金银花香豆酸3-羟化酶基因LjC3H2 的生物信息学分析
2024-02-05冯唐锴罗丽萍
冯唐锴 雷 群 罗丽萍 蓝 玉
(1景德镇学院生物与环境工程学院,江西景德镇 333499;2景德镇市第十二小学,江西景德镇 333000)
金银花在抗菌消炎和对抗病毒性疾病方面有一定作用,常用于中药方剂中。绿原酸(Chlorogenic acid,CGA)是金银花重要的药用成分之一,具有多种药用功效[1]。植物绿原酸合成途径有3 条:一是咖啡酰辅酶A 和奎宁酸由羟基化肉桂酸辅酶A或奎宁酸肉桂酸羟基化转移酶催化生成绿原酸;二是在羟基肉桂酰基转移酶等作用下生成p-香豆酰奎宁酸,再由p-香豆酸3-羟化酶(p-coumarate3-hydroxylase,C3H)羟基化生成绿原酸;三是咖啡酰葡萄糖苷作为中间产物,和奎宁酸在羟基化肉桂酰D-葡萄糖、奎宁酸羟基化肉桂酰转移酶催化作用下生成绿原酸。C3H 是绿原酸合成代谢过程中的关键酶,该酶催化咖啡酸的生物合成,这个过程是合成绿原酸的重要步骤。C3H 属于细胞色素P450家族,与各类植物的CYP450 98A 亚家族蛋白具有高度同源性。在植物苯丙素代谢中能催化苯环C3位置的羟基化反应,是绿原酸生物合成的关键酶之一。金银花基因组中有两个C3H拷贝,2013年第一个C3H 基因拷贝(LjC3H)被克隆,亓希武等[2]对另外一个C3H 基因拷贝(LjC3H2)进行了研究。其他植物C3H 也有相关研究,王漫青等[3]的研究表明,当归C3H 基因开放阅读框长1 530 bp,编码509 个氨基酸。周美娟等[4]克隆了慈竹C3H 基因并进行了分析。李高等[5]的研究表明柠条锦鸡儿C3H 与拟南芥C3H 具有部分相同的功能。本文从各类数据库收集了37 种植物40 个C3H 蛋白序列,利用多种生物信息学分析工具,以LjC3H2为研究对象,对该基因表达酶的结构、功能和分类等特点进行了进一步的分析研究。
1 材料与方法
1.1 数据材料
LjC3HL2酶蛋白在NCBI 蛋白质数据库中的序号为ART33341.1。以该蛋白氨基酸序列为样本进行Blast 比对,在NCBI 蛋白质数据库找到37 种植物40个C3H蛋白氨基酸序列,用来对LjC3H2进行分子进化分析。相关植物蛋白序列号如下:金银花(ART33341.1、AGQ48118)、桔梗(AEM63674.1)、夏威夷金合欢(AOX49220.1)、西桦(QGX89943.1)、拧条锦鸡儿(AEV93473.1)、蓟(QQH14908.1)、矮牵牛(AVA30528.1)、毛白杨(AFZ78540.1、APR63682.1)、中粒咖啡(ABB83677.1)、苦荞麦(AHA14499.1)、芝麻(AAL47545.1)、黄水仙(AUG71937.1)、蓝星睡莲(XP_031473498.1)、鲍氏文殊兰(WGU11329.1)、粗壮女贞(WEX29781.1)、红花钓钟柳(WEX29779.1)、沙蓬(WBR79877.1)、茄子(UXQ89629.1)、蓖麻(XP_002511566.3)、圆叶黄山药(XP_039114632.1)、龙葵(QIC52992.1)、缸豆(QCD97092.1)、桃儿七(AIA24412.1) 