副猪嗜血杆菌RfaF 基因的原核表达及抗原性分析
2013-08-14冯晓辉余远迪任玉鹏
冯晓辉,余远迪,任玉鹏,岳 华,张 斌
(西南民族大学生命科学与技术学院,四川成都610041)
副猪嗜血杆菌(Haemophilus parasuis)是一种不能运动的、革兰阴性短小杆菌,能引起以纤维素性多发性浆膜炎、关节炎和脑膜炎为主要特征的Glässer′s病[1]。H.parasuis引起的感染给养猪业带来了巨大的损失。革兰阴性菌的脂多糖(LPS)在其致病性中发挥重要的作用[2-5]。Bouchet B等[6]发现H.parasuis LPS部分参与了细菌对猪脑微血管内皮细胞和猪呼吸道上皮细胞的黏附作用和诱导IL-6和IL-8释放[6-7]。LPS是革兰阴性菌细胞壁的成分,由类脂A(Lipid A)、核心多糖和O多糖侧链组成。Lipid A为构成内毒素活性的糖质,以共价键链接在核心多糖上。而核心多糖是研究LPS功能和结构的重要部分。庚糖基转移酶是核心多糖重要的组成成分。庚糖基转移酶Ⅰ(HepⅠ)、庚糖基转移酶Ⅱ(HepⅡ)和庚糖基转移酶Ⅲ(HepⅢ)依次连接到Lipid A和3-脱氧-D-甘露-辛酮糖酸(Kdo)上。在革兰阴性菌中,庚糖基的缺失使得LPS的糖链严重的缩短,对血清中补体、抗菌肽和有机溶剂的敏感性上升,对宿主的致病性、黏附入侵能力等生物学特性发生变化。RfaF基因编码HepⅡ,该蛋白在细菌LPS的合成中是必不可少的。研究证实H.parasuisΔRfaF缺失株降低了对宿主细胞黏附入侵作用和抗血清中补体杀菌作用[8]。为了进一步研究H.parasuis RfaF的特性,本研究用大肠埃希菌表达系统对RfaF基因进行克隆和表达,为深入研究RfaF基因的功能和LPS在H.parasuis菌中的作用奠定了基础。
1 材料与方法
1.1 材料
1.1.1 载体和菌株 H.parasuis SC096株,由华南农业大学廖明教授馈赠;E.coil DH5α和BL21(DE3)工程菌,pET-32a(+)表达载体,由西南民族大学动物医学教研室保存。
1.1.2 主要试剂 TSA(胰蛋白胨大豆琼脂),LB固体培养基和LB液体培养基,购自青岛海博生物技术有限公司;烟酰胺腺嘌呤二核苷酸(NAD),购自Sigma公司;新生牛血清(NCS),购自浙江天杭生物技术有限公司;氨苄青霉素(AMP)、IPTG、T4连接酶,r Taq酶,BamHⅠ限制性内切酶,XhoⅠ限制性内切酶,Maker DL 2 000,Maker DL15 000,DNA胶回收试剂盒,DNA片段纯化试剂盒,小量质粒提取试剂盒和预染蛋白Marker,购自宝生物工程(大连)有限公司;辣根过氧化物酶(HRP)标记羊抗兔IgG(二抗)购自北京中杉金桥生物技术有限公司;兔抗4型副猪嗜血杆菌阳性血清(一抗)由本实验室制备并保存。
1.2 方法
1.2.1 引物设计 根据GenBank中H.parasuisSH0165菌株(№CP001321)的RfaF基因序列设计一对引物。引物如下:P1:5′-CGCGGATCCATGAATATTTTAGTTATTGC -3′, P2: 5′-CCGCTCGAGTTAAAGTGAACTTTTTTTTGA-3′(下划线部分分别为BamHⅠ和XhoⅠ酶切位点)。
1.2.2 RfaF基因的扩增 以SC096菌株DNA为模板,用P1/P2引物扩增目的片段。反应条件为:94℃3min;94℃30s,50℃40s,72℃1min,30个循环;72℃10min;16℃1h。PCR产物用胶回收纯化试剂盒进行纯化回收。
1.2.3 RfaF基因的克隆和序列测定 将纯化的PCR产物和pET-32a(+)载体用T4连接酶连接,转化至E.coliDH5α感受态细胞中,挑取单菌落进行PCR鉴定。阳性菌落接种于含AMP(100 μg/mL)的LB液体培养基中过夜,用小量质粒提取试剂盒提取质粒,BamHⅠ和XhoⅠ双酶切鉴定。
