋白或调控蛋白(accesso ry protein)之一,是一个非常保守的病毒基因,广泛存在于反转录病毒科中所有的灵长类慢病毒及非灵长类慢病毒中(除马传染性贫血病毒)。含有192个氨基酸,分子量约为23kD,在产生感染性病毒颗粒的过程中起着至关重要的作用。因此,人们根据这个特点将其命名为病毒感染因子(Viral infectivity facto r,Vif)〔1-4〕。Src 酪氨酸家族成员 Hck和Fyn酪氨酸激酶可抑制 H IV-1的复制,但当 H IV-1 Vif与 Hck和 Fyn激酶的 SH3结构域直接作用以后,不仅可以抑制激酶活性,同时可以促进病毒的复制〔5-7〕。可见Vif与 SH3结构域的结合在Vif发挥促进病毒感染功能中发挥了非常重要的作用。
本研究构建了 H IV-1Vif的原核表达载体,并纯化了Vif蛋白,体外检测了原核表达的Vif蛋白与SH 3(HCK)蛋白的特异性结合活性,为进一步研究针对Vif与SH3的靶向治疗药物的筛选奠定实验基础。
1 材料与方法
1.1 材料 菌株 E.coli DH5α、E.coli BL21(DE3)、带6x His标签的质粒p ET28a(+)、带谷胱甘肽S-转移酶(GST)标签的质粒p GEX-4T-1和重组质粒p GEX-4T-1/SH3由本研究所保存。限制性核酸内切酶、Taq DNA聚合酶、T4 DNA连接酶、dN TPs、PCR试剂盒由 Fermentas公司提供,DNA纯化试剂盒购自Omega公司。兔源 GST多克隆抗体购自 eBioscience公司,鼠源 His单克隆抗体、HRP标记的羊抗鼠 IgG、HRP标记的羊抗兔 IgG均购自北京中杉金桥生物技术有限公司,其它化学试剂由 Sigma等公司提供。