、水 仙(AGI97941.1) 、 慈 竹(AFD29885.1)、杉木(AFX98060.1)、火炬松(AAL47685.1)、欧洲云杉(CAK22403.1)、地黄(QJS39421.1)、箭叶淫羊藿(AIS92508.1)、陆地棉(ALH21661.1)、南非醉茄(ADM47799.1)、大麻槿(AGA60530.1)、药蜀葵(UOI87846.1)、巨芒草(ANB43566.1、ANB43567.1)、黄芩(BAJ09387.1)和辣椒(ACF17644.1)。这40 个植物C3H 蛋白中,除了沙蓬和欧洲云杉的酶蛋白序列不够完整以外,其他植物的酶蛋白氨基酸数都在510 左右,最少的为503(杉木),最多的为517(西桦、矮牵牛和茄子)。
1.2 生物信息学工具和分析方法
使用的生物信息学工具包括各类在线数据库、在线数据分析网站以及离线分析软件,具体名称、网址及功能如表1所示。

表1 使用的生物信息学分析软件和在线程序
美国国家生物信息中心(NCBI)可用于获得、提交和分析各类生物信息数据。BLAST是其中的一个分析工具,可对不同氨基酸或核酸序列进行同源比较。DNAMAN 是一款高度集成化的序列分析软件,本文利用该软件进行金银花C3H蛋白的辅助序列处理和分析。MEGA 是对物种和种群DNA 和蛋白质序列进行分子进化遗传分析的软件套装,本文利用该软件对不同植物C3H蛋白氨基酸序列进行比较和进化树的构建。ProtParam 是对蛋白质进行理化性质分析的软件,可计算蛋白质的各种物化参数,包括分子量、理论pI 和氨基酸组成等。ProtScale 是对蛋白质进行亲疏水性分析的在线软件。SignalP 6.0能够预测信号肽的存在及其在蛋白质中的切割位点的位置[6]。TMHMM 是对蛋白质进行跨膜螺旋区分析的服务器,用隐马尔可夫模型预测跨膜蛋白拓扑结构[7]。Euk-mPLOC是真核蛋白质亚细胞定位分析工具,包括具有多个亚细胞位点的真核蛋白质也能预测[8]。SOPMA 是一种蛋白质二级结构预测方法,分析蛋白质中α-螺旋、β-折叠和无规卷曲等主要二级结构的存在情况[9]。SWISS-MODEL 得到了Alpha-Fold 蛋白质结构数据库等资源工具的支撑,能高效精准的自动对蛋白质结构进行同源性建模[10]。Prot-Param、ProtScale、SignalP6.0、TMHMM、Euk-mPLOC、SOPMA和SWISS-MODEL这些在线程序都有功能强大,简单易用的特点。对该基因的生物信息学进行分析,将下载好的LjC3H2氨基酸序列导入其输入界面中,再点击提交即可得到相关蛋白信号肽、跨膜结构域、亚细胞定位以及蛋白质二级和三级空间结构的预测分析结果。
2 结果与分析
2.1 LjC3H2编码蛋白的理化性质