1.2.4 重组表达载体的构建 测序正确后,命名该重组质粒为pET-32a(+)-RfaF。将重组质粒转化至BL21(DE3)感受态细胞中。挑取单个菌落并进行PCR鉴定。
1.2.5 重组质粒的诱导表达及SDS-PAGE分析 将含有pET-32a(+)-RfaF和pET-32a(+)空载体的E.coilBL21(DE3)工程菌分别转接于含 AmpP(100 μg/mL)的LB培养基中,200r/min振摇培养过夜,次日按1∶100转接于含Amp(100μg/mL)的LB培养基中,37℃振摇培养至对数生长期(OD 600nm≈0.6),加入终浓度为1mmol/L的IPTG,分别于诱导后0、2、4、6、12h取菌1mL。菌液经12 000r/min离心1min后弃上清,沉淀用100μL pH 7.3的PBS重悬后,煮沸变性5min,10 000r/min离心1min取上清,加入上样缓冲液进行SDS-PAGE分析。
1.2.6 表达产物的可溶性分析 将优化诱导表达条件后的菌液12 000r/min离心1min后弃上清,沉淀用pH7.3的PBS重悬后用超声波粉碎仪粉碎30min(工作5s,间隔9s),在4℃以100 00r/min离心15min。分离上清和沉淀,沉淀用pH7.3的PBS重悬,煮沸变性5min,分别取变性后的上清液和重悬后的沉淀,加入上样缓冲液进行SDS-PAGE分析。
1.2.7 表达蛋白的Western blot分析 将表达的蛋白进行SDS-PAGE分析,然后转印到PVDF膜上,再转移至封闭液中过夜,4℃过夜后,室温下轻摇2h,用TBST洗膜3次,每次5min。再加入1∶200稀释的兔抗血清4型H.parasuis阳性血清(一抗),室温下轻摇2h。然后用TBST洗膜3次,每次10min。加入1∶3 000稀释的HRP标记的羊抗兔IgG(二抗),室温孵育2h,洗涤方法同上。最后在Versa Doc IMAGING SYSTEM下加显影液照相观察。
2 结果
2.1 RfaF基因的PCR扩增
经PCR扩增的RfaF基因片段经10g/L的琼脂糖凝胶电泳,片段大小约为1 000bp,与预期结果相符(图1)。
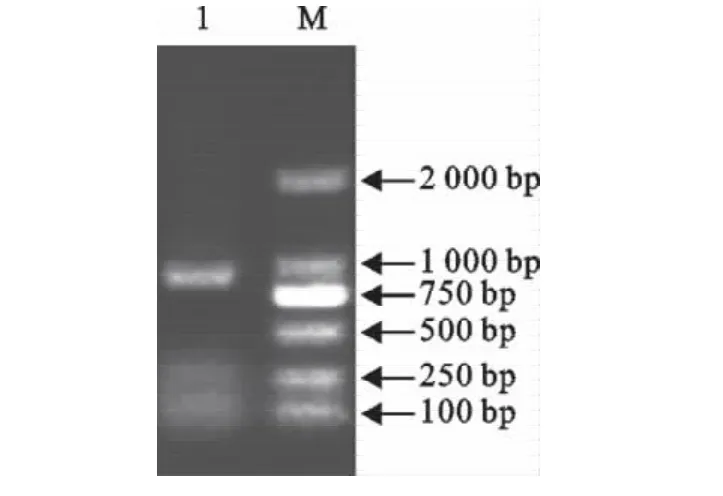
图1 RfaF基因的PCR扩增Fig.1 PCR amplification of RfaF gene
2.2 重组质粒的双酶切鉴定和PCR鉴定
将目的片段克隆到表达载体pET-32a(+)中,用BamHⅠ和XhoⅠ双酶切重组质粒,得到一条1 000bp左右的片段和一条5 900bp的片段。结果表明重组质粒与预期结果相符(图2)。重组质粒转化DH5α后,挑取单菌落经PCR扩增得到一条1 000bp左右的片段,与预期结果一致。
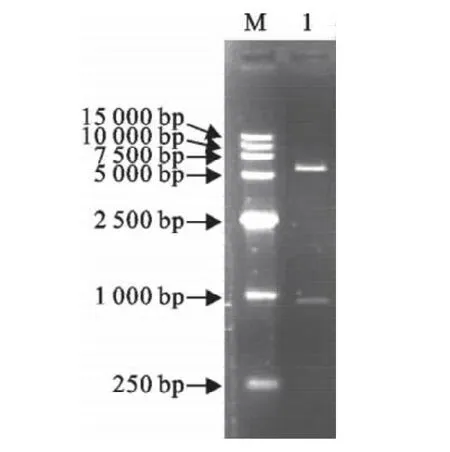
图2 重组质粒双酶切鉴定Fig.2 Identification of recombinant plasmid by enzyme digestion
2.3 RfaF基因的序列测定
将阳性重组质粒pET-32a(+)-RfaF进行序列测定。测定结果表明RfaF基因全长1 053bp,与血清5型分离株SH0165的同源性为96%(GenBank accession№NC-011852),说明目的基因已克隆于表达载体中。
2.4 重组菌的诱导表达及SDS-PAGE分析
经试验确定1.0mmol/L的IPTG诱导4h时表达量最大。表达产物约为55ku,与预期蛋白条带大小相近,而未诱导菌和诱导后的空载体均未出现此蛋白条带(图3)。
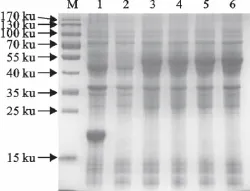
图3 重组蛋白的诱导表达Fig.3 Expression of recombinant protein induced by IPTG in E.coli BL21(DE3)
