黑壳楠染色体核型分析及基因组Survey预测
2024-01-01逯久幸刘燕许朵朵陈鹏李永华刘红利
逯久幸, 刘燕, 许朵朵, 陈鹏, 李永华, 刘红利
(1.河南农业大学风景园林与艺术学院,河南 郑州 450002; 2.河南省优质花卉蔬菜种苗工程技术中心,河南 郑州 450002)
黑壳楠(Linderamegaphylla)属樟科山胡椒属,是优良的常绿阔叶乔木。据中国植物志记载,黑壳楠为中国特有树种,主要分布于陕西、甘肃、湖北、湖南、江西、福建、四川、贵州、云南、广东等。黑壳楠耐阴,原生地主要为山坡、谷地等湿润稍荫蔽的地方。在北方的园林中,常绿阔叶类树木极少,能适应城市绿地环境又具有高观赏价值的常绿阔叶树种更为稀缺,所以针对高观赏价值的常绿阔叶类树种的引种、育种和推广非常有价值[1]。黑壳楠树形优美、四季常青、树干通直、青翠浓郁、抗性强且能分泌杀菌素起到净化空气的作用,又因其木材结构致密,纹理美观也是重要的用材树种[2]。黑壳楠以其优良的特性成为河南地区常绿阔叶类树种推广的重要候选树种之一。河南伏牛山南坡、桐柏山和大别山等800 m以下的山坡或沟谷是黑壳楠自然分布的最北缘[3]。据《河南省重点保护植物名录》记载,野生的黑壳楠已经属于濒危植物[4]。前人对黑壳楠的研究主要集中在分泌物的化学成分、栽培措施、繁育方法和栽培技术、群落结构、物种多样性等方面[5-7],针对黑壳楠细胞遗传学的研究较少。
荧光原位杂交(fluorescence in situ hybridization, FISH)最早起源于20世纪80年代,起初是利用放射性元素(同位素)作为标记,后来逐渐发展为以荧光标记取代同位素标记而形成的一种新型原位杂交技术。它的原理是将DNA(或RNA)探针用特定的核苷酸分子标记,然后将探针杂交到植物组织切片中,再通过荧光染料染色,使特定的核苷酸分子与荧光染料耦联进而激发荧光,在显微镜下就可以观测获得目标DNA(或RNA)分子的定位和相对含量。植物中重要的管家基因之一就是核糖体基因,它包括5S rDAN和45S rDAN。45S rDNA基因包含18S-5.8S-26S rDNA基因,3者纵向排列在染色体上[8]。因此,可以通过定位其中1个基因来定位45S rDNA基因在染色体上的分布情况,本研究选用的是18S rDNA基因。