异源细胞质对小麦雄性不育系主要农艺性状、花粉粒核分裂及花药绒毡层降解的影响
2020-07-03周雨馨蒋韫博张改生刘明宇宋瑜龙
罗 茜 周雨馨 马 瑞 蒋韫博 张改生 刘明宇 牛 娜 宋瑜龙
(1 西北农林科技大学农学院/国家杨凌农业生物技术育种中心/国家小麦改良中心杨凌分中心/小麦育种教育部工程研究中心/陕西省作物杂种优势研究与利用重点实验室,陕西 杨凌 712100;2 旱区作物逆境生物学国家重点实验室,陕西 杨凌 712100)
小麦(Tritiaum aestivum)是世界上总产量仅次于玉米的第二大粮食作物,同时也是我国三大主粮之一。我国大部分人口以小麦为主食,使得我国既是世界范围内最大的小麦生产国,也是最大的消费国,小麦高产和稳产对保障我国粮食安全和社会稳定具有非常重要的意义[1]。 近年来,随着城市化建设速度的加快和退耕还林工程的实施,我国耕地面积逐年减少,加之农业主管部门双减(减肥减药)项目的实施,大幅度提高小麦单位面积产量,对保障国内粮食市场稳定和口粮安全显得非常重要[2-3]。 小麦杂种优势利用技术是大幅度提高小麦单产的重要途径之一[4-6],而雄性不育系的选择则是实现小麦杂种优势利用提高单产和品质的关键。 小麦雄性不育系主要有三类:一是遗传型不育系,二是光温敏雄性不育系,三是化学杂交剂诱导的生理型雄性不育系[7]。 光温敏雄性不育系的育性转换易受环境影响[8-9],化学杂交剂制种成本较高,这极大地限制了小麦光温敏雄性不育系和化学诱导的生理型雄性不育系的利用,使得越来越多的研究逐渐转向遗传型不育系,以期实现小麦杂种优势利用技术的大面积推广[10]。
核质互作雄性不育系属遗传型不育系的一类。 近年来,核质互作雄性不育的研究多集中在不育系花药酶学特征分析[11],绒毡层代谢与花粉粒败育的关系[12-15],以及败育植株的生物学特性和细胞学特征分析[16-19]等方面,缺乏一定的系统性和完整性。 截止目前,关于异源细胞质对花粉粒核分裂及绒毡层降解特征影响的研究尚鲜有报道。 因此,本研究以2 套小麦核(偃师9 号细胞核和90-110 细胞核)质(K 型、B型、S 型、V 型和Ven 型细胞质,均来自山羊草属)互作雄性不育系及其保持系(偃师9 号细胞核和90-110细胞核,普通小麦细胞质)为材料,研究异源细胞质对小麦主要农艺性状、花粉粒核分裂及花药绒毡层降解特征的影响,以期进一步揭示小麦核质互作雄性不育机理,为更好地利用异源核质互作雄性不育系提供坚实的理论依据。
1 材料与方法
1.1 试验材料
供试小麦材料为2 套核质互作雄性不育系及其对应的保持系(表1),其中,偃师9 号细胞核属1B/1R类型,90-110 细胞核属非1B/1R 类型,均由西北农林科技大学张改生教授提供。 上述材料种植于西北农林科技大学试验农场,每份材料种植3 行,每行长2 米,单行播15 株,日常管理同大田。

表1 供试材料Table 1 Experiment almaterials in present study
1.2 取样与材料固定
待上述2 套核质互作雄性不育系及其保持系材料发育至单核早期、单核晚期、二核期和三核期时,分别取10 个长势一致的穗子放入冰盒,迅速带回实验室,剥取位于穗中部的1 个小花中的1 枚花药置于干净的载玻片上,滴加1.5%醋酸洋红,用镊子将花药夹碎,于显微镜下确定花粉粒发育时期。 随后摘取该小花中剩余的2 枚花药分别投入装有卡诺氏固定液和FAA(formalin-aceto-alcohol)固定液的1.5 mL 离心管中(每个材料各时期2 种固定液中各固定10 枚花药),并对其进行抽真空处理,最后转至4℃冰箱保存。
1.3 农艺性状调查
待小麦发育至扬花期,分别在每份材料中随机抽样取18 株,进行株高、穗长、穗下节间长和分蘖数等农艺性状的调查。 上述农艺性状调查方法均参考《农作物品种区域试验技术规程 小麦》[20]。
