口蹄疫核酸疫苗研究进展*
2010-08-21张永光
王 刚,潘 丽,张永光
口蹄疫核酸疫苗研究进展*
王 刚,潘 丽,张永光
口蹄疫(Foot-and-mouth-disease,FMD)是由口蹄疫病毒(Foot-and-mouth-disease virus,FMDV)引起的危害严重的家畜传染病之一。由于传统疫苗的安全性、免疫保护期短、多次免疫、血清型间无交叉保护效应,感染与免疫难以区分等缺点。因而,寻求新的疫苗显得很有必要,而蛋白、多肽、弱毒苗、核酸疫苗、可饲疫苗等都被广泛研究。其中,核酸疫苗有许多优点,如生产工艺简单、可以长期保存、可以表达多种抗原基因、用于标记疫苗。口蹄疫核酸疫苗被大量研究,并取得了不同程度的成功。
1 口蹄疫核酸疫苗的抗原形式
口蹄疫核酸疫苗一般在质粒载体中连入VP1基因、P12A3C基因、B细胞表位和T细胞表位。有大量文献报到,以鸡痘病毒〔1〕、痘病毒〔2〕、牛痘〔3〕、腺病毒〔4-5〕、质粒〔6〕为载体共表达 P12A 和 3C,相对与P1、VP1,可以通过表达非结构蛋白3C,加工结构蛋白P1形成病毒空衣壳,因而其免疫原性优于P1、VP1、细胞表位。有研究表明,空衣壳可以引起相对强的免疫反应,但是也有不足,如L蛋白干扰5'帽子依赖细胞内mRNA翻译,3C阻止细胞内mRNA转录。Qingxia Yao等比较了P12A3C和细胞表位的免疫原性,结果表明以质粒的形式免疫豚鼠能引起相同的细胞免疫反应,但P12A3C能引起更高的体液免疫反应〔7〕。
非结构蛋白的作用机理的阐明,也为口蹄疫核酸疫苗抗原的研究提供了新的思路。非结构蛋白2B加入编码口蹄疫空衣壳蛋白的腺病毒载体(Ad5-CI-A24-2B),能引起相对Ad5-CI-A24更强的免疫反应和更高的保护率,这可能由于2B可以引起细胞膜重排和导致细胞死亡而使抗原蛋白释放。以编码2C蛋白或其片段的质粒免疫小鼠,确定其存在CTL表位,但由2C引起的CT L反应没有保护小鼠对病毒的攻击,因为免疫没有改变病毒血症和致死率〔8〕。以编码口蹄疫抗原P12A3C和 P12A3C3D的腺病毒免疫豚鼠,前者可以完全保护口蹄疫病毒攻击,而后者只有局部保护,这可能由于更长的开放阅读框导致蛋白表达量的减少〔9〕。有研究发现,结构蛋白更倾向于引起体液免疫反应,而非结构蛋白能够更有效的引起细胞免疫反应〔10〕。口蹄疫预防的细胞免疫反应的作用被逐渐认可,因而非结构蛋白的研究是很有必要的。
在郭慧琛等人的研究中,利用核糖体进入位点(IRES,internal ribosomal entry sites)来表达多顺反子,共表达了 P12X3C 和 IFN-α(pcDNA/PIF),其中2X包括部分2B和3B序列,结果表明pcDNA/PIF和共免疫pcDNA/P12X3C和pcDNA/IFN在特异性抗体、中和抗体、T细胞免疫水平上没有显著差异,因而双顺反子载体比两个质粒共免疫更方便〔11〕。也有报到以腺病毒为载体,借助IRES共表达了两个血清型的P12A基因和3C基因,并能够引起中和抗体〔5〕。编码口蹄疫VP1蛋白和猪细胞因子IFNα的多顺反子 rAd-pIFNα-VP1相对单顺反子(rAd-VP1+rAd-pIFNα)共免疫小鼠能显著增加细胞免疫和体液免疫反应〔12〕。