2.5 重组蛋白的可溶性分析
对超声波裂解细菌的上清和沉淀进行SDS-PAGE分析,表明重组蛋白以包涵体的形式存在(图4)。
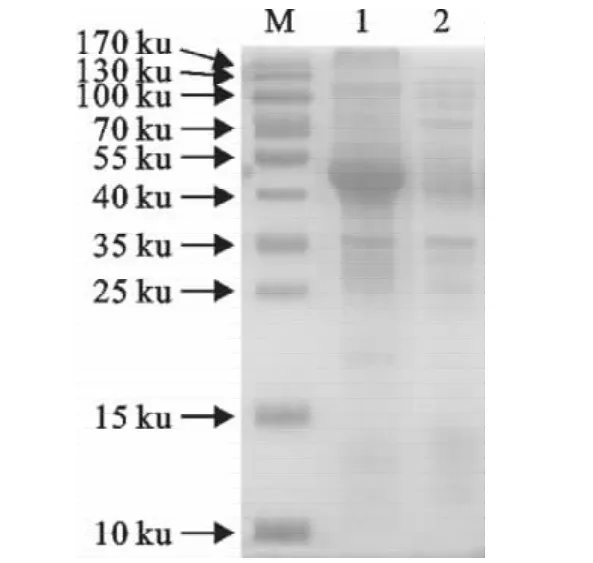
图4 重组蛋白的可溶性分析Fig.4 Solubility analysis of the recombinant protein
2.6 RfaF蛋白的Western blot分析
用兔抗副猪嗜血杆菌4型阳性血清(一抗)和HRP标记的羊抗兔IgG(二抗)对未纯化的包涵体蛋白进行Western blot分析,结果表明原核表达的RfaF重组蛋白能与副猪嗜血杆菌4型阳性血清发生特异性结合(图5中箭头所指的条带),RfaF重组蛋白具有良好的抗原性。
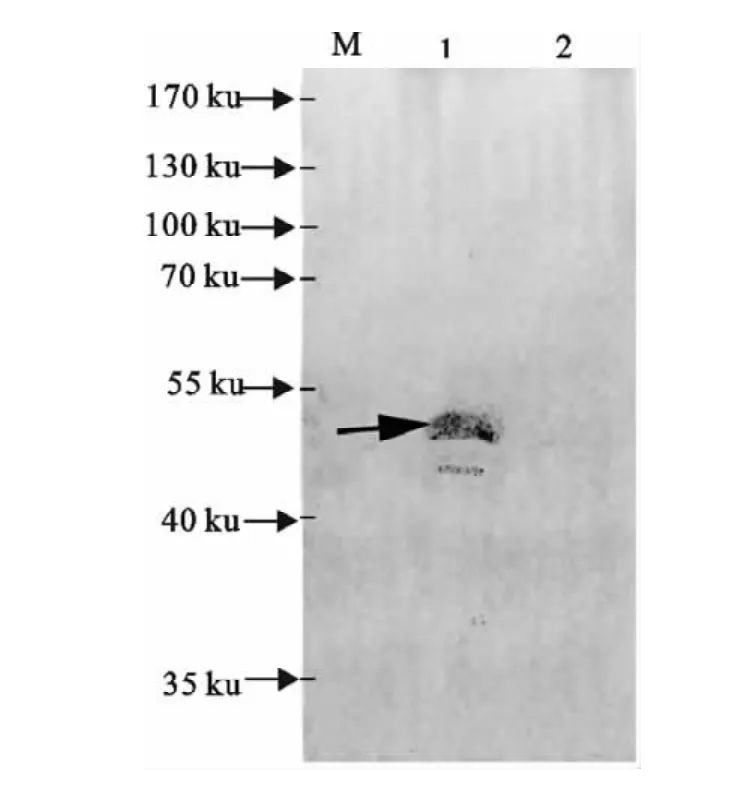
图5 RfaF蛋白的抗原性Western blot分析Fig.5 Antigenicity analysis of RfaF protein by Western blot
3 讨论
当前国内对H.parasuis的致病特性还了解的很少。已经有报道表明脂多糖在革兰阴性菌的致病中发挥着重要的作用[2-5],H.parasuisLPS可能在致病中发挥着重要的作用[9]。在革兰阴性菌中,LPS内部核心结构通常包括3-脱氧-D-甘露-辛酮糖酸(Kdo)和 L-乙酰-D-甘露庚糖(L,D-Hep)[10]。影响Kdo的合成和将Kdo转移到类脂A上的缺失株是致死的,Klena J D等[11]研究表明庚糖基转移酶Ⅱ的缺失是能实现的,并能够使LPS结构有明显的缩短。为进一步研究庚糖在H.parasuis致病性中发挥的作用,Xu C等[12]构建了RfaF基因缺失菌株和互补株。结果表明,RfaF基因缺失株的LPS结构明显的缩短,对血清的抗性有明显的下降,对RfaF基因缺失株的黏附入侵能力的研究表明缺失株对PUVEC和PK15细胞的黏附入侵能力显著降低,其互补株的黏附入侵能力恢复到了野生型水平。
本研究扩增并克隆了H.parasuisRfaF基因序列,结果表明,构建的pET-32a(+)-RfaF/BL21(DE3)系统在IPTG诱导下成功表达了RfaF蛋白,并对表达的重组蛋白进行了抗原性分析,表明RfaF重组蛋白具有良好的抗原性。本研究结果为进一步研究RfaF和脂多糖在H.parasuis致病性中的作用奠定了基础。
[1] Amano H,Shibata M,Kajio N,et al.Pathologic observations of pigs intranasally inoculated with serovar 1,4and 5of Haemophilus parasuis using immunoperoxidase method[J].J Vet Med Sci,1994,56(4):639-644.
[2] Swords W E,Buscher B A,Ver Steeg Ii K,et al.Non-typeable Haemophilus influenzae adhere to and invade human bronchial epithelial cells via an interaction of lipooligosaccharide with the PAF receptor[J].Mol Microbiol,2000,37(1):13-27.
[3] Kanipes M I,Holder L C,Corcoran A T,et al.A deep-rough mutant of Campylobacter jejuni 81-176is noninvasive for intestinal epithelial cells[J].Infect Immun,2004,72(4):2452-2455.