1.4 花粉粒核分裂观察
从卡诺氏固定液中取1 枚花药置于干净的载玻片上,滴加1.5%醋酸洋红染色,并置于光学显微镜下观察,同时选取3 个视野进行拍照。 各材料不同时期(单核早期、单核晚期、二核期和三核期)分别取3 枚花药,进行染色。 用1% KI-I2对各材料三核期的花粉粒进行染色,并于光学显微镜下观察,鉴定败育类型。
1.5 石蜡切片制备与花药绒毡层观察
参照胡丽芳等[21]、豆丽萍等[22]的石蜡制片方法,用FAA 固定4 d 后,将花药转移至70%乙醇,于4℃冰箱中保存,以备制作石蜡切片。 取上述花药分别经梯度酒精脱水、二甲苯透明、石蜡渗透、包埋和修蜡后,用轮式切片机切片,厚度为11 μm,随后展片10 min,置于40℃恒温箱中保存3 d。 待切片粘合牢固后,进行脱蜡、复水和番红-固绿复染,最后封片。
普通光学显微镜下观察不同材料各时期(单核早期、单核晚期、二核期和三核期)3 个花药横截面,每个花药截面选代表性视野拍照4 张,合计12 张,随后用Image-Pro Plus 6.0 软件(https:/ /cloud.tencent.com/developer/news/16116)将所采集的不同材料各时期花药横截面绒毡层细胞面积转化为数值。
1.6 数据分析
使用IBM SPSS Statistics 20 软件对株高、穗长、穗下节间长、分蘖数和绒毡层面积分别进行单因素方差分析(One-way ANOVA)[23],同时采用Duncan 多重比较检验法对数据进行差异显著性分析,P<0.05 表示数据间存在显著差异。 所得数据以平均值±标准误表示。
2 结果与分析
2.1 异源细胞质对小麦农艺性状的影响
S-偃师9 号和Ven-偃师9 号株高显著高于偃师9 号保持系、K-偃师9 号、B-偃师9 号与V-偃师9 号,但S-偃师9 号与Ven-偃师9 号株高间无显著差异,且偃师9 号保持系、K-偃师9 号、B-偃师9 号与V-偃师9 号株高间也无显著差异。 K-90-110 株高显著低于90-110 保持系,S-90-110 株高显著低于90-110保持系、B-90-110 和V-90-110,Ven-90-110 株高显著低于90-110 保持系、K-90-110、B-90-110 和V-90-110,但90-110 保持系、B-90-110 与V-90-110株高间无显著差异,K-90-110、B-90-110 与V-90-110 株高间无显著差异,K-90-110 与S-90-110 以及S-90-110 与Ven-90-110 株高间也无显著差异。 比较同质异核不同材料株高发现,偃师9 号保持系株高显著低于90-110 保持系,同时,K-偃师9 号、B-偃师9 号与V-偃师9 号株高也分别显著低于K-90-110、B-90-110 与V-90-110,但S-偃师9 号和Ven-偃师9号株高分别与S-90-110 和Ven-90-110 间无显著差异(图1-A)。 上述结果表明,小麦核质互作雄性不育系细胞核为1B/1R 类型或非1B/1R 类型时,与保持系比较,B 型和V 型细胞质对小麦株高均无显著影响;而K 型、S 型和Ven 型细胞质对小麦株高的影响与细胞核1B/1R 类型有关,其中S 型和Ven 型细胞质在非1B/1R 类型细胞核存在时可显著降低株高,而在1B/1R 类型细胞核存在时可显著增加株高,而K 型细胞质在非1B/1R 类型细胞核存在时可显著降低株高。在1B/1R 类型细胞核体存在时,对株高无显著影响。