N TA树脂、Glutathione Sepharose 4B、GSTrap FF和蛋白纯化仪KTAp rime购自 GE Healthcare Bioscience公司;PCR引物合成及DNA测序均由上海生工公司完成。
1.2 方法
1.2.1 H IV-1 Vif原核表达载体的构建 以 H IV-1HBX2为模板,PCR扩增Vif基因序列。Vif引物(5′端分别引入限制性内切酶 Bam H I和 Eco RI酶切位点),上游引物 5′-GCGGA TCC A TGGAAAACAGA TGGCA G-3′(Bam H I), 下 游 引 物 5′-GCAGAATTC CTAGTGTCCATTCATTGTGTG-3′
(Eco RI)。PCR反应体系体积为50μL,以2μL的第1链cDNA为模板,PCR扩增条件为:94℃预变性2 m in后,94℃变性30 s,55℃退火30 s,72℃延伸45 s,30个循环后,再于72℃延伸10 min结束扩增,PCR产物用纯化试剂盒进行纯化。用Bam H I和 Eco RI双酶切质粒p ET28a(+)和纯化后的PCR产物,将 PCR产物与质粒载体p ET28a(+)混合,加入 T4连接酶于16 ℃连接过夜,构建含有 Vif基因的重组质粒 p ET28a(+)/Vif,用CaCl2法转化宿主菌 E.coli DH5α。进行双酶切鉴定并送测序鉴定阳性克隆。
