1株分离自猩红热患儿的耐多药嵴链球菌全基因组测序及比较基因组分析
2021-08-06黄银燕贾庆军吴亦斐程庆林李清春
黄银燕,赵 刚,贾庆军,吴亦斐,程庆林,王 乐,陆 敏,李清春,谢 立
猩红热(Scarlet Fever)是由能产生红疹毒素的A组β溶血性链球菌(beta-haemolytic group A streptococcus isolates, GAS),也称化脓链球菌(S.pyogenes)感染所引起的一种急性呼吸道传染病,是我国法定的乙类传染病之一[1]。有研究表明嵴链球菌一般分离于健康或者龋齿患者的牙菌斑,有利于延缓龋齿的发生[2],而从猩红热患者咽拭子分离到嵴链球菌报道较少[3]。本课题组前期研究从确诊的猩红热患儿咽拭子分离到一株嵴链球菌S22,并且未分离到化脓链球菌[3]。本文对S22进行全基因组测序分析,并将其与NCBI基因组数据库中的嵴链球菌和部分具有代表性的链球菌基因组进行比较基因组学研究,从基因组的角度探讨嵴链球的致病机理、耐药机制及其与其他链球菌在基因水平的差异和进化关系,探索嵴链球菌与猩红热之间的关系,为临床用药和疾病防控等提供一定参考依据。
1 材料与方法
1.1 菌株来源 菌株S22分离自杭州市儿童医院猩红热患儿咽拭子[3]。细菌药物敏感实验采用纸片扩散法测定S22对红霉素、罗红霉素、阿奇霉素、泰利菌素、螺旋霉素、林可霉素、链阳霉素B、四环素、链霉素、庆大霉素、卡那霉素、西索米星、妥布霉素、阿米卡星、奈替米星和杆菌肽。将单菌落接种于含 5%牛血清的脑心浸液培养管中,37 ℃孵育过夜。调整菌液浊度为0.5 麦氏比浊浓度,作1∶1 000 倍稀释。