B-偃师9 号穗长显著高于偃师9 号保持系,且S-偃师9 号穗长显著高于偃师9 号保持系、K-偃师9号、V-偃师9 号和Ven-偃师9 号,但偃师9 号保持系、K-偃师9 号、V-偃师9 号和Ven-偃师9 号穗长间无显著差异,K-偃师9 号、B-偃师9 号、V-偃师9 号和Ven-偃师9 号穗长间也无显著差异,B-偃师9 号与S-偃师9 号穗长间同样无显著差异。 K-90-110 和B-90-110 穗长显著高于90-110 保持系和V-90-110,但90-110 保持系、S-90-110、V-90-110 和Ven-90-110 穗长间无显著差异,K-90-110、B-90-110、S-90-110 和Ven-90-110 穗长间也无显著差异。 比较同质异核不同材料穗长发现,S-偃师9 号和V-偃师9 号穗长分别显著高于S-90-110 和V-90-110,但偃师9 号保持系、K-偃师9 号、B-偃师9 号和Ven-偃师9 号穗长分别与90-110 保持系、K-90-110、B-90-110 和Ven-90-110 间无显著差异(图1-B)。 上述结果表明,两种类型细胞核存在时,V 型和Ven 型细胞质对小麦穗长均不存在显著影响,但B 型细胞质均可显著增加小麦穗长。 此外,K 型细胞质在非1B/1R 类型细胞核存在时可显著增加小麦穗长,与之相似,S 型细胞质在1B/1R 类型细胞核存在时同样可显著增加穗长。
不同异源细胞质类型偃师9 号穗下节间长均无显著差异。 S-90-110 和V-90-110 穗下节间长显著低于90-110 保持系、K-90-110 和B-90-110,且Ven-90-110 穗下节间长显著低于90-110 保持系,但90-110 保持系、K-90-110 和B-90-110 穗下节间长无显著差异,K-90-110、B-90-110 和Ven-90-110 穗下节间长无显著差异,S-90-110、V-90-110 和Ven-90-110 穗下节间长也无显著差异。 比较同质异核不同材料穗下节间长发现,相同异源细胞质类型偃师9 号穗下节间长均显著高于相应的90-110(图1-C)。 上述结果表明,K 型和B 型细胞质无论是在1B/1R 类型细胞核存在时还是非1B/1R 类型细胞核存在时,对小麦穗下节间长均无显著影响;而S 型、V 型和Ven 型细胞质在非1B/1R 类型细胞核存在时可显著降低小麦穗下节间长。
B-偃师9 号、S-偃师9 号和V-偃师9 号分蘖数显著高于偃师9 号保持系、K-偃师9 号和Ven-偃师9号,但偃师9 号保持系、K-偃师9 号和Ven-偃师9 号分蘖数间无显著差异,B-偃师9 号、S-偃师9 号和V-偃师9 号分蘖数间也无显著差异。 B-90-110 分蘖数显著高于S-90-110 和V-90-110,但90-110 保持系、K-90-110、S-90-110、V-90-110 和Ven-90-110 分蘖数间无显著差异,且90-110 保持系、K-90-110、B-90-110 和Ven-90-110 分蘖数间也无显著差异。 比较同质异核不同材料分蘖数发现,S-偃师9 号和V-偃师9 号分蘖数分别显著高于S-90-110 和V-90-110,但偃师9 号保持系、K-偃师9 号、B-偃师9 号和Ven-偃师9 号分蘖数分别与90-110 保持系、K-90-110、B-90-110 和Ven-90-110 无显著差异(图1-D)。上述结果表明,无论小麦核质互作雄性不育系细胞核为1B/1R 类型还是非1B/1R 类型,K 型和Ven 型细胞质对小麦分蘖数均无显著影响,而B 型、S 型和V 型细胞质在1B/1R 类型细胞核存在时会显著增加小麦分蘖数。
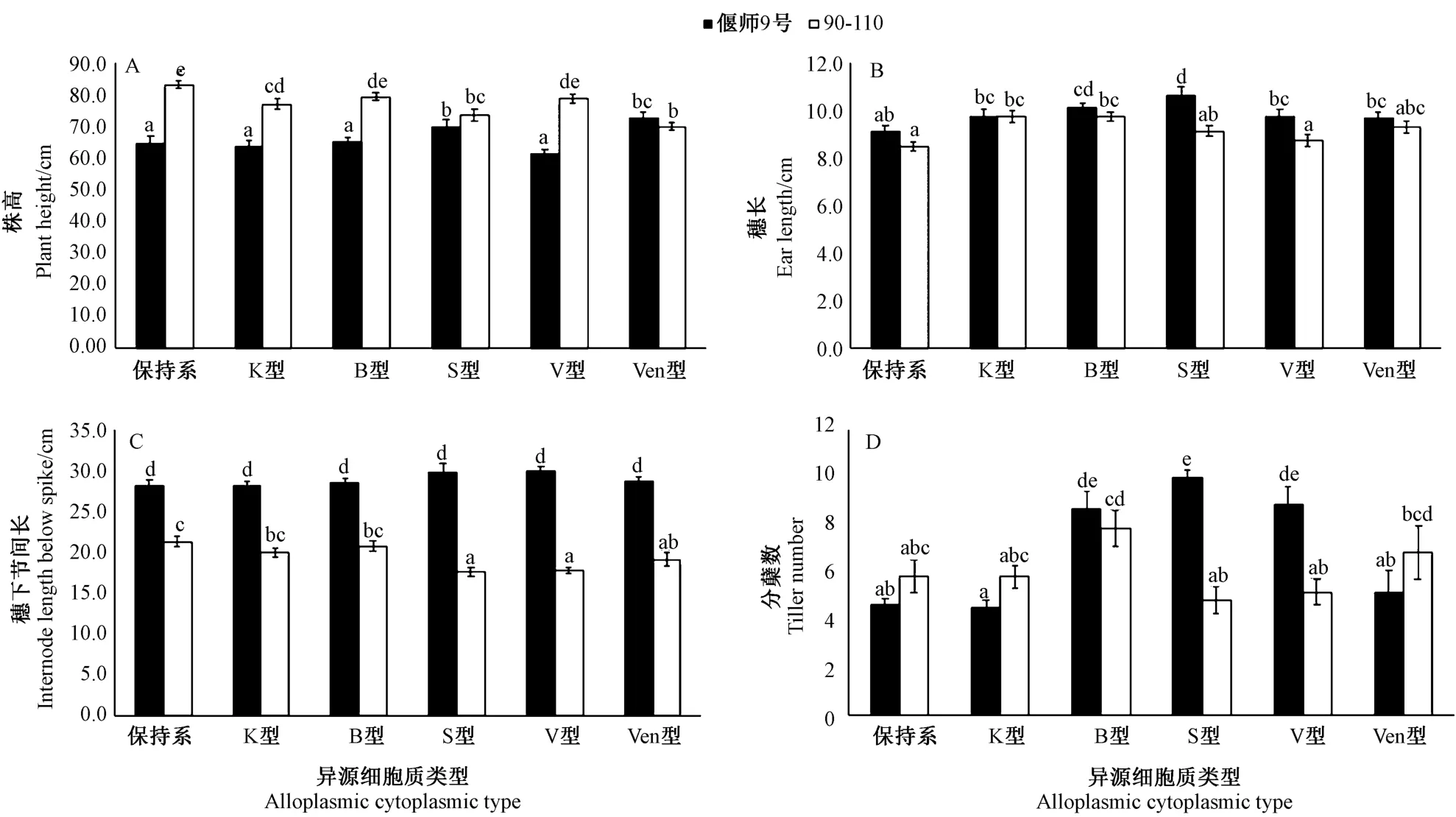
图1 异源细胞质对小麦农艺性状的影响Fig.1 Effects of alloplasmic cytoplasm on agronomic traits in wheat