在细胞表位上连上一些定位分子基因,如泛毒蛋白、定位溶酶体信号蛋白、CTLA4、信号肽。其中泛毒蛋白、定位溶酶体信号蛋白分别定位于蛋白酶体和溶酶体,但起到副作用,可能因为在表位上存在一些酶切位点,从而被降解,因而在抗原表位递程过程中,选择其它途径可能值得研究。CTLA4则定位于抗原递程细胞上的CT LA4细胞受体,也没有表现出增强抗体的作用。信号肽则在体外提高抗原分泌,有1/5的动物引起中和抗体,并且完全保护病毒攻击〔13〕。把口蹄疫VP1蛋白插入到乙型肝炎病毒(HBV)核心区的显性表位C形成嵌合的核心-VP1病毒样颗粒,并插入带有hCG-β前导肽的真核表达载体,转染Hela细胞,可以在细胞上清中通过扫描电镜观察到病毒样颗粒,说明能成功分泌到细胞外〔14〕。
猪IgG恒定区作为细胞表位载体,免疫小鼠和猪均可以增强免疫反应〔15〕。连有GM-CSF和细胞表位的腺病毒载体免疫小鼠,结果表明可以引起较高的细胞免疫反应〔16〕。有研究在没有分子佐剂的情况下,表达口蹄疫病毒细胞表位的核酸疫苗免疫小鼠,可以在没有特异性体液免疫反应的情况下有50%小鼠(6/12)保护病毒的攻击〔13〕。
一些特殊的载体也被用于口蹄疫核酸疫苗抗原。伪狂犬病毒糖蛋白B由于可以被吞噬和在细胞与细胞间的运输,有研究以其作为口蹄疫病毒细胞表位的载体,发现当细胞表位插入在伪狂犬病毒糖蛋白B的N端两个B细胞表位之间,引起平衡的细胞免疫和体液免疫反应,而当细胞表位插入在伪狂犬病毒糖蛋白B的C端的B细胞中间,可以增强中和抗体水平〔17〕。把口蹄疫VP1蛋白插入到乙型肝炎病毒(HBV)核心区的显性表位C形成嵌合的核心-VP1病毒样颗粒,以质粒形式免疫动物,相对传统的只表达VP1的DNA疫苗,可以引起更强的免疫反应,特别是中和抗体水平,可能是因为形成特殊的三级结构,从而有增强免疫的作用〔14〕。
用重组病毒表达口蹄疫病毒衣壳蛋白多肽(P1),自然宿主在没有中和抗体的情况下也能部分保护,而这种保护可能是由T细胞和细胞因子引起的〔18-19〕。而没有引起中和抗体的原因,可能是 P1只表达于细胞质,而不能被B细胞识别〔19〕。以西门利克森林病毒为载体表达 P1蛋白,相对以质粒pcDNA3.1+为载体表达P1蛋白免疫小鼠能引起相对更强的免疫反应,且可以引起明显的细胞凋亡,这可能是免疫增强的原因之一,可能还由于其自我复制原因〔20〕。这些研究都表明,P1作为抗原由于不能分泌到胞外从而不能有效引起中和抗体,然后加入3C能增加中和抗体水平。
在豚鼠的攻毒保护实验中,伪狂犬病毒编码口蹄疫P12A3C组能完全保护,而质粒编码口蹄疫P12A3C或细胞表位组则部分保护〔7〕。显示出活载体疫苗相对质粒为载体的核酸疫苗有更好的效果,但是由于活载体疫苗的病毒载体背景,限制了其广泛的应用,因而研究者还在研究更高效的质粒载体疫苗。表达口蹄疫病毒P12A3C的基于辛德毕斯病毒的非复制质粒免疫动物,并没有显示出相对以PcDNA3.1为载体的传统核酸疫苗有增强抗口蹄疫的特异性抗体的反应〔21〕。以有复制能力的犬腺病毒表达口蹄疫 VP1蛋白,可以引起体液免疫反应〔22〕。