1.2.2 Vif蛋白的原核表达与纯化 将构建正确的重组质粒p ET28a(+)/Vif转化宿主菌BL 21(DE3),挑单克隆接种于含卡那霉素的LB液体培养基,37℃振荡培养至A600nm≈0.6~0.8时,加入IPTG(1.0 mmol/L)诱导表达4 h。离心收集菌体,菌体重悬于Buffer A中(0.5mo l/L NaCl,20 mmol/L Tris-HCl,p H 8.0),在冰浴中以超声波破碎菌体,离心后收集沉淀,8mol/L尿素溶解沉淀,将N TA树脂装入层析柱,通过Vif蛋白上的 His标签与镍结合的特点进行亲和层析。层析用10倍N TA体积的N TA-washing Buffer洗涤。将样品加到N TA层析柱中,流速控制在15 m L/h左右,收集穿透部分,然后分别用5倍N TA体积的N TA-p H8.0,N TA-p H6.8,N TA-p H5.6,N TA-p H4.5 Buffer洗脱,收集洗脱液,透析方法进行复性,储存于-40℃。
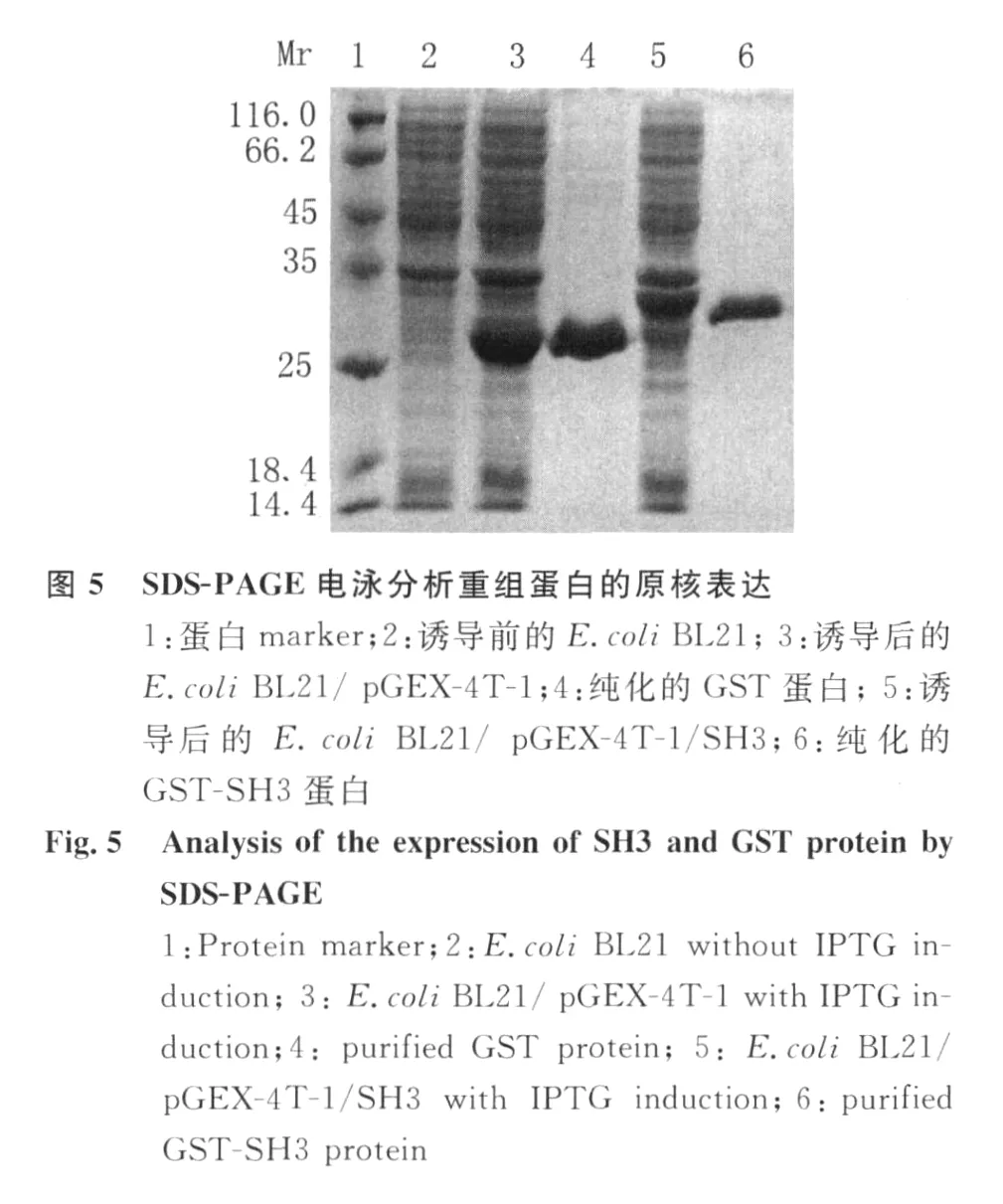
1.2.3 SH3和 GST蛋白的原核表达与纯化 将本实验室保存的质粒p GEX-4T-1和重组质粒p GEX-4T-1/SH3分别转化宿主菌BL 21(DE3),次日各挑单克隆接种于含氨苄青霉素的LB液体培养基,37℃振荡培养至 A600nm≈0.6~0.8时,加入IPTG(1.0 mmol/L)诱导表达4 h。分别离心收集菌体,菌体用PBS洗涤1次,用适量 PBS重悬沉淀,超声波裂解,应用 GSTrap FF亲和层析柱在ÄKTAp rime蛋白纯化仪纯化 GST和 SH3蛋白,SDS-PAGE和Western blot鉴定。
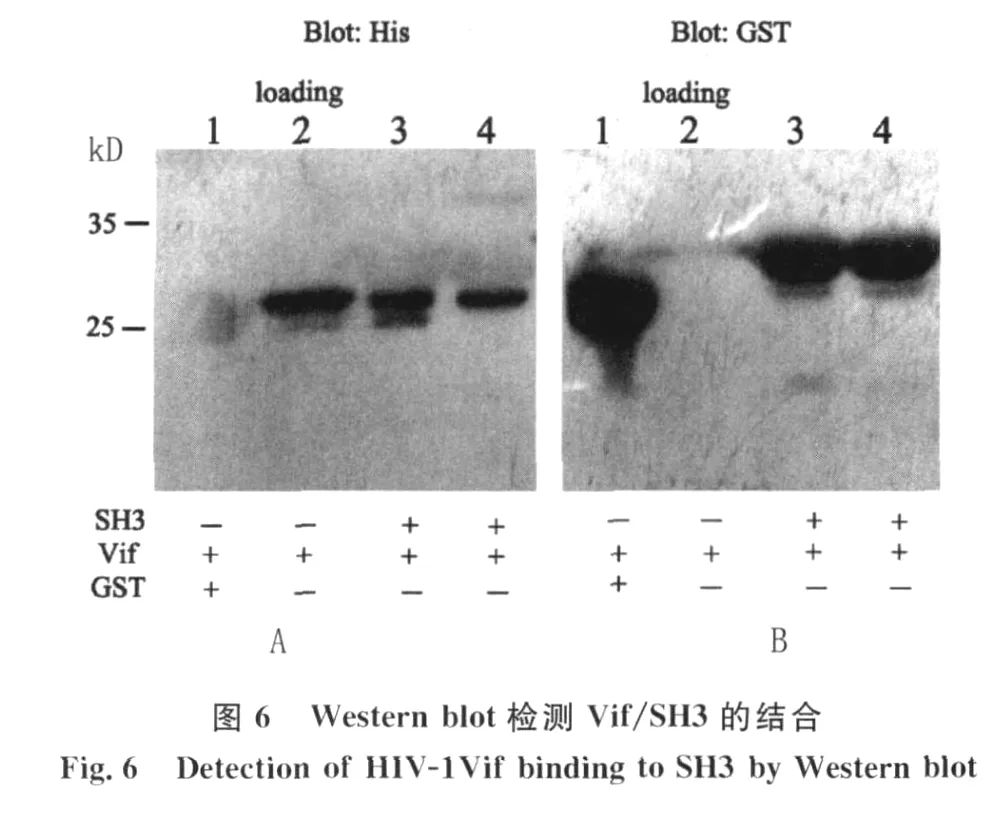