以无菌棉拭子蘸取稀释液均匀涂布在MH琼脂培养基表面,用无菌镊子贴上各药敏纸片并轻压,两纸片之间距离以 2~3 cm 为宜。平皿倒置于 37 ℃恒温培养 18~24 h,测定抑菌圈直径大小,参照《纸片法药敏试验抑菌环直径判断标准(mm)》(CLSI)中链球菌抑菌环直径标准判定菌株对药物的敏感性。
文中用于比较基因组学分析的28株链球菌基因组信息从NCBI基因组数据库下载,详细信息见表1、表2。
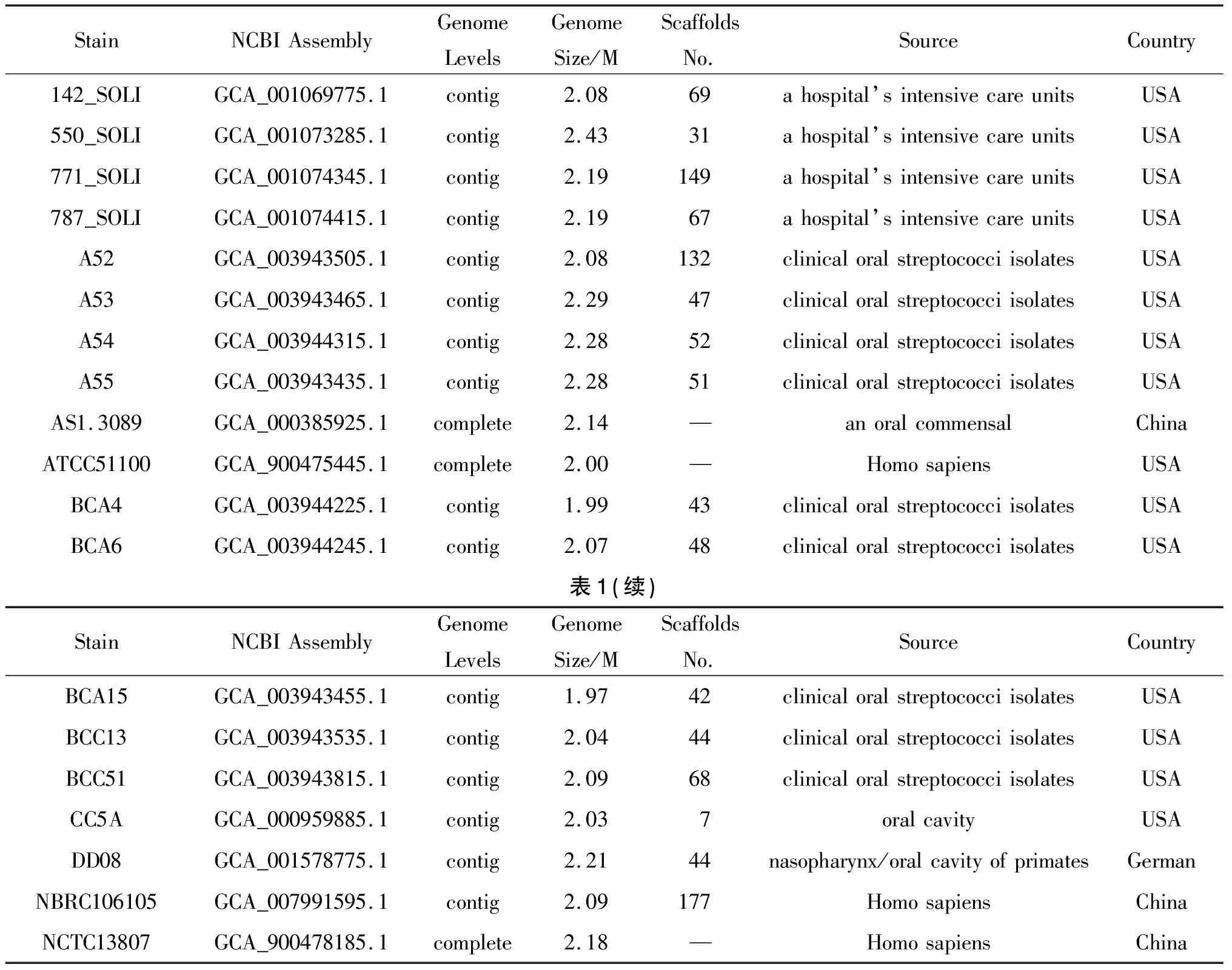
表1 NCBI基因组数据库中19株S.cristatus基因组信息
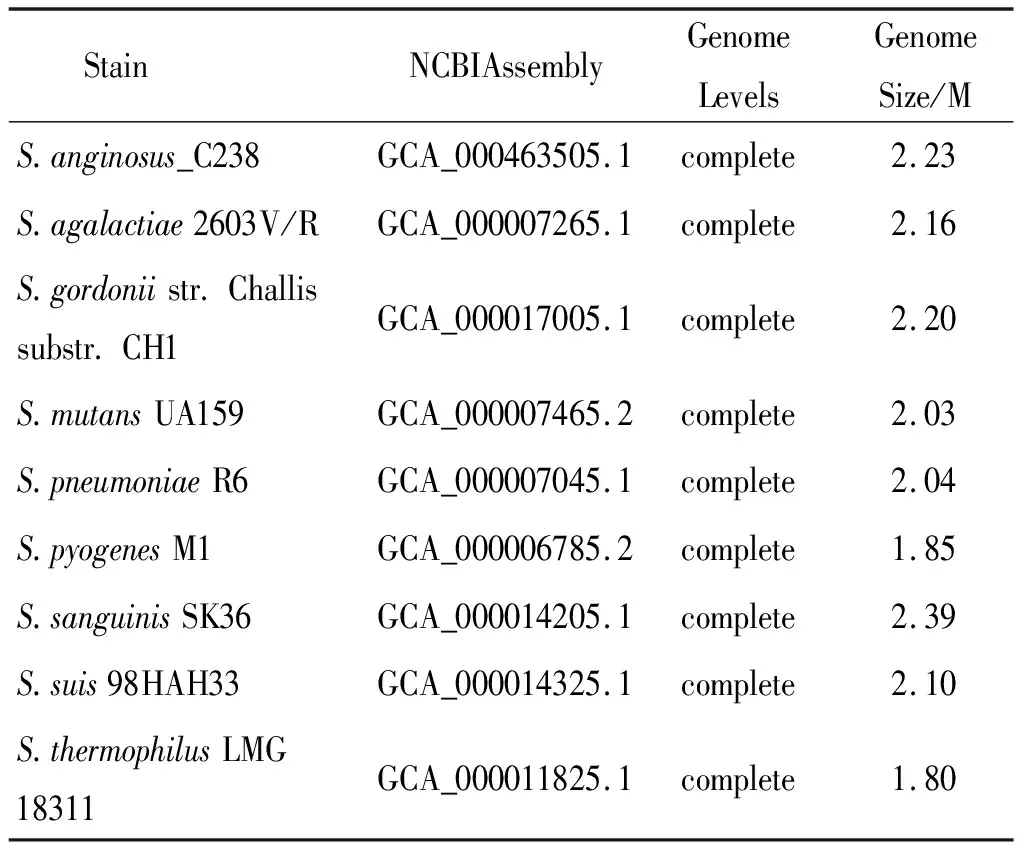
表2 相关链球菌基因组信息
1.2 DNA提取、基因组测序、注释和组装 总DNA的抽提使用DNA抽提试剂盒(Blood & Cell Culture DNA Mini Kit(Qiagen)),参照操作步骤完成。全基因组测序由武汉菲沙基因信息有限公司完成,采用Illumina Hiseq平台测序,使用 SMRT LINK 5.1 软件进行数据处理。基因组组装用SOAPdenovoV2.04软件完成,用多种参数反复调试,选取最佳组装结果。用软件Gapcloser v1.12填补组装后的缺口。组装后的基因组提交NCBI注释,S22的NCBI序列号为VIBR01000000。
1.3 基因组基本特征、毒力基因、耐药基因分析 CDS预测采用Prokka1.12(https://github.com/tseemann/prokka)软件进行;tRNA和rRNA预测分别采用tRNAscan-SE-1.3.1和RNAmmer-1.2;crispr 预测采用CRISPRfinder(http://crispr.i2bc.paris-saclay.fr/Server/)。