通过程序EXPASY 的ProtParam 分析金银花LjC3H2编码蛋白的理化性质。结果显示,LjC3H2编码蛋白质原子总数8 133,分子式为C2601H4083N715O715S19;分子量57 419.53;等电点pI 为8.92;序列的N 末端是M(Met)时估计半衰期为30 h(哺乳动物网织红细胞)>20 h(酵母)>10 h(大肠杆菌)。不稳定性指数计算为38.41,即该蛋白质应归类为稳定蛋白质。脂肪指数92.33,亲水性总平均值(重力)-0.231,偏亲水性蛋白。LjC3H2蛋白共含氨基酸残基506 个,包括蛋白质中常见的20 种氨基酸,其中亮氨酸(Leu,11.7%),丙氨酸(Ala,8.3%),精氨酸(Arg,6.9%),缬氨酸(Val,6.9%),脯胺酸(Pro,6.9%),谷氨酸(Glu,6.3%),甘氨酸(Gly,6.1%),赖氨酸(Lys,5.7%),天冬氨酸(Asp,5.1%)含量相对较多,半胱氨酸(Cys,0.8%)最少,带负电荷的残基总数(天冬氨酸+谷氨酸)为58,带正电的残基总数(精氨酸+赖氨酸)为64(表2)。

表2 LjC3H2蛋白质的氨基酸组成
预测LjC3H2在水中280 nm处的消光系数,若假设所有的半胱氨酸残基都形成胱氨酸,则消光系数为80 120,吸光度值(0.1%浓度)为1.395。假设所有的半胱氨酸残基都不形成胱氨酸,则消光系数为79 870,吸光度值(0.1%浓度)为1.391。用DNAMAN软件对LjC3H2做胰蛋白酶的酶切分析,结果表明胰蛋白酶对LjC3H2做酶切分析可得39 个片段。用DNAMNA 软件预测LjC3H2抗原位点的结果表明,在LjC3H2中存在的抗原位点有8个(表3)。

表3 LjC3H2抗原位点预测
用ProtScale 软件对LjC3H2做亲水性和疏水性预测分析,得到的结果如图1 所示。其中以LjC3H2中的氨基酸(aa)序列号作横坐标,亲水性分值作为纵坐标,负值越高表示亲水性越强,正值越高表示疏水性越强。结果表明,除了起始的少数几个氨基酸疏水性较高以外,整个蛋白质没有特别明显的“峰顶”和“谷底”,亲/疏水性相对均匀,整体来看亲水性略高。
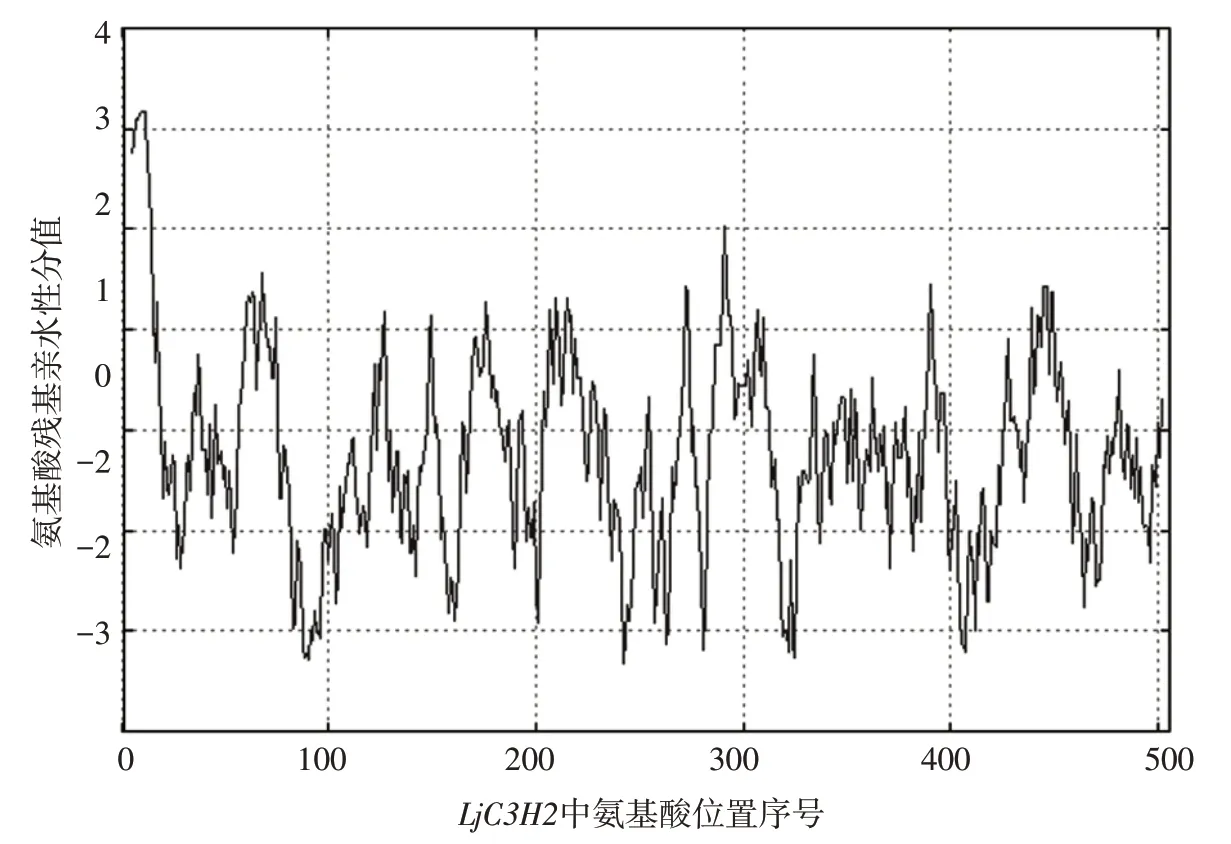
图1 LjC3H2亲水性分析
2.2 LjC3H2蛋白的信号肽情况
用DNAMAN 软件对LjC3H2进行信号肽预测分析,结果表明该蛋白自然分裂位点CS 的峰值为0.134 778,在第22 位;OTHER 的峰值为0.999 885,在第70 位;而信号肽峰值为0.262,无明显峰值,即LjC3H2无信号肽。
2.3 LjC3H2蛋白的跨膜结构情况
运用程序工具TMHMM 对LjC3H2蛋白质进行跨膜区分析。结果表明,该蛋白质氨基酸序列长度506,预测出的跨膜螺旋数量为0,跨膜螺旋氨基酸残基数量的期望值为4.674 93,蛋白前60 个氨基酸中,跨膜螺旋的氨基酸量的期望值为4.430 10,位于膜细胞质侧的总概率为0.221 44,预测结果表明LjC3H2没有跨膜结构域。通常蛋白质跨膜螺旋中的氨基酸残基多为疏水性氨基酸残基,亲/疏水性分析结果表明该蛋白偏亲水,这与该蛋白缺乏跨膜螺旋结果是吻合的。