2.2 异源细胞质对偃师9 号核型和90-110 核型小麦不育系花粉粒核分裂的影响
用1.5%醋酸洋红对不同发育时期偃师9 号核质互作雄性不育系及其保持系花粉粒进行染色,观察核分裂特征,结果显示:单核早期,偃师9 号保持系花粉粒核位于花粉粒中央,至单核晚期时,受中央液泡膨胀影响,花粉粒核被挤压至花粉粒一侧,此时期又称单核靠边期(图2-a1、a2);花粉粒进一步发育至二核期,花粉粒内有2 个点状核,分别是营养核和生殖核,随着生殖核的进一步分裂,形成三核期花粉粒,此时花粉粒内有1 个点状的营养核和2 个鱼状的精核(图2-a3、a4)。与偃师9 号保持系相比,K-偃师9 号单核早期和单核晚期花粉粒核型与偃师9 号保持系的核型一致,发育正常(图2-a1、b1、a2、b2),而至二核期和三核期时,K-偃师9 号花粉粒核不能正常分裂,具体表现为二核期核分裂迟缓(图2-a3、b3),三核期花粉粒中既有单核又有不正常的二核和三核(非鱼状精核)(图2-a4、b4);B-偃师9 号花粉粒核在单核早期、单核晚期和二核期发育正常(图2-a1、c1、a2、c2、a3、c3),而三核期花粉粒核不能正常形成1 个营养核和2 个呈鱼状的精核(图2-a4、c4);S-偃师9 号单核早期花粉粒呈不规则球形,且有部分花粉粒表现出无核现象(图2-a1、d1),至单核晚期时,同一视野下,无核花粉粒占比明显增加(图2-a2、d2),至二核期和三核期时,S-偃师9 号花粉粒均呈无核的椭圆形或多边形(图2-a3、d3、a4、d4);V-偃师9 号单核早期和单核晚期花粉粒核发育正常(图2-a1、e1、a2、e2),而二核期至三核期发育迟缓,具体表现为大多数花粉粒仅含1 个核或3 个点状核(图2-a3、e3、a4、e4);Ven-偃师9 号花粉粒在单核早期核发育正常(图2-a1、f1),而单核晚期至二核期时,出现少量无核花粉粒(图2-a2、f2、a3、f3),至三核期,Ven-偃师9 号花粉粒核异常分裂,部分花粉粒表现出多核现象(图2-a4、f4)。
用1%KI-I2对偃师9 号核质互作雄性不育系及其保持系成熟期花粉粒进行染色,结果显示:偃师9 号保持系花粉粒因淀粉积累饱满而呈蓝黑色(图2-a5),而K-偃师9 号、B-偃师9 号和V-偃师9 号花粉粒虽呈深蓝色,但因淀粉积累不充分其边缘未染色或染色较浅,属于典型的染败类型(图2-b5、c5、e5),S-偃师9号成熟花粉粒因其内部不存在淀粉积累,表现为黄褐色,加之核型在单核早期异常,属于典型的典败类型(图2-d5),Ven-偃师9 号成熟花粉粒少量未着色且形状不规则,但仍有部分花粉粒染色呈浅蓝色,且其边缘未染色,属于典染杂合型败育类型(图2-f5)。
用1.5%醋酸洋红对不同时期90-110 核质互作雄性不育系及其保持系花粉粒进行染色,观察核分裂特征,结果显示:单核早期,90-110 保持系花粉粒核位于花粉粒中央,至单核晚期,因中央液泡膨胀,细胞核被挤压至花粉粒的一侧(图3-a1、a2),至二核期,花粉粒核分裂为1 个营养核和1 个生殖核(图3-a3),进一步发育到三核期时,营养核不再分裂,生殖核正常有丝分裂形成2 个鱼状精核(图3-a4)。 单核早期,K-90-110 花粉粒与90-110 保持系花粉粒核型一致,发育正常,至单核晚期时,出现少量无核且形状不规则的花粉粒(图3-a1、b1、a2、b2),且二核期至三核期表现为大多数花粉粒中仅有1 个核或2 个核(图3-a3、b3、a4、b4);B-90-110 花粉粒核型在单核早期、单核晚期和二核期与90-110 保持系一致,发育正常(图3-a1、c1、a2、c2、a3、c3),而在三核期不能正常分裂为1 个营养核和2 个鱼状的精核(图3-a4、c4);S-90-110 花粉粒核型仅在单核早期与90-110 