S22毒力基因预测使用BLAST软件将CDS翻译的蛋白与VFDB[4](http://www.mgc.ac.cn/VFs/)中链球菌毒力基因进行比对,参数e-value:1-e5,相似度50%,比对区域覆盖比对结果为两条序列中至少一条的50%。20株S.cristatus的毒力基因分析是将注释基因组提交到VFDB的VFanalyzer[4]生成。其余9株链球菌毒力基因直接从VFDB的链球菌属下载。耐药基因预测使用Antibiotic Resistance Genes Database(https://ardb.cbcb.umd.edu/)数据库及其自带脚本进行分析。
1.4 比较基因组分析 平均核苷酸相似度(ANI)采用软件Jspecies分析[5]。链球菌属基于全基因组的SNPs进化树采用软件kSNP3[6]分析(参数-k 29,-min_frac 0.5)。
2 结 果
2.1 S22的抗生素耐药状况 纸片扩散法结果显示S22对红霉素、罗红霉素、阿奇霉素、泰利菌素、螺旋霉素、林可霉素、链阳霉素B、四环素、杆菌肽耐药,对链霉素、庆大霉素、卡那霉素、西索米星、妥布霉素、阿米卡星、奈替米星敏感。
2.2 基因组基本信息 菌株S22基因组全长2.04184 Mb,平均GC含量42.7%,共组装成30个Scaffold。Prokka1.12 预测S22有2 020个基因,39个编码20种氨基酸的tRNA,1 980个潜在的CDS,1个rRNA操纵子,22个基因岛,1个crispr。