由于核酸疫苗引起抗体水平低且慢,因而有研究,利用灭活疫苗和口蹄疫非结构蛋白3D来加强免疫,以质粒P1-2A3C3D和质粒 pGM-CSF初免猪,结果表明相对灭活疫苗有36倍的抗体反应,且能引起血清型间显著的交叉保护〔23〕。把核酸疫苗和灭活疫苗或合成肽疫苗的优点融合一起,这为口蹄疫预防提供了新的方向。
2 口蹄疫核酸疫苗佐剂
2.1 分子佐剂 多种细胞因子佐剂或功能蛋白的基因被用于口蹄疫核酸疫苗佐剂,大部分都被证明可以起到促进免疫反应的作用,但是效果存在差异,然后由于抗原形式、免疫动物、佐剂、免疫途径、载体、血清型等的差异,因而很难比较其免疫效果。以下就在口蹄疫核酸疫苗研究中,所使用的分子佐剂简要概括。
编码IL-15的质粒作为佐剂和编码口蹄疫抗原基因VP1的质粒共同免疫动物,结果表明相对于口蹄疫抗原基因VP1单免组能够显著增强细胞介导的免疫反应,及引起更高水平的口蹄疫特异性中和抗体水平和在黏膜部位大量分泌IgA〔24〕。
IL-18有促进Th1和Th2免疫的作用,以及可能促进抗体产生,并被认为能有效地诱导IFN-γ产生,有研究称IFN-γ对于口蹄疫的预防有很重要的作用。以DNA疫苗(pVIR-P12A-IL18-3C)共表达猪IL18来免疫猪,相对DNA疫苗(pVIR-P12A3C)可以引起更强的免疫反应和更高的保护率,表明IL18可以增强体液、细胞免疫反应〔6〕。
IFN-α不仅有抑制抗原的作用,还有免疫调节的作用,有研究表明IFN-α在细胞内有抑制口蹄疫的作用。rAd-pIFNα-VPe/rAd-pIFNα-VP1双顺反子免疫或 rAd-pIFNα与 rAd-pIFNα-VPe/rAdpIFNα-VP1单顺反子共免疫,都显示出相对不加IFN-α的VPe或VP1有更强的免疫反应,且双顺反子比单顺反子共免疫能引起显著的免疫增强反应,攻毒保护实验结果显示免疫 rAd-pIFNα-VPe或rAd-pIFNα-VP1豚鼠均能保护病毒攻击,免疫VP1的猪也能保护病毒攻击〔12,25〕。Ad5-A24(P12A3C)和Ad5-pIFNα共免疫也表明相对Ad5-A24能引起更强的免疫反应,猪在免疫后5天攻毒能完全保护病毒的攻击〔26-27〕。
质粒pGM-CSF与pcDNA 3.1/P1-2A3C3D共免疫猪可以引起更强的特异性抗体和中和抗体反应,以及增加细胞因子 IL-8和 IFN-γ的分泌〔28〕。rAd-GMCSF-VPe和 rAd-GMCSF-VP1能引起相对rAd-VPe和rAd-VP1更强的体液和细胞免疫反应,刺激分泌 Th1和Th2型细胞因子〔16〕。
质粒pCEIS(编码VP1细胞表位且以猪IgG恒定区作载体)和 pIL2S共免疫猪,相对质粒pCEIS单免能引起更高的T细胞增殖,而抗口蹄疫中和抗体并无显著差异〔15〕。以IRES构建多顺反子真表达载体pVAX-SG-IL-2,相对单顺反子共免疫组或双表位基因可以增强抗体反应和细胞增殖,及细胞因子分泌〔29〕。