保持系花粉粒核型一致(图3-a1、d1),发育至单核晚期时,部分花粉粒呈不规则球形或多边形(图3-a2、d2),二核期出现了少量无核花粉粒(图3-a3、d3),最后至三核期时,S-90-110花粉粒核分裂异常,花粉粒中既有单核,又有不正常的二核和三核(未形成鱼状精核)(图3-a4、d4);V-90-110 在单核早期和单核晚期花粉粒核发育正常(图3-a1、e1、a2、e2),而二核期至三核期表现为大多数花粉粒仅含1 个核或2 个核(图3-a3、e3、a4、e4);Ven-90-110 的花粉粒在单核早期和单核晚期核发育正常(图3-a1、f1、a2、f2),与90-110 保持系花粉粒核型一致,而二核期花粉粒核发育迟缓,仅有1 个核(图3-a3、f3),至三核期时,大多数花粉粒仅含1 个核、2 个核或3 个点状核(图3-a4、f4)。
用1%KI-I2对90-110 核质互作雄性不育系及其保持系成熟期花粉粒进行染色,结果发现:90-110 保持系花粉粒淀粉积累饱满,故染色呈蓝黑色(图3-a5),而K-90-110、B-90-110 和V-90-110 因淀粉积累不充分,染色虽呈蓝黑色,但边缘未染色或染色较浅,属于典型的染败类型(图3-b5、c5、e5);S-90-110因其内部不存在淀粉积累,故染色呈黄褐色,但形状大小接近正常花粉粒,属于典型的圆败类型(图3-d5);Ven-90-110 成熟期花粉粒中有少量未染色且形状不规则,但部分花粉粒染色呈浅蓝色,属于典染杂合型败育类型(图3-f5)。
上述结果表明,S-偃师9 号花粉粒核分裂异常发生在单核早期,属典败型;Ven-偃师9 号花粉粒核分裂异常发生在单核晚期,属典染杂合型;K-偃师9 号和V-偃师9 号花粉粒核分裂异常发生在二核期,属染败型;B-偃师9 号花粉粒核分裂异常发生在三核期,属染败型。 同时,K-90-110 与S-90-110 花粉粒核分裂异常发生在单核晚期,分别属染败型和圆败型;V-90-110 和Ven-90-110 花粉粒核分裂异常均发生在二核期,V-90-110 属染败型,Ven-90-110 属典染杂合型;B-90-110 花粉粒核分裂异常发生在三核期,属染败型。
2.3 异源细胞质对偃师9 号核型和90-110 核型不育系小麦绒毡层降解特征的影响
对偃师9 号核质互作雄性不育系及其保持系各时期花药进行切片,观察绒毡层降解特征。 由图4 可知,单核早期至三核期,偃师9 号保持系花药绒毡层细胞面积呈递减趋势,且各时期间大多呈显著差异(图5-a1~a4)。 K-偃师9 号绒毡层面积在单核早期和单核晚期与偃师9 号保持系无显著差异,而在二核期和三核期分别显著低于和高于偃师9 号保持系(图5-b1 ~b4);同时,B-偃师9 号绒毡层面积在单核早期、单核晚期和三核期与偃师9 号保持系无显著差异,而在二核期显著低于偃师9 号保持系(图5-c1 ~c4);S-偃师9 号绒毡层面积在单核早期、单核晚期和二核期均与偃师9 号保持系无显著差异,而在三核期显著高于偃师9 号保持系(图5-d1~d4);V-偃师9 号绒毡层面积在单核早期显著低于偃师9 号保持系,而在单核晚期、二核期和三核期与偃师9 号保持系无显著差异(图5-e1~e4);Ven-偃师9 号绒毡层面积在单核早期和晚期显著低于偃师9 号保持系,在二核期与偃师9 号保持系无显著差异,而在三核期显著高于偃师9 号保持系(图5-f1~f4)。
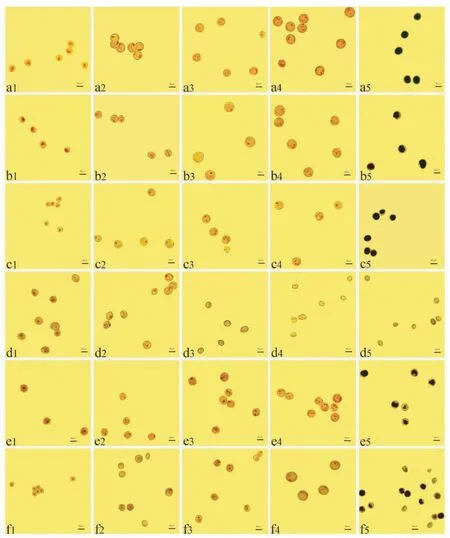
图2 异源细胞质对偃师9 号花粉粒核分裂的影响Fig.2 Effects of alloplasmic cytoplasm on pollen grains division inYanshi9
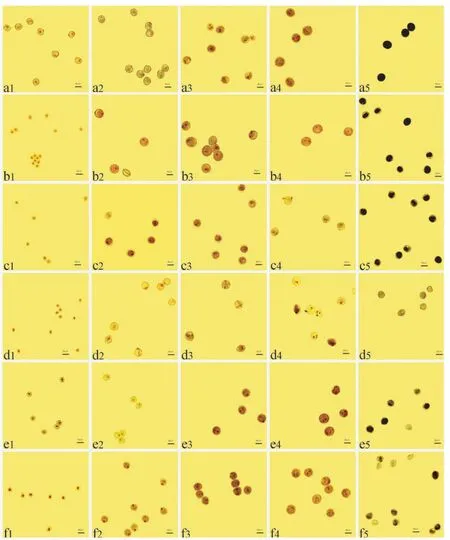