这些基因的转录产物可以与核糖体蛋白结合组成核糖体。核糖体基因多由高度重复的序列组成,它们在植物中高度保守,并通过串联重复的方式分布在染色体的某个或多个部位。利用5S rDAN和18S rDAN探针对植物染色体进行荧光原位杂交,可以定位5S rDNA和18S rDAN在染色体上的位置,进而确定植物染色体的倍性和基因组进化情况。
染色体是遗传信息的载体,对染色体数目、形态和分子特征的分析称为核型分析[9]。遗传物质在细胞层面上的表型特征反应在核型上,能揭示物种在染色体层面上的所有特性,为研究物种起源、演化、亲缘关系、遗传进化等问题提供及其重要的细胞学依据[10]。本研究以端粒重复序列、5S rDNA和18S rDNA为探针,通过荧光原位杂交的方法对黑壳楠染色体进行了精细的核型分析,又通过高通量测序的方法对黑壳楠基因组进行了预测。染色体核型分析和基因组预测为黑壳楠染色体定位、物种起源等细胞遗传学及表观遗传学研究奠定基础,并为进一步良种选育等提供重要的细胞学数据。
1 材料与方法
1.1 试验材料
试验材料取自河南省南阳市西峡县种苗试验站野生黑壳楠种子。种子经过3个月沙藏,取露白的种子播种于穴盘中,待幼苗长至15 cm时,取生长旺盛的侧根根尖制片。
1.2 染色体制备
剪取侧根根尖2~3 cm。将剪好的根用蒸馏水冲洗干净,放入0.5 mL特制离心管中。在离心管中冲入0.9~1.0 MPa的笑气(N2O),静置处理2 h。静置后立即转入冰浴,离心管中加入质量分数90%预冷冰乙酸,静置固定。10 min后取出冲洗,并放入质量分数70%乙醇中保存。制片时先用酶液混合液37 ℃酶解1 h,后用质量分数70%乙醇清洗3次,再用解剖针破碎根尖,震荡离心弃废液。再次加入冰乙酸,充分震荡混匀后,滴入载玻片中央,将染色体标本在普通光学显微镜下镜检。
1.3 探针制备
端粒探针为末端(TTTAGGG)6探针,5S rDNA和18S rDNA探针分别为标记好的pTa794和pTa71质粒DNA。
1.4 荧光原位杂交
将制好的根尖玻片加入标记好的探针溶液,盖上盖玻片,80 ℃热变性5 min,放入杂交盒中,后放入原位杂交箱,在37 ℃的条件下杂交过夜。室温清洗玻片2~3次,将玻片吹干。最后加上8 μL DAPI染液,利用荧光显微镜观察拍照。