IL-6和TNF-α以质粒形式与质粒pcD-VP1共免疫小鼠,均能引起高比例的IgG2a/IgG1,高表达IFN-γ和IL-4,及细胞毒作用,促使DC细胞成熟,特别是增强抗原特异的细胞免疫反应〔30〕。
rAd-GMCSF-VPe(VPe指VP1上的表位)引起更高的细胞免疫反应,而rAd-GMCSF-VP1引起更高的体液免疫反应,但中和抗体水平比灭活疫苗都低二倍以上,两者联合免疫可以完全保护口蹄疫病毒攻击〔16〕。
补体C3d可以与B细胞上CD21结合,从而起到促进抗原摄取的作用。VP1和C3d/C3d受体顺式连接到带有信号肽的真核表达载体连接,免疫豚鼠,能促进体液免疫和细胞免疫反应,sVP1-mC3d3组有87.5%的保护率〔31〕。
以B细胞激活因子(BAFF)与质粒pcDNA 3.1/P1-2A3C3D共免疫猪,结果显示BAFF并未增强抗体生成,因而不适合作为佐剂〔32〕。
2.2 化学佐剂 以化学佐剂左旋咪唑、(Tween 80)、布比卡因(bupivacaine)、乙醇、盐溶液等与pcD-VP1共免疫小鼠,(tPA ,tissue plasminogen activatorleader sequence,a signal sequence)
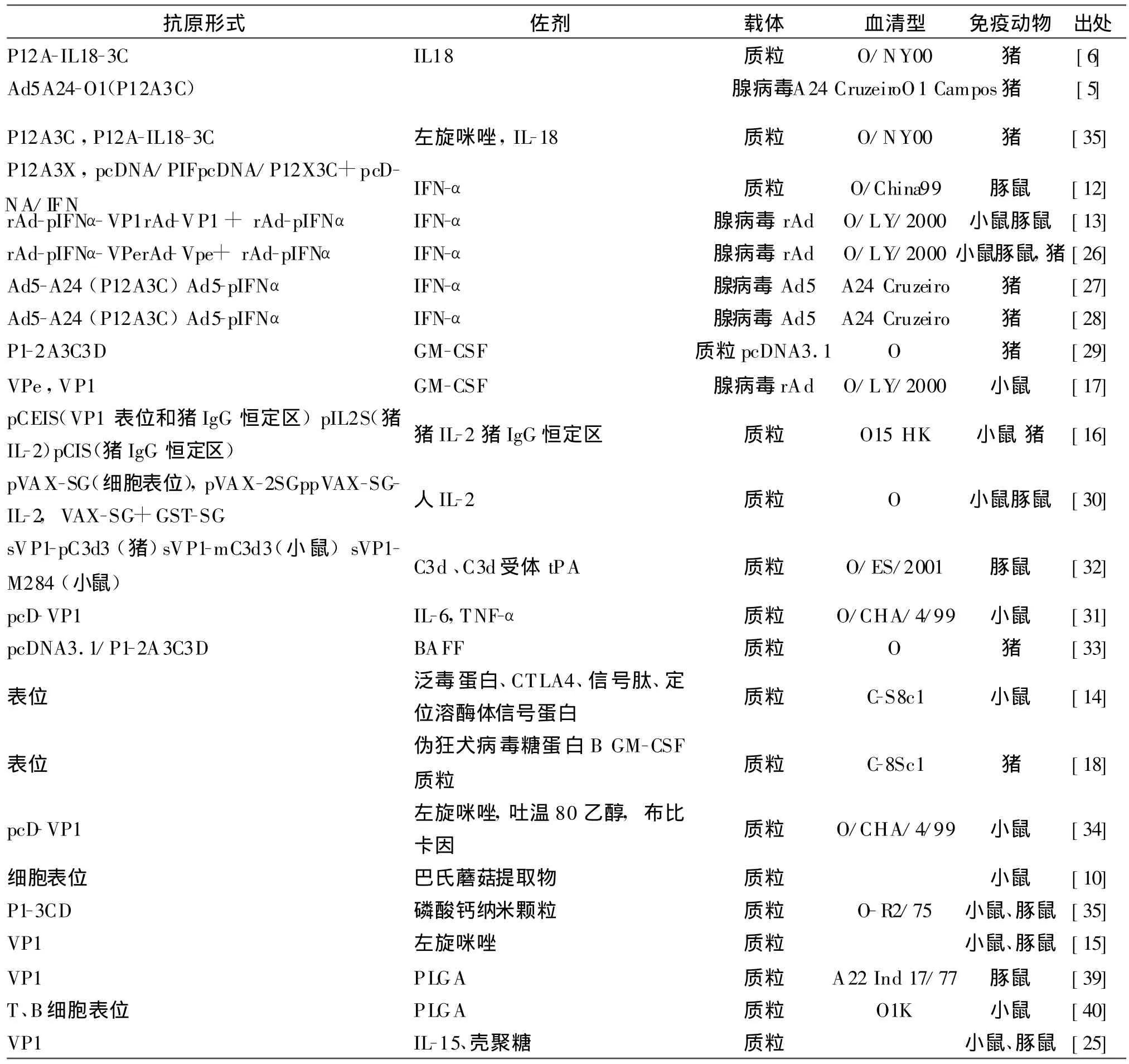
表1 有关口蹄疫核酸疫苗文献
结果表明左旋咪唑能引起相对最强的T h1反应,吐温80则显示出最弱的Th1反应,左旋咪唑可以引起相对盐溶液约100倍的IFN-γ分泌,布比卡因只引起中等水平的抗体,及相对低水平的Ig2a〔33〕。以左旋咪唑作为佐剂,核酸疫苗(pVIRP12A-IL18-3C)免疫猪,相对单免疫 pVIR-P12AIL18-3C,可以引起更强的细胞和体液免疫反应,也有更高的保护率;且引起显著的口蹄疫特异性的IFN-γ反应(P<0.001)〔34〕。巴氏蘑菇提取物富含多糖,口蹄疫核酸疫苗及以巴氏蘑菇提取物为佐剂免疫小鼠,相对仅以疫苗免疫组能增强抗体水平和T 细胞增殖〔35〕。
2.3 纳米佐剂 Hong-Ying Zhang等以编码口蹄疫表位的质粒以水溶液的形式注射动物,能快速吸收,广泛分布,但是利用率很低〔36〕。因而,口蹄疫核酸疫苗要有更高的利用率,就必须有适当的载体或佐剂,以保护质粒在体内不被快速降解。编码口蹄疫P1-3CD基因的核酸疫苗以磷酸钙纳米颗粒为载体,体外转染效率和商业的脂质体转染试剂相当,免疫小鼠和豚鼠可以增强细胞免疫和体液免疫,且能够保护病毒攻击。编码IL-15的质粒和口蹄疫抗原基因VP1的质粒以壳聚糖为载体,经鼻免疫小鼠,可以在黏膜组织分泌IgA抗体,和引起系统免疫反应〔24〕。有研究结果表明,编码口蹄疫 VP1和细胞表位的核酸疫苗和聚乳酸乙醇酸(PLGA)免疫动物,可以增强免疫反应〔37-38〕。
3 口蹄疫核酸疫苗的递送方式