图3 异源细胞质对90-110 花粉粒核分裂的影响Fig.3 Effects of alloplasmic cytoplasm on pollen grains division in 90-110
上述结果表明,K-偃师9 号和B-偃师9 号绒毡层细胞代谢均在二核期开始出现异常,而S-偃师9 号绒毡层细胞异常发生在三核期,具体表现为:K-偃师9号和B-偃师9 号均在单核晚期到二核期降解速度加快,K-偃师9 号绒毡层降解在二核期停滞,而S-偃师9 号绒毡层降解在三核期停滞。 同时,V-偃师9 号和Ven-偃师9 号绒毡层细胞则在单核早期开始出现代谢异常,具体表现为:V-偃师9 号和Ven-偃师9 号绒毡层面积均在单核早期显著低于偃师9 号保持系,且绒毡层降解在单核晚期停滞,二核期恢复,但Ven-偃师9 号在三核期时降解不充分,仍有绒毡层细胞残留。

图4 异源细胞质对偃师9 号绒毡层降解的影响Fig.4 Effects of alloplasmic cytoplasm on tapetum degradation in Yanshi 9
对90-110 核质互作雄性不育系及其保持系各时期花药进行切片,观察绒毡层降解特征。 由图6 可知,单核早期至三核期,90-110 保持系花药绒毡层细胞面积呈递减趋势,且各时期之间均呈显著差异(图7-a1 ~a4)。 K-90-110 绒毡层面积在单核早期、二核期和三核期与90-110 保持系无显著差异,仅在单核晚期显著低于90-110 保持系(图7-b1 ~b4);B-90-110 绒毡层面积在单核早期和单核晚期显著低于90-110 保持系,在二核期和三核期与90-110 保持系无显著差异(图7-c1~c4);S-90-110 绒毡层面积在单核早期显著低于90-110 保持系,而在单核晚期、二核期和三核期与90-110 保持系无显著差异(图7-d1 ~d4);V-90-110 绒毡层面积在单核早期和三核期与90-110 保持系无显著差异,而在单核晚期和二核期显著低于和高于90-110 保持系(图7-e1 ~e4);Ven-90-110 绒毡层面积在单核早期和单核晚期显著低于90-110 保持系,在二核期显著高于90-110 保持系,而在三核期与90-110 保持系无显著差异(图7-f1~f4)。
上述结果表明,K-90-110 和V-90-110 绒毡层细胞代谢均在单核晚期开始出现异常,具体表现为:K-90-110 和V-90-110 可能在单核早期到单核晚期降解速度过快,导致单核晚期绒毡层细胞面积显著低于90-110 保持系,而V-90-110 绒毡层细胞降解可能在单核晚期到二核期间短暂停滞,随后恢复,导致其绒毡层面积在二核期显著高于90-110 保持系。 同时,B-90-110、S-90-110 和Ven-90-110 绒毡层细胞均在单核早期出现代谢异常,具体表现为:B-90-110 和Ven-90-110 绒毡层面积均在单核早期和单核晚期显著低于90-110 保持系,而S-90-110 绒毡层面积仅在单核早期显著低于90-110 保持系,其余时期与保持系均无显著差异。 此外,Ven-90-110 绒毡层降解可能在单核晚期停滞,二核期后恢复正常。
3 讨论
3.1 异源细胞质对小麦农艺性状的影响
雄性不育系是实现作物杂种优势利用的基础,除需具备较高的雄性不育率和配合力外,同时要求综合农艺性状符合生产需求。 因此研究异源细胞质对雄性不育系农艺性状的影响对筛选和创制农艺性状优良的雄性不育系具有重要意义[10]。