1.3 GST pull-dow n检测V if/SH3的结合活性取适量 Glutathione Sepharose 4B beads,分别加入纯化的 GST蛋白或SH3蛋白,混匀,室温缓慢摇动作用1 h,使 GST标签蛋白与beads特异性结合,PBS洗涤3次。在各管中加入等量纯化的V if蛋白,室温缓慢摇动1 h,用PBS洗涤3次,将未结合的V if蛋白洗涤。加入1×加样缓冲液,煮沸5 min,取上清按常规方法进行12%SDS-PAGE,同时,以直接上样纯化Vif或者 GST蛋白作为阳性对照。产物电转移至PVDF膜,5 g/L脱脂奶粉封闭2 h,加入 His抗体(1∶3 000),室温作用 1 h,用TBST洗膜3次后加入 HRP标记的羊抗鼠 IgG(1∶5 000),37℃孵育1 h,TBST洗膜3次后 ECL显色。Stripping缓冲液于50℃处理上述 PVDF膜30 min,TBST洗涤3次,再以5 g/L脱脂奶粉室温封闭2 h后,加入1∶2 000稀释的GST兔源多克隆抗体,室温孵育1h,TBST洗膜3次后加入 HRP标记的羊抗兔IgG(1∶3 000),37℃孵育1 h,TBST洗膜3次后ECL显色。
2 结 果
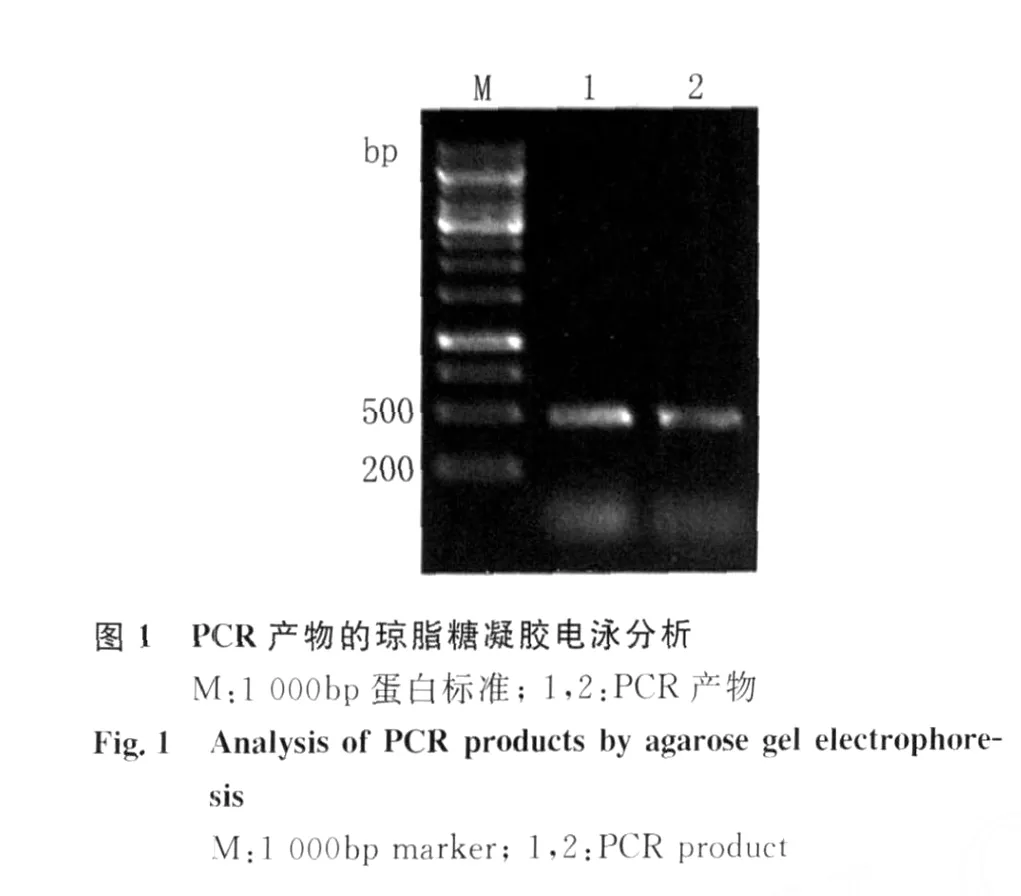
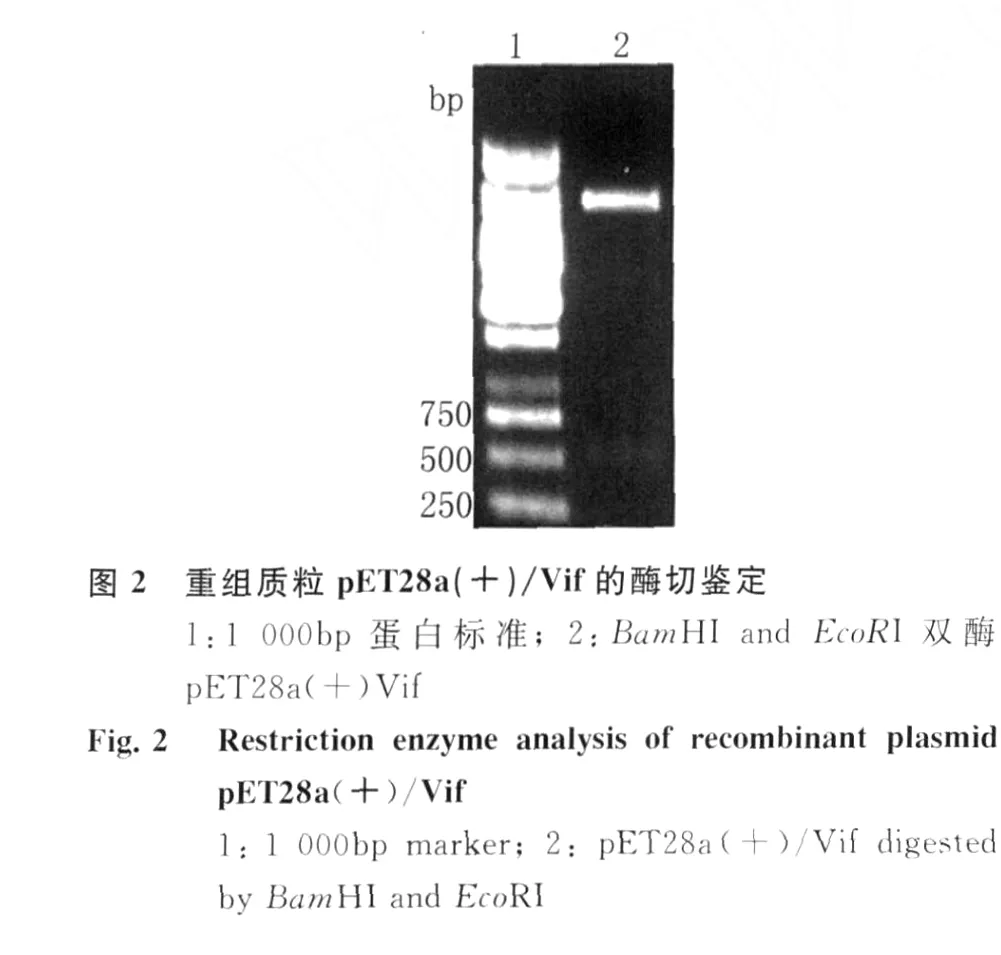
2.1 H IV-1 Vif重组质粒的构建 以 H IV-1 HBX2质粒为模板,经PCR扩增 H IV-1 Vif基因片段,扩增产物经10 g/L琼脂糖凝胶电泳,可见大小约为580 bp的目的条带见图1。