单增李斯特菌inl A/ inl B双基因缺失株生长特性观察及对小鼠致病性研究
2016-08-25任静静杨铭伟陈云飞蒋建军王鹏雁
任静静,杨铭伟,陈云飞,蒋建军,王鹏雁
单增李斯特菌inl A/ inl B双基因缺失株生长特性观察及对小鼠致病性研究
任静静,杨铭伟,陈云飞,蒋建军,王鹏雁
目的探究单增李斯特菌inl A/inl B双基因缺失株的生物学特性。方法通过体外胁迫培养条件下OD600测定以及小鼠感染试验,对Lm90-△inlAB和Lm90的抗胁迫能力及细菌致病力进行研究。结果低温(4 ℃、30 ℃)环境下,Lm90和Lm90-△inlAB生长差异无统计学意义(t4 ℃=0.057,P>0.05;t30 ℃=0.441,P>0.05),适温37 ℃及42 ℃高温环境下,Lm90-△inlAB较Lm90生长受到抑制,生长差异具有统计学意义(t37 ℃=0.763,P<0.05;t42 ℃=1.147,P<0.05);体外耐酸碱试验中,Lm90-△inlAB耐酸性未明显改变,而耐碱能力强于亲本株Lm90,生长差异具有统计学意义(pH=9时t=2.837,P<0.05);在含有3.5%酒精的BHI培养基及含5% NaCl高渗BHI培养基中,Lm90-△inlAB增长趋势明显低于Lm90,生长差异具有统计学意义(t酒精=1.422,P<0.05;tNaCl=1.382,P<0.05),Lm90-△inlAB对酒精及高盐的耐受力显著低于Lm90;小鼠毒力测定结果表明,Lm90-△inlAB与Lm90半数致死量分别为106.94CFU、105.68CFU,Lm90-△inlAB毒力下降明显,在不同检测时间点Lm90-△inlAB在小鼠肝脏和脾脏内的载菌量均少于Lm90,载菌量差异具有统计学意义(t肝=2.454,P<0.05;t脾=2.443,P<0.05)。结论Lm的抗胁迫能力可能随着基因的缺失发生显著改变,内化素InlA/InlB与单增李斯特菌侵染宿主的能力有一定的调控作用。缺失株生物学特性测定对研究Lm胁迫环境生存及毒力因子的致病机理提供了科学依据。
单核细胞增生性李斯特菌;inlA基因;inlB基因;环境耐受;毒力
Supported by the Program of the National Natural Science Foundation of China (No. 31260606), the Open Fund Project of State Key Laboratory of Veterinary Etiological Biology (No. SKLVEB2011KFKT007) and the High Level Talents Scientific Research Project of Shihezi University (No. RCZX201104 )
单核细胞增生性李斯特菌(Listeriamonocytogenes,Lm)是目前国际公认的重要食源性病原菌之一。自1926年Murray等首次分离到本病原后,李斯特菌病现已呈世界性分布[1]。Lm做为一类重要的人兽共患的条件致病菌,可引起人类和多种畜禽的疾病,人畜感染后主要表现为脑膜炎、败血症;家禽感染后主要表现为脑膜炎、坏死性心肌炎及坏死性肝炎[2-3]。致病性李斯特菌为侵袭性胞内菌,其共同特点是可穿越感染宿主的3道屏障,即肠道屏障、血脑屏障和胎盘屏障[4]。内化素A(InlA)与内化素B(InlB)作为Lm特有的与黏附和侵袭相关的毒力因子,位于菌体表面,介导细菌侵袭入宿主细胞并内化至吞噬小体,从而建立感染。在细菌穿越肠道上皮细胞和胎盘屏障中,InlA是必需的[5],由800个氨基酸组成,N端具有亮氨酸重复(LRR)区域,并由中间重复区(IR)与B重复区隔开,该LRR与IR区域为Lm入侵上皮细胞所必需。InlA的C端所包含的LPXTG基元能够以共价键连接肽聚糖,在细菌穿越肠道屏障中发挥重要作用[6]。InlB可介导Lm侵入肝细胞和非上皮细胞等[7],由630个氨基酸组成,其N端到C端分别为信号肽序列、LRR重复区、IR重复区、B重复区和GW区。其中GW区可介导InlB以非共价键连接脂磷壁酸吸附至细菌细胞壁表面,LRR重复区及B重复区也参与InlB与靶细胞的互作[6]。肝细胞生长因子受体(Met)为InlB的主要受体,在LRR重复区的凹陷部与InlB结合。而补体分子Clq受体为InlB的另一个受体,该受体同时可抑制不同细胞HIV的生长[8]。