用于比较基因组学分析的28株链球菌包括19株嵴链球菌(表1)和9株经典的致病性链球菌/缓症链球菌(表2),9株菌均有全基因组完成图,并且是研究链球菌毒力因子的参考菌株[4]。19株嵴链球菌有3株分离自中国,1株分离自德国,其余均分离自美国[7-8]。S22和其他28株链球菌16S rRNA基因系统进化树如图1所示。除S22以外,分离自中国的其他3株菌都在一个分支上,与S22 16S rRNA基因亲缘关系最近的是嵴链球菌CC5A。与嵴链球菌550_SOLI 16S rRNA基因亲缘关系最近的是血链球菌SK36,与嵴链球菌771_SOLI和嵴链球菌787_SOLI 16S rRNA基因亲缘关系最近的是格氏链球菌CH1。

图1 N-J法构建的基于16S rRNA基因的系统进化树
2.3 S22的耐药基因和耐药转座子分布 在S22中存在3个耐药基因,分别为ermB、tetM、bacA(表3)。其中ermB、tetM,位于同一个Scaffold上的相近区域,该区域与Enterococcusfaecalis质粒pCF10转座子Tn925(NC_006827)大部分同源(图2)。另外,S22的转座子在Tn925序列的基础上又多插入5个基因,包括ISL3 family transposase,ermB,23SrRNA methyltransferase attenuation leader peptide和2个未知功能蛋白。
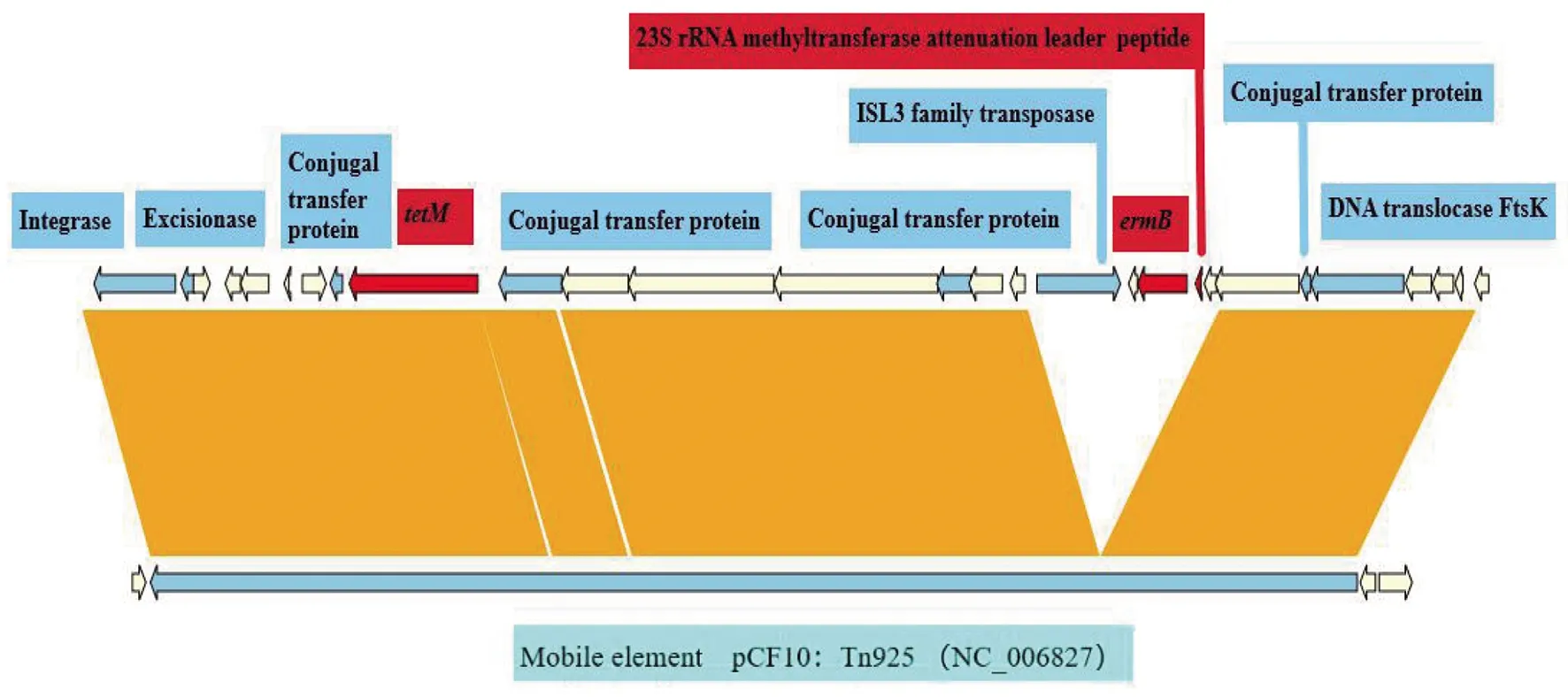
图2 S22基因组耐药转座子区域
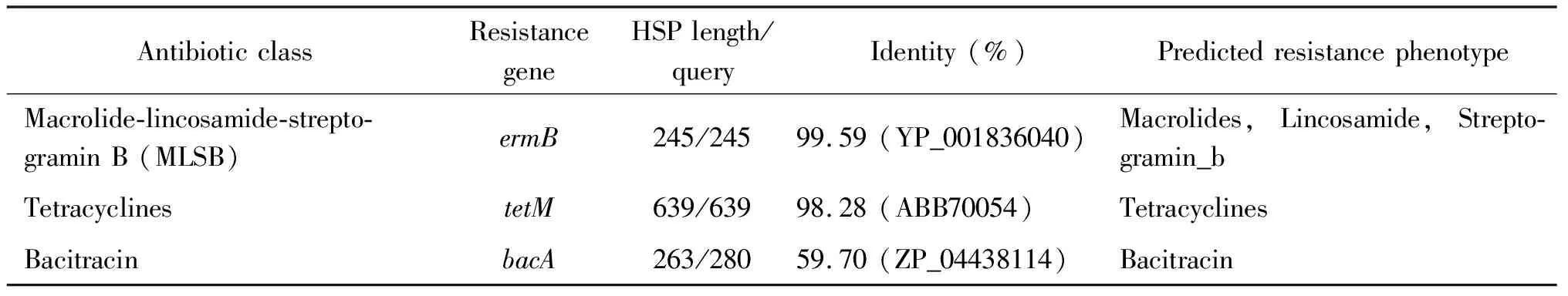
表3 S22的耐药基因