将此PCR产物连接到p ET28a(+)质粒中,经过酶切、连接和转化,获得重组质粒 p ET28a(+)/Vif,经 Bam H I和Eco RI双酶切鉴定,获得的插入片段大小与预期结果一致,10 g/L琼脂糖凝胶电泳结果见图2。DNA测序结果显示Vif编码区序列读码框正确,表明重组质粒p ET28a(+)/Vif构建成功。
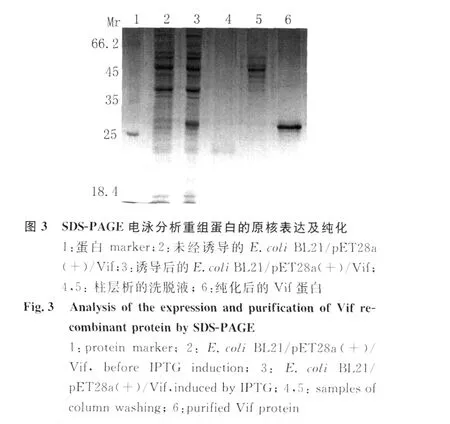
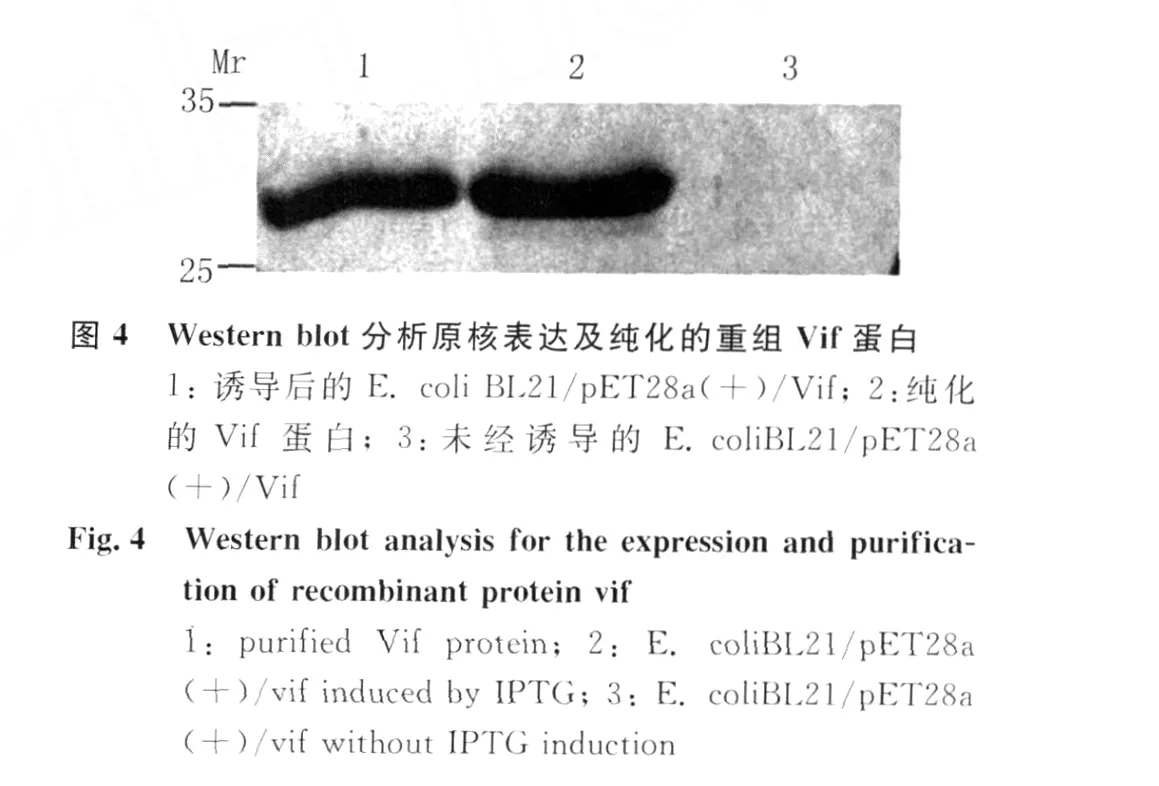
2.2 V if蛋白的原核表达与纯化 用1.0mmol/L IPTG对转化p ET28a(+)/Vif质粒的细菌进行诱导表达,SDS-PAGE电泳显示诱导后的细菌中有一新生的蛋白带,表观分子量与Vif蛋白预期结果相符。大量诱导目的蛋白表达,经尿素溶解包涵体,N TA柱层析得到纯化的V if蛋白见图3。电泳分离的蛋白转移到 PVDF膜与 His单克隆抗体结合后,M r约30kD处可检测到Vif蛋白的 Western blot显色带见图 4,表明 p ET28a(+)/Vif表达正确。
2.3 SH3和 GST蛋白的原核表达与纯化 用1.0 mmol/L IPTG对转化质粒p GEX-4T-1和p GEX-4T-1/SH 3的细菌进行诱导表达 4~6h,SDSPA GE电泳显示诱导后的细菌中,分别有一条大小约为26 kD的 GST蛋白和大小约32 kD的重组GST-SH3蛋白带,表观分子量与预期结果相符。大量诱导目的蛋白表达,经 SDS-PAGE电泳,过柱获得纯化的 GST-SH3和 GST蛋白,见图5。