2.4 LjC3H2的基因亚细胞定位
利用Euk-mPLOC对LjC3H2蛋白进行亚细胞定位分析,结果显示LjC3H2蛋白可能在内质网上发挥作用。
2.5 LjC3H2蛋白的二、三级结构
采用SOPMA 对LjC3H2的二级结构进行分析,结果表明,α-螺旋(Alpha helix)占比50.20%;无规则卷曲(Random coil)占比33.00%;β-折叠也称延伸链结构(Extended strand)占比11.66%;β 转角(Beta turn)占比5.14%。表明LjC3H2的二级结构是以α-螺旋为主要类型,无规则卷曲较多,β-折叠即延伸链结构数量较少。在SWISS-MODEL对LjC3H2进行三级结构的比较建模分析,结果表明,LjC3H2与细胞色素P450的三级结构较相似(图2)。

图2 LjC3H2和细胞色素P450建模预测的三级结构比较
2.6 LjC3H2与不同植物间的进化关系
以LjC3H2氨基酸序列为样本进行Blast 比对,获得了37 种植物40 个C3H 蛋白氨基酸序列样本,对这些样本进行同源比较和分子进化分析。结果表明,LjC3H2基因编码蛋白的氨基酸序列与蓟、桔梗的C3H 亲缘关系较近,而金银花另一个C3H 蛋白则与龙葵、粗壮女贞、黄芩、芝麻、红花钓钟柳、地黄以及桃儿七等植物的进化关系较近(图3),这表明金银花中两个C3H 拷贝的分子进化路线可能略有不同。此外,这两个蛋白质在数据库中的命名和作用对象也略有不同,其中LjC3H可能更多作用于香豆酰基酯类,LjC3H2则多作用于香豆酸。

图3 37种植物中的40个C3H蛋白的系统进化情况
3 结论与讨论
LjC3H2作为细胞色素P450 CYP98A 亚家族成员,具有该亚家族蛋白高度相似的空间结构,该蛋白分子偏亲水,没有跨膜结构域,没有信号肽,二级结构以α-螺旋为主,无规则卷曲较多,定位于内质网。分子进化方面,与蓟、桔梗的C3H 亲缘关系较近,与同属于金银花的LjC3H亲缘关系较远。
金银花中有2 个C3H 蛋白的基因拷贝[11-12],对LjC3H2的研究相对成熟,因此本文选择以LjC3H2作为研究对象,进行深入分析。结果表明,该酶没有跨膜结构域、没有信号肽,位于内质网上,关于该酶的各类分析表明,该酶主要在细胞内起作用。
对多种植物的C3H 进行系统进化分析,结果表明金银花已知的2 种C3H 蛋白在进化上的亲缘关系并不接近,表明这2 个拷贝的基因在进化上可能分属两条不同的路线,有一定的区别。即这2 个基因拷贝表达的酶蛋白在功能上可能存在细微的差异。后续研究可进一步对这2个基因表达的酶蛋白结构进行精细分析和比较,并通过试验方法研究两者的差异。
本文从数据库收集了37 种植物40 个C3H 蛋白序列,利用生物信息学分析工具,以LjC3H2为研究对象,对该基因表达酶的结构、功能和分类等特点进行了分析研究,为今后继续研究金银花C3H 蛋白酶提供参考。