本研究以2 套同核异质雄性不育系为材料,探究同质异核雄性不育系的遗传效应,结果发现K 型、B型、S 型、V 型和Ven 型细胞质在不同类型细胞核(1B/1R 类型和非1B/1R 类型)存在时对小麦不同农艺性状存在不同程度的影响。 上述研究结果与易自力等[24]以同质异核小麦为材料,研究小麦属和山羊草属的细胞质遗传效应的结论基本一致,说明在不同核背景下同一异源细胞质的遗传效应不同。 关于不同异源细胞质在相同核背景下对普通小麦主要农艺性状的影响,徐乃瑜等[25]已进行了较为全面和系统的研究,其结果发现:中国春细胞核(非1B/1R 类型)存在时,B型和Ven 型细胞质可促进植株高度和分蘖数增加,但与对照相比无明显差异,同时对穗下节间长和穗长存在负调控效应;与对照相比K 型细胞质可降低株高、缩短穗下节长和增加有效分蘖数,但无明显差异,而对穗长存在极显著的负调控效应;V 型细胞质可极显著降低株高和缩短穗下节间长与穗长,同时也减少有效分蘖数,但与对照相比无显著差异。 上述部分研究结果在本研究中也得到了证实,对同核(非1B/1R 类型)异质核质互作雄性不育系农艺性状调查及统计发现:在非1B/1R 类型细胞核存在时,K 型细胞质对穗下节间比,B 型、V 型和Ven 型细胞质对分蘖数的影响与徐乃瑜等[25]的研究结果完全一致,表明在同一核背景下不同异源细胞质对小麦不同农艺性状的影响程度不同。 表明同质异核和同核异质材料不同农艺性状的遗传效应不同,核质互作雄性不育系各农艺性状的遗传效应是核质相互作用的结果。 该结论在水稻[26]、叶用芥菜[27]和玉米[28]等作物中也得到了证实。
与此同时,在本研究中非1B/1R 类型细胞核存在时,K 型和V 型细胞质对株高,B 型、V 型和Ven 型细胞质对穗下节间长的影响,与徐乃瑜等[25]研究发现的影响趋势相同,但显著性均存在不同程度的差异。 此外,非1B/1R 类型细胞核存在时,B 型和Ven 型细胞质对株高,K 型、B 型、V 型和Ven 型细胞质对穗长,K型细胞质对分蘖数的影响,与前人研究结果不一致。导致上述显著性和影响趋势差异的原因可能是试验材料及播种环境(地域及气候)不同,说明同一异源细胞质在不同遗传背景和环境条件下可能对同一性状产生不同程度的影响。 本研究分析了异源细胞质对小麦株高、穗长、穗下节间长和分蘖数的影响,至于异源细胞质对其他农艺性状的影响以及不同细胞核1B/1R 类型、细胞质和环境(如:光照、温度、湿度等)之间的互作模式对农艺性状的影响还有待进一步研究。
3.2 异源细胞质对小麦花粉粒核分裂的影响
前人研究表明,小麦花粉粒发生败育的时期主要有减数分裂期、单核早期、单核晚期、二核期和三核期等,进一步研究表明,败育发生的时期越早,花粉粒败育越彻底,雄性不育率越高[29]。 目前,关于来自山羊草属不同种的异源细胞质对小麦花粉粒核分裂影响的研究已有报道。 李传友等[30]研究发现K 型不育系花粉核分裂异常发生在二核期或三核期,属浅染败型或深染败型;Ven 型不育系花粉核分裂异常发生在单核晚期至二核期,属圆败型或浅染败型。 王小利等[31]在对两类二角山羊草细胞质小麦雄性不育系的研究发现B 型核质互作雄性不育系在1B/1R 类型细胞核和非1B/1R 类型细胞核存在时花粉粒核分裂异常均发生在三核期,属染败型和圆败型。 本研究中,K 型、B 型和Ven 型核质互作不育系花粉粒核分裂发生异常的时期和花粉败育类型,与前人研究结果基本一致。 本研究以2 套同核异质小麦为材料,较为系统地研究了K 型、B 型、S 型、V 型和Ven 型细胞质对小麦花粉粒核分裂的影响,在印证前人结论的同时发现不同异源细胞质在相同核背景下对不育系花粉粒核分裂的影响不同,这与李凤霞等[32]对烟草2 种同核异质不育系小孢子发育进行细胞学观察后得出的结论一致。