核酸疫苗一般通过肌肉注射,皮下注射、鼻黏膜免疫,经口免疫。气雾传播被认为是口蹄疫传播的一个重要途径,因而诱导黏膜免疫被认为是有效的预防途径。而黏膜免疫分为经口、鼻、及肺黏膜免疫。利用壳聚糖来载口蹄疫抗原基因VP1和IL15基因由鼻黏膜免疫动物,可增强抗原特异性的黏膜和系统免疫反应,从而可能保护口蹄疫病毒在起始的感染〔24〕。口蹄疫核酸疫苗免疫猪,实验结果表明皮下注射相对肌肉注射能引起更高的抗体反应〔17〕。
4 讨 论
核酸疫苗研究由于具备生产工艺简单、可以长期保存、可以表达多种抗原基因、用于标记疫苗等优点,而受到广泛研究,又由于口蹄疫核酸疫苗的诸多缺点,口蹄疫核酸疫苗的研究便成为一种有前景的潜在的疫苗。在抗原形式上,取得了一定的突破,但仍然不能在免疫原性和抗原基因的长度上达到平衡,也就是说既保证抗原基因上有足够的细胞表位或片段来引起免疫反应,又不能加入太长的序列影响抗原基因的表达。另外,其它一些抗原形式被研究,如多顺反子比单顺反子共同免疫效果要好,细胞表位需要加入一些载体以提供一定的三维结构等。核酸疫苗的保护效果,特别是抗体水平与灭活疫苗所产生的抗体水平仍有很大的差距,为了增强免疫效果,一些细胞因子佐剂或分子佐剂被应用于核酸疫苗免疫,还有一些化学佐剂也被研究。对于分子佐剂,有 IL-1α,IL-1β,IL-2,IL-6,IL-15,IL-18,IFN-α,IFN-β,IFN-γ,B-cell activating factor,GMCSF,C3d,4-1BBL、OX40L,IgG重链恒定区等被研究,但是由于抗原形式、免疫动物、佐剂、免疫途径、载体、血清型等的差异,因而很难比较其免疫效果。
核酸疫苗的缺点,如插入基因组的可能性、免疫产生的抗体水平低从而保护率低。口蹄疫核酸疫苗要达到理想的效果,仍然需要研究抗原基因的作用机理,而使用合适的抗原形式;选择合适的载体保证核酸疫苗被动物有效摄取,并能到达相应的免疫器官;选择高效的表达载体,确保足够的抗原基因在体内表达;筛选有效的佐剂,从而增强免疫;从而引起高效的免疫应答。相信,口蹄疫核酸疫苗的研究仍然有很大的挑战,但也有广阔的前景。
〔1〕张洪勇,金宁,葛淑敏,等.共表达FMDV P12A 和3C重组鸡痘病毒疫苗与核酸疫苗的实验免疫研究〔J〕.高技术通讯,2004,8:(20-23).
〔2〕Abrams CC,King AM,Belsham GJ.Assembly of foot-andmouth disease virus empty capsids synthesized by a vaccinia virus expression system〔J〕.J Gen Virol,1995,76(12):3089.
〔3〕Mingxiao Ma,Ningyi Jin,Guoshun Shen,et al.Immune responses of swine inoculated with a recombinant fowlpox virus co-expressing P12A and 3C of FMDV and swine IL-18〔J〕.Veterinary Immunology and Immunopathology,2008,121(1-2):1-7.