2.4 链球菌属毒力基因分析 链球菌的毒力基因分为黏附素(Adherence),酶类(Enzyme),免疫逃逸(Immune evasion),锰吸收(Manganese uptake),蛋白酶(Protease),超抗原(Superantigen),抗吞噬(Antiphagocytosis)7大类,45个毒力基因(簇)(图3),其中S22含有33个毒力基因,包括黏附素基因5个,酶类基因1个,免疫逃逸(荚膜)基因21个,锰吸收基因1个,蛋白酶基因4个,抗吞噬蛋白基因1个(图3)。除550_SOLI,771_SOLI,787_SOLI以外,其余17株嵴链球菌毒力基因分布大致相同。S22与化脓链球菌M1毒力基因差异很大,M1中与猩红热致病相关的 M protein、Immunoreactive antigen、Superantigen、Hyaluronidase 等毒力基因在S22中均未发现。相比其他嵴链球菌,550_SOLI与SK36,771_SOLI、787_SOLI与CH1的毒力基因更相似。
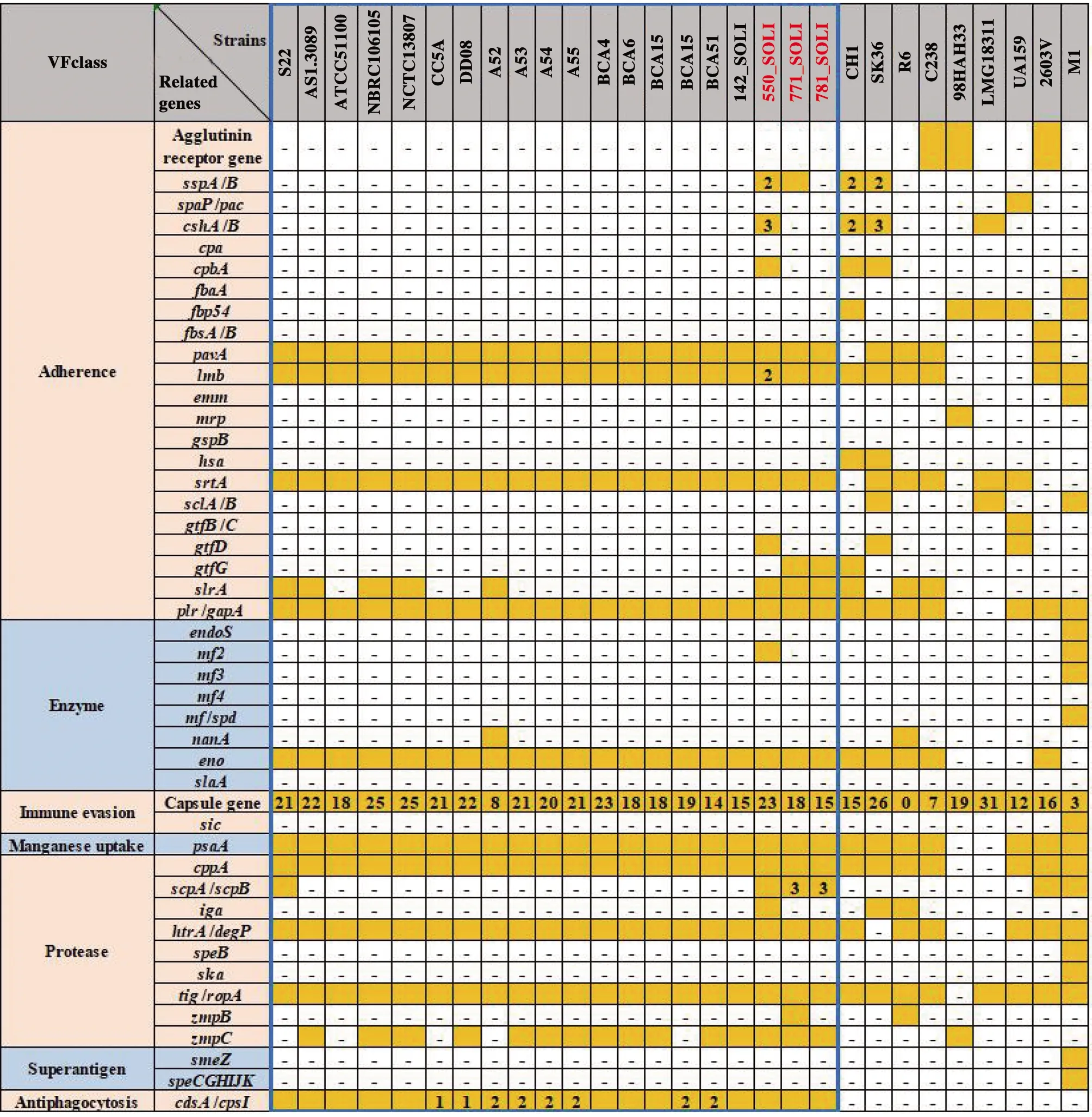
注:蓝色方框中是20株S.cristatus
2.5 嵴链球菌ANI分析 20株嵴链球菌的ANI如表4所示。研究表明属于同一种的两株菌之间ANI大于95%[9]。所有嵴链球菌中未发现与S22 ANI大于95%的菌株。与S22 ANI值最大的是NCTC13807(94.39%),其次是NBRC106105(94.37%)和AS1.3089(94.36%),3株菌均分离自中国且3株菌间的ANI大于99%。A53、A54、A55 3株菌的ANI也大于99%。