本文以同源重组构建的inl A、inl B双基因缺失株(Lm90-△inlAB)为研究对象,研究Lm90-△inlAB的抗胁迫能力变化并通过小鼠感染试验探究缺失株毒力的改变,为探讨Lm毒力因子InlA、InlB的致病机理提供科学依据。
1 材料与方法
1.1材料
1.1.1菌株和实验动物单增李斯特菌Lm90(4b血清型)新疆致绵羊脑炎分离株,由石河子大学动物科技学院预防兽医学实验室分离、鉴定并保存,基因缺失株Lm90-△inlAB本实验室自行构建。8周龄健康昆明系小鼠由石河子大学实验动物中心提供。
1.1.2主要试剂脑心浸液肉汤BHI,购自BD公司;PBS(pH=7.2)实验室配制。
1.1.3主要仪器设备生物安全柜(BHC-Ⅱ-A/B3型)购自The Baker Company;HANNA PH211台式酸度计购自ROMANIA Company;BioTeck Power Wave XS2 连续波长酶标仪购自美国Gene Limited Company。
1.2方法1.2.1Lm 90与Lm 90-△inl AB不同温度下生长曲线测定将Lm 90和Lm 90-△inlAB按照1∶100的比例分别接种到10 mL的BHI培养基中,37 ℃180 r/min条件下过夜培养至OD600为0.4。后将Lm 90和Lm 90-△inlAB按照1∶100的比例分别接种到20 mL的BHI中,每个菌株接种4管分别置于4 ℃、30 ℃、37 ℃、42 ℃于180 r/min条件下培养;以加入细菌的时刻为时间起点(t=0),在t=1、2、4、6、8、10、12 h 7个不同的时间点取200 μL菌液加入到96孔酶标板中,测定两株菌的OD600值并记录。根据不同时间点OD600值绘制Lm 90和Lm 90-△inlAB在4种不同温度条件下的生长曲线并比较分析。
1.2.2Lm90与Lm90-△inl AB耐酸、耐碱生长曲线测定Lm90和Lm90-△inlAB培养同1.2.1(BHI培养基pH=7.0)。将Lm90和Lm90-△inlAB按照1∶100的比例分别接种到20 mL不同pH(pH=3.0、pH=5.0、pH=7.0、pH=9.0)的BHI液体培养基中进行培养。以加入细菌的时刻为时间起点(t=0),在t=1、2、4、6、8、10、12 h 4个不同的时间点取200 μL菌液加入到96孔酶标板中,测定2株菌的OD600值并记录。根据不同时间点的OD600值,绘制Lm90和Lm90-△inlAB在4种不同pH条件下的生长曲线并进行分析。
1.2.3Lm90与Lm90-△inl AB高盐、酒精试验
Lm90和Lm90-△inlAB培养同1.2.1。将Lm90和Lm90-△inlAB按照1∶100的比例分别接种到20 mL含有3.5%酒精BHI液体培养基、5% NaCl BHI液体培养基中培养。以加入细菌的时刻为时间起点(t=0),在t=1、2、4、6、8、10、12 h 7个不同的时间测定2株菌的OD600值并记录。根据不同时间点的OD600值,绘制Lm90和Lm90-△inlAB在酒精及高盐条件下的生长曲线,并进行比较分析。