〔4〕Moraes M P,Mayr G A,Mason P W,et al.Early protection against homologous challenge after a single dose of replication-defective human adenovirus type 5 expressing capsid proteins of foot-and-mouth disease virus(FMDV)strain A24〔J〕.2002,20:1631-1639.
〔5〕Qiaohua Wu,Moraes M P,G rubman M J.Recombinant adenovirus co-expressing capsid proteins of two seroty pes of foot-andmouth disease virus(FMDV):in vitro characterization and induction of neutralizing antibodies against FMDV in swine〔J〕.Virus Research,2003,93(2):211-219.
〔6〕Mingxiao M,Ningyi J,Juan LH,et al.Immunogenicity of plasmids encoding P12A and 3C of FMDV and swine IL-18〔J〕.Antiviral Res,2007,76(1):59-67.
〔7〕Yao Q,Qian P,Huang Q,et al.Comparison of immune responses to different foot-and-mouth disease genetically engineered vaccines in guinea pigs〔J〕.J Virol Methods,2008,147(1):143.
〔8〕Barfoed AM,Rodriguez F,Therrien D,et al.DNA immunization with 2C FMDV non-structural protein reveals the presence of an immunodominant CD8+,CTL epitope for BALB/c mice〔J〕.Antiviral Res,2006,72(3):178.
〔9〕Lu Z,Bao H,Cao Y,et al.Protection of guinea pigs and swine by a recombinant adenovirus expressing O serotype of foot-andmouth disease virus whole capsid and 3C protease〔J〕.Vaccine,2008,26(6):48-53.