[4] Erwin A L,Allen S,Ho D K,et al.Role of lgtC in resistance of nontypeable Haemophilus influenzaestrain R2866to human serum [J].Infect Immun,2006,74(11):6226-6235.
[5] Ho D K,Ram S,Nelson K L,et al.lgtC expression modulates resistance to C4bdeposition on an invasive nontypeable Haemophilus influenzae[J].J Immunol,2007,178(2):1002-1012.
[6] Bouchet B,Vanier G,Jacques M,et al.Interactions of Haemophilus parasuis and its LOS with porcine brain microvascular endothelial cells[J].Vet Res,2008,39(5):42.
[7] Bouchet B,Vanier G,Jacques M,et al.Studies on the interactions of Haemophilus parasuis with porcine epithelial tracheal cells:limited role of LOS in apoptosis and pro-inflammatory cytokine release[J].Microb Pathog,2009,46(2):108-113.
[8] Xu C,Zhang L,Zhang B,et al.Involvement of lipooligosaccharide heptose residues of Haemophilus parasuis SC096strain in serum resistance,adhesion and invasion [J].Vet J,2013,195(2):200-204
[9] Tadjine M,Mittal K R,Bourdon S,et al.Production and characterization of murine monoclonal antibodies against Haemophilus parasuis and study of their protective role in mice[J].Microbiology,2004,150:3935-3945.
[10] Raetz C R,Whitfield C.Lipopolysaccharide endotoxins[J].Annu Rev Biochem,2002,71:635-700.
[11] Klena J D,Gray S A,Konkel M E.Cloning,sequencing,and characterization of the lipopolysaccharide biosynthetic enzyme heptosyltransferase I gene(waaC)fromCampylobacter jejuni and Campylobacter coli[J].Gene,1998,222(2):177-185.
[12] Xu Z,Yue M,Zhou R,et al.Genomic characterization of Haemophilus parasuis SH0165,a highly virulent strain of serovar 5prevalent in China [J].PLoS One,2011,6 (5):e19631.