1.2.4Lm90与Lm90-△inl AB LD50的测定Lm90与Lm90-△inl AB 过夜振荡培养。当OD600为0.6时,8 000 r/min离心5 min收集细菌。然后将管底的细菌用PBS清洗2遍,用PBS将细菌悬液进行10倍梯度稀释。用不同稀释度的菌液对小鼠腹腔注射,确定2株细菌的绝对致死剂量(Absolute lethal dose,LD100)和最小致死剂量(Minimal lethal dose,MLD)。将小鼠分成10组,每组10只,然后将稀释后的菌液腹腔注射8周龄昆明系小鼠,每个梯度10只,100 μL/只,Lm90与Lm90-△inl AB各分5个梯度,连续观察14 d,统计死亡数。对照组注射等剂量PBS,100 μL/只。利用寇氏法计算LD50,计算公式:log LD50=[Xm-i(∑P-0.5)]。式中:Xm为最大剂量的对数;P为各组动物的死亡率(以小数表示);∑P为各组动物死亡率的总和(P1+P2+P3……);i为组间距(相邻两组对数剂量的差值)。
1.2.5小鼠肝脏、脾脏载菌量的测定8周龄健康昆明系小鼠随机分2组,每组10只。Lm90与Lm90-△inl AB过夜活化培养,将活化后的菌液按1∶100分别接种到新鲜BHI中,当其OD600值为0.3时,分别给每组中的每只小鼠腹腔注射0.1 mL的菌液(亚致死量)。注射菌液的时刻t=0(day),分别在t=2,4,6,8,10 d时每组取出1只小鼠,无菌条件下剖检取出肝、脾,加入1 mL的PBS液研磨均匀充分后进行10倍系列稀释,分别均匀涂布到BHI固体培养基上37 ℃条件下培养24~48 h,进行标准细菌平板计数。
1.2.6统计学分析应用SPSS20.0统计软件,计量资料分析采用独立样本t检验。P<0.05表示差异具有统计学意义。
2 结 果
2.1不同温度Lm90与Lm90-△inl AB的生长曲线将Lm90与Lm90-△inlAB分别置于不同温度条件下培养,结果发现2株菌在4 ℃生长极其微弱,生长状态基本相同,生长差异无统计学意义(t=0.057,P>0.05);在30 ℃条件下2株菌均表现出生长适应期延长,6 h左右才达到稳定期,生长差异无统计学意义(t=0.441,P>0.05);37 ℃与42 ℃培养条件下,2株菌生长迅速,Lm90与Lm90-△inlAB在0至2 h时间段内均处于生长适应期, 2至4 h时间段内处于生长对数期,4至8 h时间段内处于生长稳定期,在生长适应期二者生长情况基本一致,而2 h之后Lm90生长趋势明显高于Lm90-△inlAB,生长差异具有统计学意义(37 ℃:t=0.763,P<0.05;42 ℃:t=1.147,P<0.05),表明Lm90耐高温能力显著强于Lm90-△inlAB(如图1所示)。
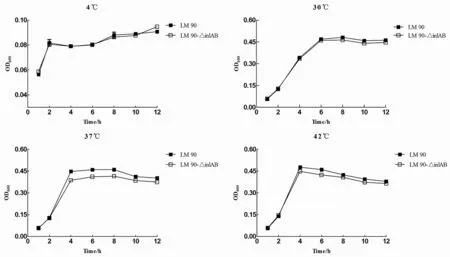
图1 不同温度条件下Lm 90与Lm 90-△inl AB生长曲线Fig.1 Growth curve of Lm 90 and Lm 90-△inl AB in different temperature
2.2不同酸度Lm90与Lm90-△inl AB的生长曲线如图2所示Lm90与Lm90-△inlAB在pH=3.0、pH=5.0条件下培养,两株菌生长状态基本相同,均表现出生长停滞,生长差异无统计学意义(pH=3.0时t=1.079,P>0.05;pH=5.0时t=0.333,P>0.05);pH=7.0时,在检测的12 h内二者生长趋势基本一致,生长差异无统计学意义(t=0.156,P>0.05);pH=9.0时,在0至2 h时间段内二者均处于生长适应期,而在2 h之后二者均进入对数生长期,Lm90生长趋势明显高于Lm90-△inlAB,生长差异具有统计学意义(t=2.837,P<0.05),表明Lm90耐碱能力强于Lm90-△inlAB。
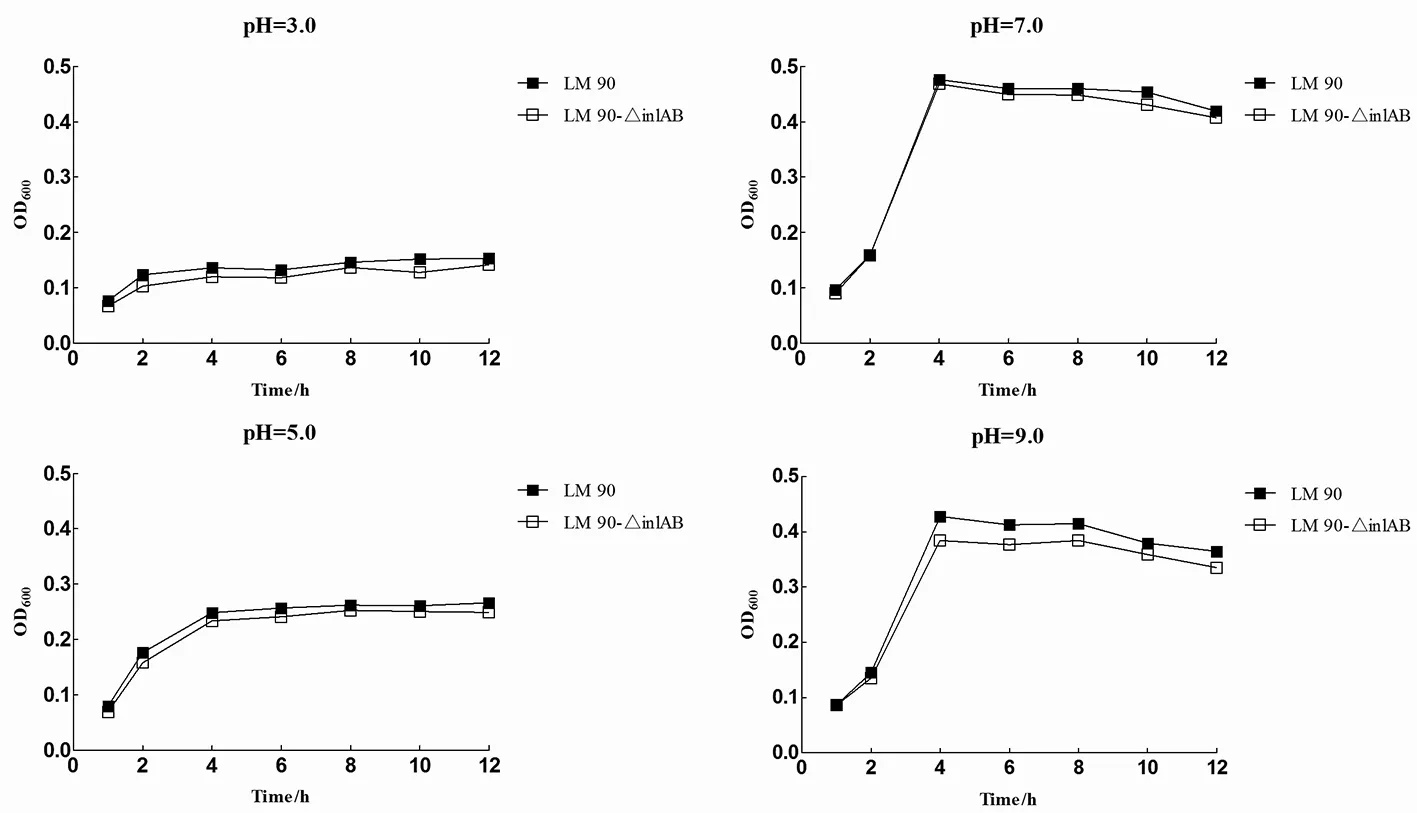