〔10〕Cedillo-Barrón L,Foster-Cuevas M,Belsham,et al.Induction of a protective response in swine vaccinated with DNA encoding foot-and-mouth disease virus empty capsid proteins and the 3D RNA polymerase〔J〕.J Gen Virol,2001,82:1713-1724.
〔11〕Huichen GUO,Shiqi SUN,Jiangtao MA,et al.A Bicistronic DNA Vaccine Containing Gene of FMDV and Bovine IFN-αCan Prime Humoral and Cellular Immune Responses of Guinea Pigs Turk〔J〕.J Vet Anim Sci,2008,32(2):85-91.
〔12〕Du Y,Dai J,Li Y,et al.Immune responses of recombinant adenovirus co-expressing VP1 of foot-and-mouth disease virus and porcine interferon alpha in mice and guinea pigs〔J〕 .Vet Immunol Immunopathol,2008,124(3-4):274.
〔13〕Borrego B,Fernandez-Pacheco P,Ganges L,et al.DNA vaccines expressing B and T cell epitopes can protect mice from FMDV infection in the absence of specific humoral responses〔J〕.Vaccine,2006,24(18):3889.
〔14〕Jin H,Xiao W,Xiao C,et al.Protective immune responses against foot-and-mouth disease virus by vaccination with a DNA vaccine ex pressing virus-like particles〔J〕.Viral Immunol,2007,20(3):429.
〔15〕Wong HT,Cheng SC,Sin FW,et al.DN A vaccine against foot-and-mouth disease elicits an immune response in swine which is enhanced by co-administration with interleukin-2〔J〕.Vaccine,2002,20(21-22):2641.
〔16〕Du Y,Jiang P,Li Y,et al.Immune responses of two recombinant adenoviruses expressing VP1 antigens of FMDV fused with porcine granulocy te macrophage colony-stimulating factor〔J〕.Vaccine,2007,25(49):8209-19.
〔17〕Dory D,Rémond M,Béven V,et al.Pseudorabies virus glycoprotein B can be used to carry foot and mouth disease antigens in DNA vaccination of pigs〔J〕.Antiviral Res,2009,81(3):217.
〔18〕Sanz-Parra A,Vazquez B,Sobrino F,et al.Evidence of partial protection against foot-and-mouth disease in cattle immunized with a recombinant adenovirus vector expressing the precursor polypeptide(P1)of foot-and-mouth disease virus capsid proteins〔J〕.J Gen Virol,1999,80(3):671.
〔19〕Sanz-Parra A,Jimenez-Clavero MA,Garcia-Briones MM,et al.Recombinant viruses expressing the foot-andmouth disease virus capsid precursor polypeptide(P1)induce cellular but not humoral antiviral immunity and partial protection in pigs〔J〕.Virology,1999,259(1):129.
〔20〕 Yu X,Xiao S,Fang L,et al.Enhanced immunogenicity to food-and-mouth disease virus in mice vaccination with alphaviral replicon-based DNA vaccine expressing the capsid precursor polypeptide(P1)〔J〕.1:Virus Genes,2006,33(3):337.
〔21〕Dory D,R mond M,B ven V,et al.Foot-and-Mouth Disease Virus neutralizing antibodies production induced by pcDNA3 and Sindbis virus based plasmid encoding FMDV P1-2A3C3D in swine〔J〕.Antiviral Res,2009,83(1):45-52.
〔22〕Liu Y,Hu R,Zhang S,et al.Expression of the Foot-and-M outh Disease Virus VP1 protein using a replication-competent recombinant canine adenovirus ty pe 2 elicits a humoral antibody response in a porcine model〔J〕.Viral Immunol,2006,19(2):202.
〔23〕Li Y,Stirling CM,Denyer MS,et al.Dramatic improvement in FMD DNA vaccine efficacy and cross-serotype antibody induction in pigs following a protein boost〔J〕.Vaccine,2008,26(21):2647.