2.4 GST pull-dow n检测Vif/SH3的结合活性GST蛋白能与 Glutathione Sepharose 4B beads特异性结合,形成 GST-beads和 GST-SH3 beads,加入V if蛋白(含 His标签),通过离心的方式可以使GST-beads沉淀,由于Vif/SH3结合从而使Vif也滞留于沉淀中。图6 A是以 His单抗为一抗的Western blot结果,3#和4#可检测到一条相对分子质量约30kD的Vif蛋白阳性带,而1#的 GST未见条带为阴性,2#为直接上样的纯化Vif蛋白作为阳性对照。为了验证 Glutathione Sepharose 4B beads与 GST-SH3或者 GST特异性结合,上述PVDF膜经stripping缓冲液处理后,将 His抗体洗涤,再以 GST抗体为一抗进行 Western blot检测,图6B结果显示1#的 GST(26kD)和 3#、4#的GST-SH3(32kD)条带位置正确,而直接上样的Vif蛋白孔(2#)为阴性。结果证实原核表达的Vif能与 HCK的SH3结构域特异性结合。
3 讨 论
H IV-1的Vif蛋白可抑制宿主细胞蛋白APOBEC3G介导的抗病毒途径,从而影响 H IV-1病毒颗粒的包装,引起病毒的持续感染〔9〕。有研究发现,细胞内 Fyn和 Hck激酶具有调节A POBEC3G的功能,而Vif蛋白可直接与细胞内 Fyn和 Hck的SH3结构域结合,从而降低激酶的催化活性〔6-8〕,进而降低APOBEC3G的抗病毒作用。因此 H IV-1 Vif蛋白与 Hck SH3结合成为抗病毒的潜在靶点,而建立有效的高通量分子筛选模型用于靶向分子的药物筛选具有非常重要的意义。
本实验成功构建了原核重组质粒p ET28a(+)/Vif,表达并纯化了Vif蛋白。应用亲和层析方法可以有效的纯化 His-Vif蛋白及 GST-SH3蛋白。应用经典的 GST pull-dow n实验,建立了原核表达的Vif蛋白与SH3蛋白特异性结合的分子模型,为研究针对Vif与SH3结合的靶向药物的筛选提供了实验基础〔10-12〕。
〔1〕Walker RC Jr,Khan MA,Kao S,et al.Identification of dominant negative human immunodeficiency virus type 1 Vif mutants that interfere with the functional inactivation of APOBEC3G by virus-encoded Vif〔J〕.J Virol,2010,84(10):5201-5211.
〔2〕Sheehy AM,Gaddis NC,Choi JD,et al.Isolation of a human gene that inhibits H IV-1 infection and is supp ressed by the viral Vif protein〔J〕.Nature,2002,418(6898):646-650.
〔3〕Niew iadom ska AM,Yu XF.Host restriction of H IV-1 by APOBEC3 and viral evasion through Vif〔J〕.Curr Top Microbiol Immunol,2009,339:1-25.