图2 不同pH条件下Lm 90与Lm 90-△inl AB生长曲线Fig.2 Growth curve of Lm 90 and Lm 90-△inl AB in different pH
2.3Lm90与Lm90-△inl AB酒精、NaCl耐受性结果将Lm90与Lm90-△inlAB分别接种BHI、含3.5%酒精BHI和5.0%NaCl BHI,观察发现二者在含3.5%酒精BHI和5.0%NaCl BHI中均生长缓慢,生长延滞现象明显且生长量较低,生长情况与空白BHI培养基相比差异具有统计学意义(Lm90:t酒精=1.157,P<0.05;tNacl=1.169,P<0.05;Lm90-△inl AB:t=0.669,P<0.05;t=1.281,P<0.05)(图3-A、3-B);分别对比Lm90与Lm90-△inlAB发现,在0至4 h时间段内二者均处于生长适应期,而在4 h之后Lm90增长趋势明显高于Lm90-△inlAB,生长差异具有统计学意义(t酒精=1.422,P<0.05;tNaCl=1.382,P<0.05),表明Lm90-△inlAB对酒精及高盐的耐受力低于Lm90(图3-C、3-D)。
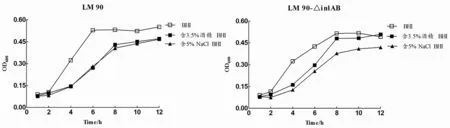
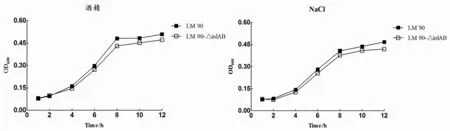
图3 含3.5%酒精、5.0%NaCl BHI中Lm 90与Lm 90-△inl AB生长曲线Fig.3 Growth curve of Lm 90 and Lm 90-△inl AB in different medium
2.4Lm90与Lm90-△inl AB 的LD50测定结果
试验小鼠经腹腔注射Lm90与Lm90-△inlAB,12 h后出现精神萎靡、畏寒扎堆、采食量与饮水量下降,24 h后部分小鼠陆续出现死亡现象。连续观察10 d,统计各组小鼠死亡率并记录结果。根据寇氏算法计算Lm90与Lm90-△inlAB的LD50分别为105.68CFU和106.94CFU(如表1),Lm90-△inlAB的LD50升高了1.26个对数数量级,表明inl A与inl B缺失后使得Lm毒力下降。
表1Lm 90和Lm 90-△inlAB对小鼠LD50的测定结果
Tab.1LD50measurement of Lm 90 & Lm 90-△inlAB

Lm90Lm90-△inlAB组别Group剂量Dose动物数No.ofanimals死亡数Deathno.死亡率MortalityLD50Medianlethaldose组别Group剂量Dose动物数No.ofanimals死亡数Deathno.死亡率MortalityLD50Medianlethaldose11.9×10910101.0105.6812.2×10910101.0106.9421.9×1081090.922.2×1081080.831.9×1071090.932.2×1071060.641.9×1061080.842.2×1061050.551.9×1051050.552.2×1051000
2.5Lm90与Lm90-△inl AB组织载菌量测定结果两组小鼠接种Lm90与Lm90-△inlAB后,小鼠出现精神萎靡,双眼微闭,结膜潮红有少量分泌物,活动迟缓等现象。病理解剖发现,接种Lm90组的小鼠脾脏和肝脏明显肿大,肠道黏膜脱落,部分肠道出现充血、出血现象,而Lm90-△inlAB注射组小鼠脾脏与肝脏肿大但相比Lm90注射组不明显;Lm90组小鼠在第2、4、6、8、10 d的肝脏载菌量结果分别为103.9、104.2、104.7、103.5、102.8CFU,脾脏的细菌计数结果分别为103.7、104.0、104.5、103.1、102.4CFU;Lm90-△inlAB组小鼠在第2、4、6、8、10 d肝脏的细菌计数结果分别为102.8、103.2、103.8、101.8、101.5,脾脏的细菌计数结果分别为102.2、102.9、103.3、101.5、101.2。