〔24〕 Wang X,Zhang X,Kang Y,et al.Interleukin-15 enhance DNA vaccine elicited mucosal and systemic immunity against foot and mouth disease virus〔J〕.Vaccine,2008,26(40):5135.〔25〕Du Y,Li Y,He H,et al.Enhanced immunogenicity of multiple-epitopes of foot-and-mouth disease virus fused with porcine interferon alpha in mice and protective efficacy in guinea pigs and swine〔J〕 .J Virol Methods,2008,149(1):144.
〔26〕Mo raes MP,Chinsangaram J,Brum MC,et al.Immediate protection of swine from foot-and-mouth disease:a combination of adenoviruses expressing interferon alpha and a foot-and-mouth disease virus subunit vaccine〔J〕.Vaccine,2003,22(2):268.
〔27〕de Avila Botton S,Brum MC,Bautista E,et al.Immunopotentiation of a foot-and-mouth disease virus subunit vaccine by interferon alpha〔J〕.Vaccine,2006,24(17):3446-56.
〔28〕Li Y,Aggarwal N,Takamatsu HH,et al.Enhancing immune responses against a plasmid DNA vaccine encoding a FMDV empty capsid from serotype O〔J〕.Vaccine,2006,24(21):4602.
〔29〕Zhang HY,Sun SH,Guo YJ,et al.Optimization strategy for plasmid DNAs containing multiple-epitopes of foot-and-mouth disease virus by cis-ex pression with IL-2〔J〕.Vaccine,2008,26(6):769.
〔30〕Su B,Wang J,Wang X,et al.The effects of IL-6 and TNF-alpha as molecular adjuvants on immune responses to FM DV and maturation of dendritic cells by DNA vaccination〔J〕.Vaccine,2008,19;26(40):5111.
〔31〕Fan H,Tong T,Chen H,et al.Immunization of DNA vaccine encoding C3d-VP1 fusion enhanced protective immune response against foot-and-mouth disease virus〔J〕.Virus Genes,2007,35(2):347.
〔32〕Bergamin F,Saurer L,Neuhaus V,et al.Porcine B-cell activating factor promotes anti-FMDV antibodies in vitro but not in vivo after DNA vaccination of pigs〔 J〕 .Vet Immunol Immunopathol,2007,120(3-4):115.
〔33〕Jin H,Li Y,Ma Z,et al.Effect of chemical adjuvants on DNA vaccination〔J〕.Vaccine,2004,22(21-22):2925-35.
〔34〕Lu H,Huo X,Zhang Y,et al.Enhancing effects of the chemical adjuvant levamisole on the DNA vaccine pVIR-P12A-IL18-3C〔J〕.Microbiol Immunol,2008,52(9):440.
〔35〕Chen L,Shao H.Extract from Agaricus blazei Murill can enhance immune responses elicited by DNA vaccine against footand-mouth disease〔J〕.Vet Immunol Immunopathol,2006,109(1-2):177.
〔36〕Zhang HY,Sun SH,Guo YJ,et al.Tissue distribution of a plasmid DNA containing epitopes of foot-and-mouth disease virus in mice〔J〕.Vaccine,2005,23(48-49).
〔37〕Choudary S,Ravikumar P,A shok Kumar C,et al.Enhanced immune response of DNA vaccine(VP1-pCDNA)adsorbed on cationic PLG for foot and mouth disease in guinea pigs〔J〕.Virus Genes,2008,37(1):81.
〔38〕Wang F,He XW,Jiang L,et al.Enhanced immunogenicity of microencapsulated multiepitope DNA vaccine encoding T and B cell epitopes of foot-and-mouth disease virus in mice〔J〕.Vaccine,2006,24(12):2017-26.
R392.1
A
1002-2694(2010)09-0874-05
*欧盟项目FMD-DISCONVAC(G rant No.226556)
张永光,Email:zhangyg@public.lz.ys.cn
中国农业科学院兰州兽医研究所,家畜疫病病原生物学国家重点实验室,农业部畜禽病毒学重点开放实验室,甘肃 兰州 730046
2009-09-28;
2010-01-04