550_SOLI、771_SOLI、787_SOLI与其他嵴链球菌的ANI均小于90%。771_SOLI和787_SOLI间的ANI是95.81%,2株菌与格氏链球菌CH1的ANI分别是97.75%和95.24%,550_SOLI与血链球菌SK36的ANI是95.33%。
2.6 链球菌属全基因组SNPs进化树 19株已测序嵴链球菌与9株毒力基因参考链球菌的全基因组SNPs进化树如图4所示。整个进化树分为3大分支:S22与分离自中国的NCTC13807、NBRC106105、AS1.3089在一个分支上;771_SOLI、787_SOLI、550_SOLI与9株毒力基因参考链球菌在一个分支上;其余13株嵴链球菌在一个分支上。其中,771_SOLI、787_SOLI与CH1在一个小分支上,550_SOLI与SK36在一个小分支上。
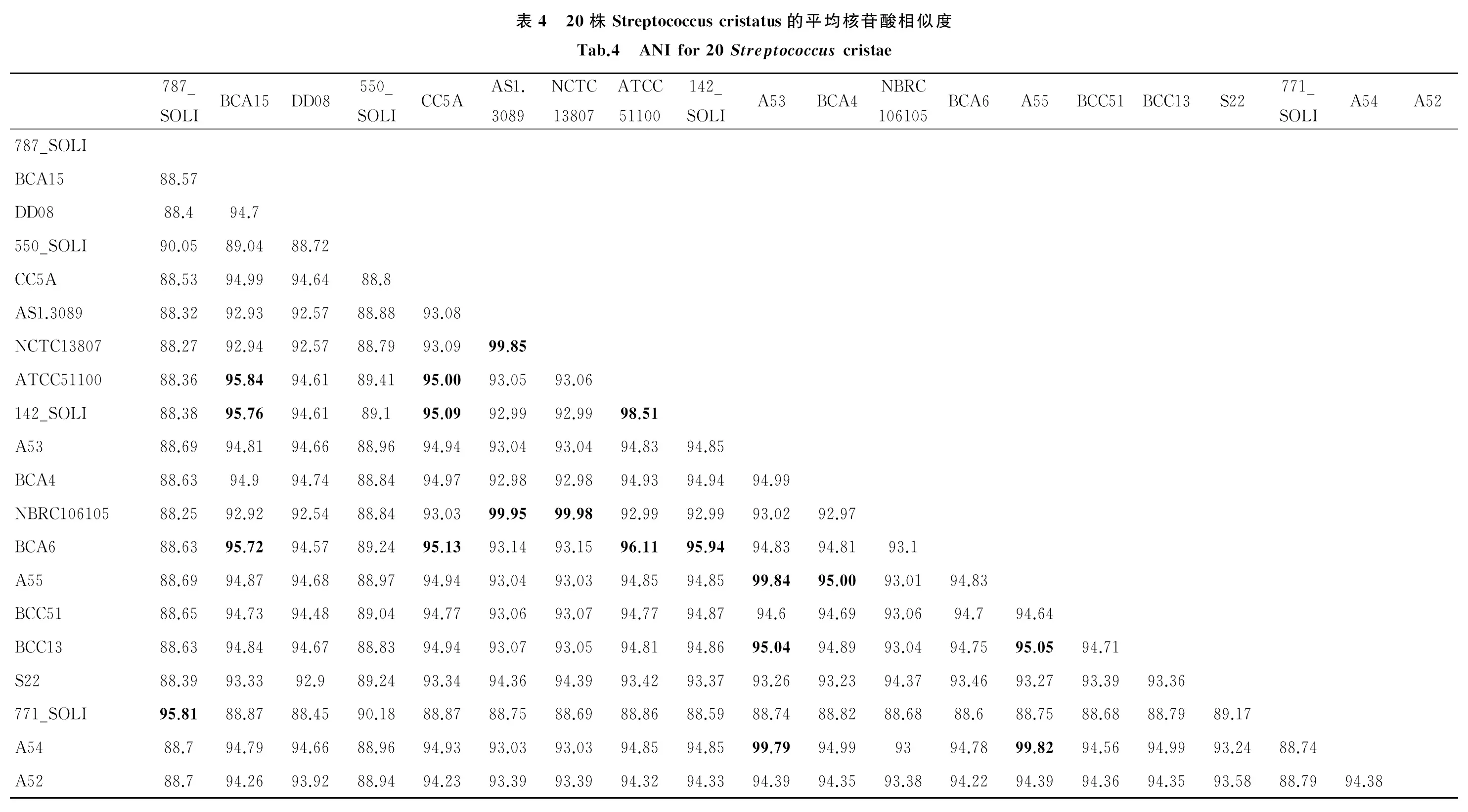

图4 N-J法构建的基于全基因组SNPs的系统进化树
3 讨 论
本文是首次系统地研究了嵴链球属的基因组特征、毒力基因、耐药基因和进化关系,也是首次从猩红热患儿咽拭子分离嵴链球菌进行基因组测序。综上数据,在嵴链球菌中,16S rRNA基因进化树亲缘关系越近,全基因组SNPs进化树亲缘关系也越近,ANI值也越高,毒力基因数量和分布也越相似。因此从16S rRNA基因进化树、全基因组SNP进化树、ANI结果、毒力基因分布可以判断S22属于链球菌属嵴链球菌种。S22与引起猩红热的化脓链球菌进化关系远,毒力基因分布差异大,因此从基因组角度没有直接证据显示S22与猩红热的关系。