〔4〕Yamashita T,Nomaguchi M,Miyake A,et al.Status of APOBEC3G/F in cells and p rogeny virions modulated by Vif determines H IV-1 infectivity〔J〕.Microbes Infect,2010,12(2):166-171.
〔5〕Guiet R,Poincloux R,Castandet J,et al.Hematopoietic cell kinase(Hck)isoforms and phagocyte duties-from signaling and actin reorganization to migration and phagocytosis〔J〕.Eur J Cell Biol,2008,87(8-9):527-542.
〔6〕Douaisi M,Dussart S,Courcoul M,et al.The tyrosine kinases Fyn and Hck favo r the recruitment of tyrosine-phospho rylated APOBEC3G into vif-defective H IV-1 particles〔J〕.Biochemical and Biophysical Research Communications,2005,(329):917-924.
〔7〕Hassaine G,Courcoul M,Bessou G,et al.The tyrosine kinase Hck is an inhibitor of H IV-1 replication counteracted by the viral vif protein〔J〕.JBiol Chem,2001,(276):16885-16893.
〔8〕Dang Y,Davis RW,York IA,et al.Identification of 81LGxGxx IxW 89 and 171EDRW 174 domains from human immunodeficiency virus type 1 Vif that regulate APOBEC3G and APOBEC3F neutralizing activity〔J〕.J Virol,2010,84(11):5741-5750.
〔9〕Iwatani Y,Chan DS,Liu L,et al.H IV-1 Vif-mediated ubiquitination/degradation of APOBEC3G involves four critical lysine residues in its C-term inal domain〔J〕.Proc Natl Acad Sci U SA,2009,106(46):19539-19544.
〔10〕MezeiM,Minarovits J.Reversalof HIV drug resistance and novel strategies to curb HIV infection:the viral in fectivity factor Vif as a target and tool of therapy〔J〕.Curr Drug Targets,2006,7(7):881-885.
〔11〕Nowotny B,Schneider T,Pradel G,et al.Inducible APOBEC3G-Vif double stable cell line as a high-throughput screening platform to identify antiviral compounds〔J〕.Antimicrob A-gents Chemother,2010,54(1):78-87.
〔12〕Emert-Sedlak L,Kodama T,Lerner EC,et al.Chemical library screens targeting an H IV-1 accessory facto r/host cell kinase complex identify novel antiretroviral compounds〔J〕.ACSChem Biol,2009,4(11):939-947.
Clon ing and expressing of HIV-1 Vif gene in prokaryotic cells,and evaluation of its bioactivity
Q IN Xiao-lin,L IU Chao-qi,L IU Hui-ting,WANG Gong-ze
(Institute of Molecular Biology,Three Gorges University,Yichang 443002,China)
In order to clone H IV-1 Vif gene,construct prokaryotic expression plasmid and detect itsexpression and biological activity,H IV-1 Vif gene wasobtained by PCR and cloned into the vector p ET28a(+)to construct p rokaryotic expression plasmid p ET28a(+)/Vif.The positive plasmid was identified by double enzyme digestion and sequencing.Positive plasmid was transformed to E.coli BL 21(DE3)and the expressed product was identified by SDS-PAGE electropho resis and western blot analysis.Vif protein was expressed and purified,and its biological activity was assayed.Results indicated that the plasmid p ET28a(+)/Vif was correctly constructed.Through transformation of p ET28a(+)/Vif into E.coli BL21(DE3),the Vif protein was expressed and purified.The renatured protein showed that Vif could bind into SH3(HCK)domain in the pulldown assay.It’s concluded that the successful cloning,expression and purification of Vif protein provide the basis for further studieson the functions of the protein.
H IV-1 Vif;SH3(HCK);binding
R392.11
A
1002-2694(2010)12-1141-04
刘朝奇,Email:zqliu@ctgu.edu.cn,qinaiaolin200810@163.com
三峡大学分子生物学研究所,宜昌 443002
2010-07-10;
2010-09-23