本研究还发现同一异源细胞质在不同核背景下对小麦花粉粒核分裂及败育类型的影响不完全相同,如在1B/1R 和非1B/1R 类型细胞核存在时,K 型、S 型、V 型和Ven 型核质互作雄性不育系花粉粒核分裂发生异常的时期和败育类型均存在不同程度的差异。 因此,可将上述5 种小麦核质互作雄性不育系分为以下三大类型:1)花粉粒核分裂异常发生的时期以及败育类型与细胞核1B/1R 类型(1B/1R 类型和非1B/1R类型)无关,包括B 型和V 型;2)花粉粒核分裂异常发生的时期与细胞核1B/1R 类型(1B/1R 类型和非1B/1R 类型)有关,但败育类型与细胞核1B/1R 类型(1B/1R 类型和非1B/1R 类型)无关,包括K 型和Ven 型;3)花粉粒核分裂异常发生的时期以及败育类型均与细胞核1B/1R 类型(1B/1R 类型和非1B/1R 类型)有关,包括S 型。 综上,同核异质与同质异核材料的败育机理不完全相同,核质互作雄性不育系的花粉败育与异源细胞质和不同1B/1R 类型细胞核的互作模式有关,即核质互作雄性不育系的花粉败育是核质互作的结果,与韩笑冰等[33]对小麦不同细胞质不育系花粉败育进行细胞学比较研究后得出的结论完全一致。
3.3 异源细胞质对小麦绒毡层降解特征的影响
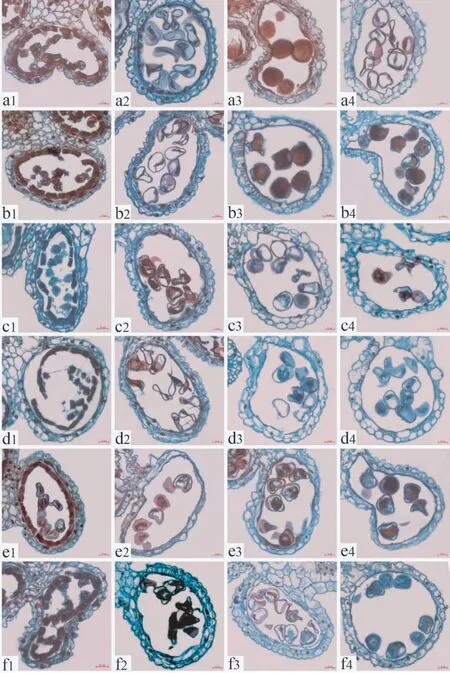
图7 异源细胞质对90-110 绒毡层降解的影响(石蜡切片)Fig.7 Effects of alloplasmic cytoplasm on tapetum degradation in 90-110 (paraffin section)
绒毡层是小麦花药壁最内层细胞,其在降解过程中向药室释放大量用于花粉粒发育的结构和营养物质[34-36]。 张鹏飞等[12]对小麦S 型1376 不育系及其保持系绒毡层发育过程进行比较后发现,绒毡层降解代谢发生紊乱,致使花粉粒发育过程中物质供应不足,使花粉粒核分裂出现异常,这是导致花粉败育的主要原因。 上述结果在本研究中也得到了证实,如2 套核质互作雄性不育系不同时期花药绒毡层细胞降解均与其对应保持系存在不同程度的差异,而其花粉粒核分裂均在不同发育阶段出现不同程度的异常。 花药绒毡层降解被认为是一种典型的细胞程序化死亡过程,刘子涵等[37]研究表明,小麦绒毡层细胞程序化死亡的延迟启动是花粉败育的细胞学原因。 这一结论在本研究中也得到了证实,如K-偃师9 号绒毡层降解在二核期停滞,与偃师9 号保持系绒毡层降解相比存在显著延迟,导致其花粉粒核分裂在二核期发生异常;V-偃师9 号和Ven-偃师9 号绒毡层降解在单核晚期停滞,其花粉粒核分裂异常分别发生在二核期和单核晚期。 此外,本研究还发现S-偃师9 号绒毡层降解异常发生在三核期,然而其花粉粒异常分裂发生在单核早期,表明可能另有原因导致了核分裂异常。 Liu 等[38]对甘蓝型油菜核质互作雄性不育系进行细胞学研究,发现花药绒毡层和中层细胞过早的细胞程序化死亡是花粉败育的主要原因,指出绒毡层异常降解并非花粉败育的唯一原因,至于本研究中核质互作雄性不育系S-偃师9 号花粉核分裂异常是否与花药中层细胞降解异常有关还有待进一步研究。
本研究发现同质异核不育系的绒毡层降解特征不同,具体表现为K 型、B 型、S 型、V 型和Ven 型细胞质在1B/1R 类型细胞核和非1B/1R 类型细胞核存在时,不育系绒毡层降解特征均存在不同程度的差异。 表明核质互作雄性不育系绒毡层细胞代谢紊乱与异源细胞质和不同1B/1R 类型细胞核的互作模式有关,即核质互作雄性不育系的绒毡层降解代谢异常可能是核质互作的结果。
4 结论
本研究结果表明,异源细胞质分别与1B/1R 类型细胞核和非1B/1R 类型细胞核间存在互作模式的差异,不同程度地影响核质互作雄性不育系的农艺性状,同时引发花药绒毡层细胞差异性代谢异常,最终导致花粉粒核分裂异常,产生不同的败育类型。 本研究结果为进一步揭示小麦核质互作雄性不育机理和更好地利用异源核质互作雄性不育系提供了坚实的理论依据。