S22的33个毒力基因,除了与致病有关的荚膜基因以外,基因plr/gapA[10]、lmb[11]、psaA[12]、srtA[13]、slrA[14]、lmb[15]、eno[16]、pavA[17]与细菌的黏附和定植相关。基因htrA/degP[18]、tig[19]、cppA[20]和scpA/scpB[21]为侵袭性的酶类基因。因此,条件致病菌S22是存在潜在致病能力的,图3中其他的嵴链球菌都是有潜在致病能力的。
嵴链球菌771_SOLI和嵴链球菌787_SOLI虽被命名为嵴链球菌,但16S rRNA基因进化树、全基因组SNPs进化树、ANI数据和毒力基因分布都显示与771_SOLI和787_SOLI亲缘关系最近的是格氏链球菌CH1。因此,771_SOLI和787_SOLI应属格氏链球菌。同理,与550_SOLI亲缘关系最近的是血链球菌SK36,故550_SOLI应属血链球菌。
SNPs进化树上,分离自中国的4株菌在同一个分支上,相互间的ANI值也是最高的。有趣的是与S22 16S rRNA基因亲缘关系最近的是分离自美国的CC5A,并非其他3株分离自中国的菌株。环境影响细菌基因组的进化,中国的4株菌分离自黄种人,而其他的嵴链球菌分离自白种人(1株德国,其余美国),相同生境的菌全基因组亲缘关系越近。SNPs进化树上,S22虽与AS 1.3089、NCTC13807、NBRC 106105在同一个分支上,但相对于其他3株菌的亲缘关系,S22更远,可能与其分离自咽部有关。