分析计数结果可以发现,在各个时间节点注射Lm90组的小鼠肝和脾脏载菌量要高于注射Lm90-△inlAB肝和脾载菌量。计数结果取对数绘制Time-log值的柱状图(如图4所示),发现在第0 d至6 d,肝和脾内的细菌数量呈上升趋势,并且肝和脾内的细菌含量都较多;在第8 d和第10 d的时候,2组小鼠的肝脏和脾脏内的细菌含量明显减少,载菌量差异具有统计学意义(t肝=2.454,P<0.05;t脾=2.443,P<0.05)。

图4 小鼠肝脏和脾脏载菌量计数结果Fig.4 Amount of bacteria in the liver and spleen of mice
3 讨 论
Lm作为一种重要的食源性致病菌,在自然界广泛存在,不仅给畜牧业带来巨大的经济损失,同时也严重威胁公共卫生安全。作为一种胞内寄生的模式菌,其感染过程包括抵抗外界环境与宿主内环境的应激、黏附并侵入细胞,在细胞内增殖以及在细胞间扩散,每一步均由特定的毒力因子介导[9]。
本试验重点研究亲本株Lm90与缺失株Lm90-△inlAB在低温、高温、不同酸度以及高盐、酒精等压力环境下Lm的生长状况并初步对其毒力进行判定。Lm生长温度范围为2~42 ℃,最适生长温度范围为30~37 ℃,能在普通冰箱冷藏室生长,是一种典型的耐冷性细菌[10]。本研究发现inlA、inlB基因对Lm抵抗低温逆境不是必需的,但当处于高温环境时,Lm90-△inlAB生长趋势明显低于Lm90,生长差异具有统计学意义,推测可能是由于基因缺失后使其对高温的耐受力下降。
有研究证实Lm嗜中性,最适pH为6~7[11]。酸碱实验测定结果表明,于酸性、中性环境下,Lm90与Lm90-△inlAB相比,生长差异无统计学意义;碱性环境中Lm90耐碱能力显著强于Lm90-△inlAB,生长差异具有统计学意义;有报道指出Lm对盐有较高的耐受性,在浓度为7%的氯化钠中都能生长,本试验中Lm90在5%的盐溶液中生长良好,表现出一致性,而Lm90-△inlAB对高盐的耐受力明显下降,与Lm90相比较,生长差异具有统计学意义。
在肝脾载菌量试验中Lm90-△inlAB在小鼠肝脏和脾脏中的数量明显少于Lm90,载菌量差异具有统计学意义,对小鼠的感染能力减弱,推测原因可能是由于InlA参与介导Lm的内化途径[13-14],当inlA缺失后其介导作用受阻,从而使Lm毒力下降;另外内化素家族成员在侵染宿主的过程中往往具有协同作用,例如在InlB和InlC存在的条件下InlA介导的侵染能力会增强,而在inlB缺失的情况下InlA介导的入侵作用明显下降[15],本试验中Lm90-△inlAB毒力的下降与其表现出一致性,推测原因可能是肝脏和脾脏中InlB的受体不同(Met和Clq)所致。
本研究通过体外胁迫培养以及小鼠感染试验,对Lm90-△inlAB和Lm90的抗胁迫能力及细菌致病力进行了研究,在一定程度上证明了inlA与inlB对Lm毒力有一定的调节作用,为进一步阐明Lm的毒力因子的致病机制提供了一定的科学依据。
[1] Chao GX, Jiao XA, Xu Q, et al. Study on epidemic status and drug resistance ofListeriamonocytogenesin food[J]. Chin J Heal Lab Technol, 2005,15(5): 519-521. DOI:10.3969/j.issn.1004-8685.2005.05.003.(in Chinese)
巢国祥,焦新安,徐勤,等.8类食品单核细胞增生李斯特菌流行特征及耐药性状研究[J]. 中国卫生检验杂志, 2005,15(5): 519-521.
[2] Birte FV, Hans HH, Bente O, et al. Elucidation ofListeriamonocytongenescontramination routes in cold-smoked salmon processing plants detected by DNA-based typing methods[J]. Appl Envir Mic Robiol, 2001, 67(6): 2586-2595. DOI: 10. 1128/AEM. 67. 6. 2586-2595. 2001
[3] Laemynck GV, Lafarge V, scotter S. Improvement of the detection ofListeriamonocytogenesby the application of ALOA, a diagnostic, chromogenic isolation medium[J]. J Appl Microbiol, 2000, 88: 430-441. DOI: 10. 1046/j. 1365-2672. 2000. 00978. x
[4] Vazquez-Boland JA, Kuhn M, Berche P, et al.Listeriapathogenesis and molecular virulence determinants[J]. Clin Microbiol Rev, 2001, 14(548): 584-640. DOI: 10. 1128/CMR.14. 3. 584-640. 2001
[5] Lecuit M, Nelson DM, Smith SD, et al. Targeting and crossing of the human maternofetal barrier byListeriamonocytogenes: role of internalin interaction with trophoblast E-cadherin[J]. Proc Natl Acad Sci U S A, 2004, 101(16): 6152-6157. DOI: 10. 1073/pnas. 0401434101
[6] Dussurget O, Pizarro-Cerda J,Cossart P. Molecular determinants ofListeriamonocytogenesvirulence[J]. Annu Rev Microbiol, 2004,58:587-610. DOI: 10. 1146/annurev. Micro. 57. 030502. 090934
[7] Hamon M, Bierne H, Cossart P.Listeriamonocytogenes: a multifaceted model[J]. Nat Rev Microbiol, 2006, 4(6): 423-434. DOI: 10. 1038/nrmicro1413
[8] Ghebrehiwet B, Jesty J, Peerschke EI. gClq-R/p33: structure function predicitions from the crystal structure[J]. Immunobiology, 2002, 205(4/5): 421-432. DOI: 10. 1078/0171-2985-00143
[9] Jiang LL, Chen JS, Fang WH. Study on the main virulence factors ofListeriamonocytogenes[J]. Chin J Zoonoses, 2007, 23(7): 733-736. (in Chinese)
江玲丽,陈建舜,方维焕.单核细胞增多性李斯特菌主要毒力因子研究进展[J]. 中国人兽共患病学报, 2007, 23(7): 733-736.