S22的耐药基因ermB编码 23S rRNA 甲基化酶,耐药表型为 MLSB,即对大环内酯类、林可霉素和链阳霉素B交叉耐药,TetM基因编码四环素耐药基因,bacA为杆菌肽耐药基因。S22中未发现其他耐药基因,耐药基因和表型完全一致。耐药基因bacA在所有已测序的嵴链球菌中均有发现。耐药基因转座子是在四环素转座子Tn925[22]的基础上又插了1个ermB基因,与转座子Tn6002(AY898750.1)[23]同源性99%。该转座子同时也存在于分离自中国的 AS 1.3089(CP004409.1),NCTC13807(LS483471.1),NBRC 106105(NZ_BJYQ01000021.1)。本研究中的其他菌只有550_SOLI,771_SOLI,ATCC51100含无ermB基因插入的四环素转座子。该转座子比我们在血链球菌S28中发现的耐药基因转座子刚好少1个耐氨基糖苷类抗生素的基因aacA-aphD[24]。
研究显示大环内酯类、头孢菌素和青霉素这3种抗生素在国内儿童医院用量很大[25]。研究人员对2002-2006年5家儿童医院的调查发现,有3家医院使用前3位的抗生素是阿奇霉素、头孢菌素二代、三代[26],大环内酯类抗生素是肺炎支原体感染的首选药物[27]。S22寄居在咽部,若患者使用大环内酯类抗生素后,因S22耐大环内酯类抗生素,有可能成为咽部的优势菌群,甚至因为抗生素的使用,引起菌群失调。S22作为条件致病菌可能引起致病。
目前,从中国分离的4株已测序的嵴链球菌中都含有ermB-tetM类Tn6002转座子,其他已测序的16株嵴链球菌中均未发现该转座子。虽然没有直接证据证明该转座子在中国地区分离的嵴链球菌中的出现与国内抗生素使用的直接关联,但本研究也提示我们:严格控制抗生素的使用,减少抗生素诱导细菌耐药的发生,刻不容缓。
利益冲突:无