[10] Zhou XH, Li XY, Yang XS, et al. Study the effect of cultural condition on growth state ofListeriamonocytogenes[J]. Sci Technol Food Indus, 2013,15: 140-148.DOI:10.13386/j.issn.1002-0306.2013.15.013. (in Chinese)
周小红,李学英,杨宪时,等. 培养条件对单增李斯特菌生长的影响[J]. 食品工业科技, 2013,15: 140-148.
[11] Abee T, Wouters JA. Microbial stress response in minimal processing[J]. Int J Food Microbiol, 1999, 50(1/2): 65-91. DOI: 10. 1016/S0168-1605(99)00078-1
[12] Schubert WD, Urbanke C, Ziehm T, et al. Structure of internalin a major invasion protein ofListeriamonocytogenesin complex with its human receptor E-cadherin[J]. Cell, 2002, 111(6): 825-836. DOI: 10. 1016/S0092-8674(02)01136-4
[13] Shen Y, Ujokas M, Park M, et al. Inl B-dependent internalization of listeriais mediated by the met receptor tyrosine kinas[J]. Cell, 2000, 103(3): 501-510. DOI: 10. 1016/S0092-8674(00)00141
[14] Ireton K. Entry of the bacterial pathogenListeriamonocytogenesinto mammalian cells[J]. Cell Microbiol, 2007, 9(6): 1365-1375. DOI: 10.1111/j. 1462-5822. 2007. 00933. x
Biological characters and pathogenicity ofListeriamonocytogenes△inlA/△inlB double gene deletion strains
Ren Jing-jing, Yang Ming-wei, Chen Yun-fei, Jiang Jian-jun, Wang Peng-yan
(CollegeofAnimalScienceandTechnology,ShiheziUniversity,Shihezi832000,China)
In order to explore the biological characters of the △inlA/△inlB double gene deletion strains ofListeriamonocytogenes, the ability to resist stress and pathogenicity ofLm90 andLm90-△inlAB were studied byinvitrostress culture and mouse infection experiment. Results showed that there was no significant difference(t4 ℃=0.057,P>0.05;t30 ℃=0.441,P>0.05)betweenLm90 andLm90-△inlAB at low temperature (4-30 ℃) and the growth ofLm90 was better than that ofLm90-△inlAB at 37 ℃ and 42 ℃(t37 ℃=0.763,P<0.05;t42 ℃=1.147,P<0.05). For acid and alkali resistance testinvitro, there was no obvious change of acid resistant ability but a better alkali resistant ability ofLm90-△inlAB than parent strainLm90(pH=9,t=2.837,P<0.05). The growth ofLm90-△inlAB was significantly lower thanLm90 in BHI culture medium containing 3.5% alcohol and 5% NaCl hypertonic BHI medium and the difference was significant(talcohol=1.422,P<0.05;tNaCl=1.382,P<0.05). In other words, the tolerance of alcohol and high salt ofLm90-△inlAB was significantly lower than that ofLm90. Results of mice virulence determination showed that the median lethal dose ofLm90-△inlAB andLm90 were 106.94CFU and 105.68CFU respectively, and the virulence ofLm90-△inlAB decreased significantly. The microbial load ofLm90-△inlAB in mice’s liver and spleen was much less than that ofLm90 at different time points(tliver=2.454,P<0.05;tspleen=2.443,P<0.05). Results reveal that the ability to resist stress ofLm90 could have a significant change as genetic absence. Internalization InlA/InlB had a certain regulation effect on the ability ofListeriamonocytogenesto infect host. Determination of biological characteristics of deletion strains provides a scientific basis for exploringListeriamonocytogenessurvival under stress environment and pathogenic mechanism of virulence factors.
Listeriamonocytogenes; inlA gene; inlB gene; environment tolerance; virulence
Jiang Jian-jun, Email: jiangjianjun@shzu.edu.cn
蒋建军,Email: jiangjianjun@shzu.edu.cn
石河子大学动物科技学院,石河子832000
R378.99
A
1002-2694(2016)06-0529-06
2015-09-14;
2016-04-03
DOI:10.3969/j.issn.1002-2694.2016.06.005
国家自然科学基金项目(No. 31260606),家畜疫病病原生物学国家重点实验室开放基金课题项目(No. SKLVEB2011KFKT007),石河子大学高层次人才科研启动项目(No. RCZX201104)联合资助