1.5 染色体核型分析
染色体核型分析依据李懋学等[11]提出的标准进行,以30个中期分裂相的细胞为对象,统计黑壳楠植物染色体数目。选取5个分散良好、图片清晰的染色体细胞,使用Photoshop图像处理器统计染色体的长度数据。染色体形态判断依据LEVAN等[12]的方法,核型对称性分析依据STEBBINS[13]的标准进行划分。
1.6 基因组survey测序
基因组survey测序由武汉菲沙基因信息有限公司测序完成。
2 结果与分析
2.1 黑壳楠染色体的核型分析
通过对黑壳楠处于细胞中期分裂相的细胞进行观察统计,结果发现,黑壳楠染色体数目为2n=2X=24,其染色体核型参数见表1,染色体核型图见图1-A,染色体核型模式图见1-B。对染色体的长度进行统计测量发现,染色体组绝对长度变异范围为2.92~5.83,相对长度的变化范围为5.33%~11.11%,相对长度组成的最长染色体与最短染色体的比值为2.09。通过相对长度系数计算获得黑壳楠相对长度组成(IRL)为2n=6L+6M2+10M1+2S。通过对长短臂的分析发现,黑壳楠的臂比变化范围为1.08~2.16,臂比大于2的染色体数量百分比为17%,属于2B型染色体。通过每条染色体臂比的分析最终确定黑壳楠的核型公式为:2n=2X=24=18m(2 SAT)+6sm。其中9号染色体存在1对随体。
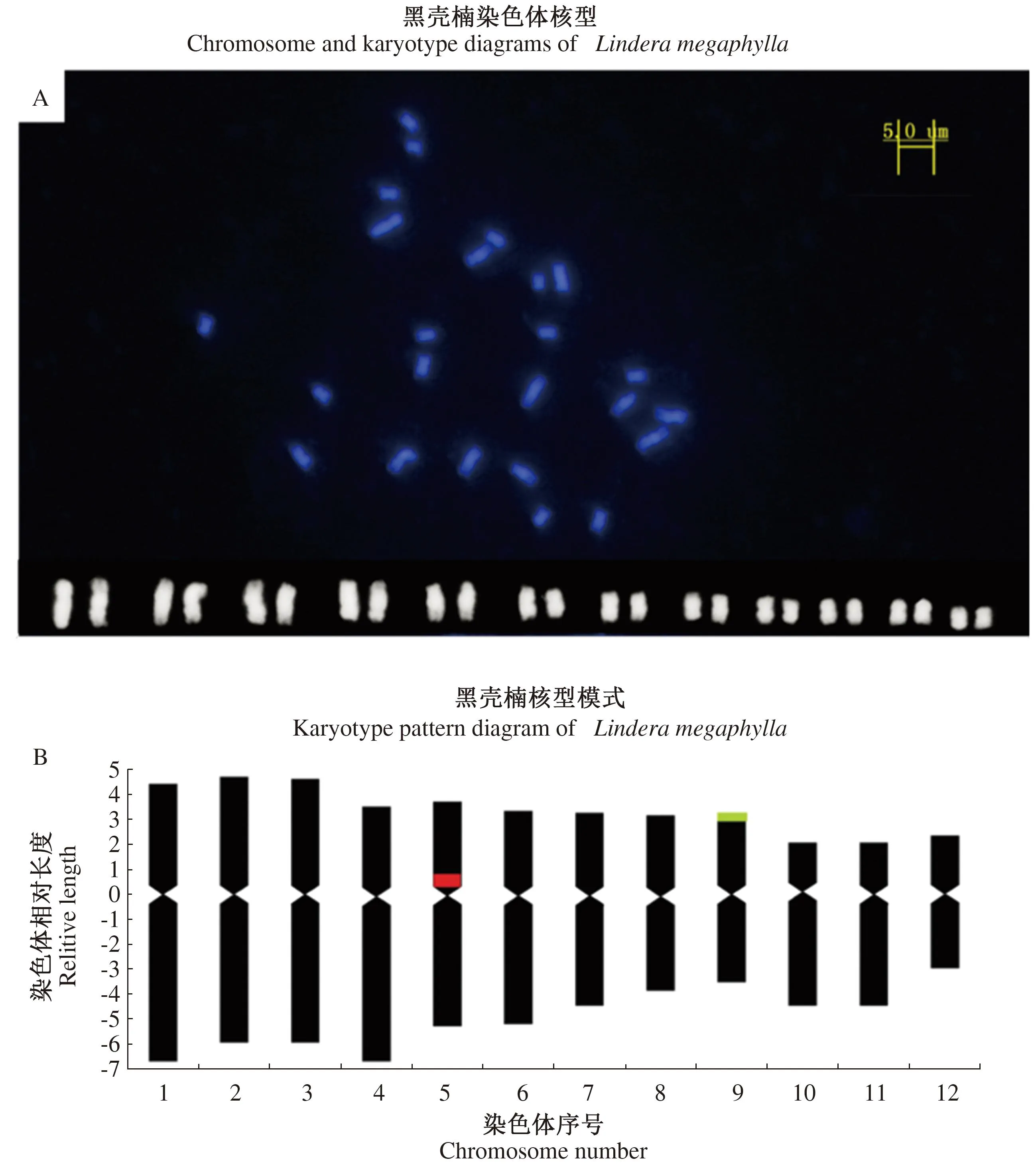
红色信号为5S rDNA荧光杂交信号位置,绿色信号为18S rDNA荧光杂交信号位置。
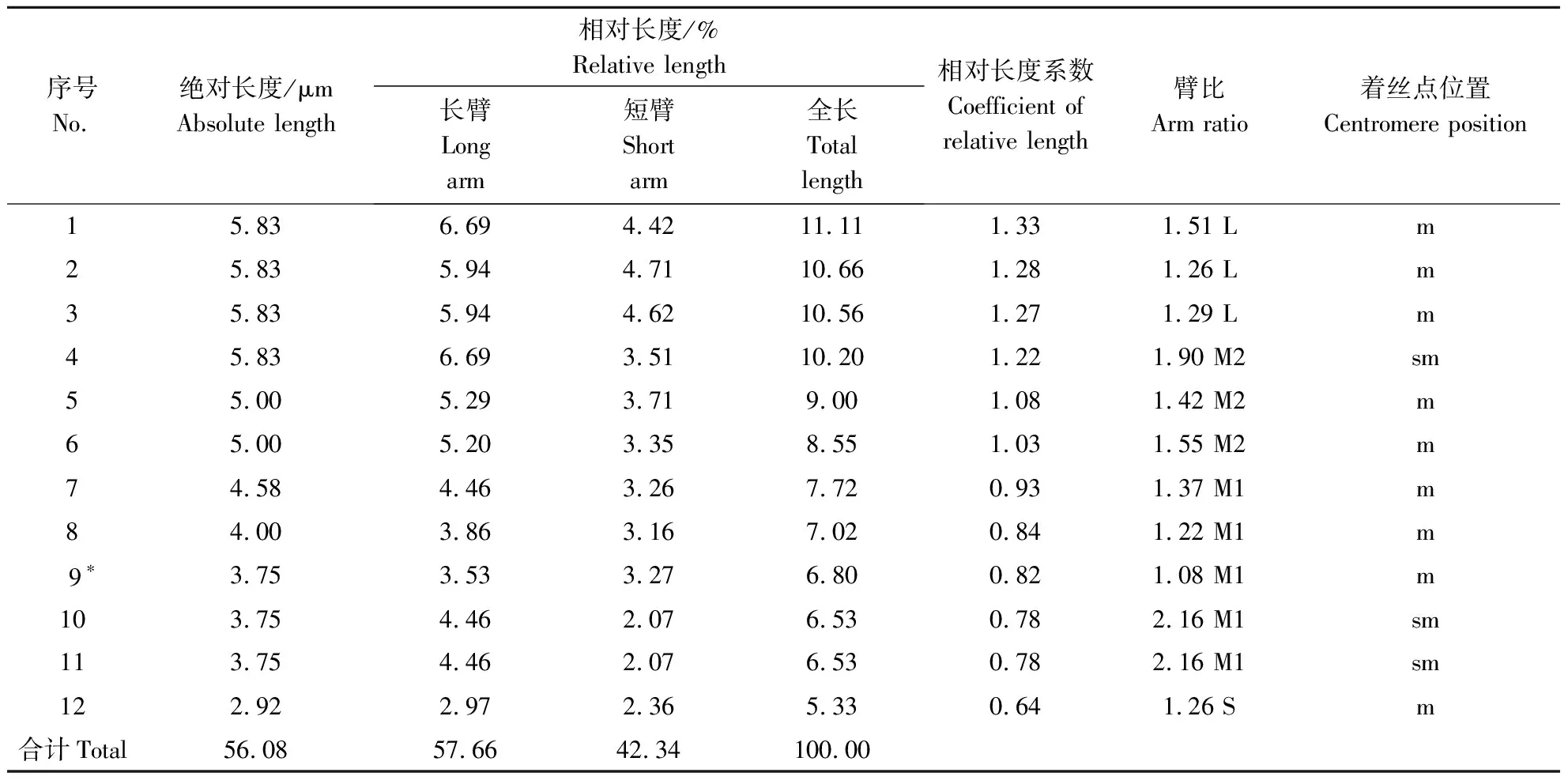
表1 黑壳楠染色体核型参数Table 1 Chromosome karyotype parameters of Lindera megaphylla
从黑壳楠的核型模式图中发现,黑壳楠染色体组共有12对染色体,黑壳楠的组成类型为:9对染色体(1/2/3/5/6/7/8/9/12号染色体)为中部着丝点染色体(m),3对染色体(4/10/11号染色体)为近端部着丝点染色体(sm),通过计算获得黑壳楠核型不对称系数(As.K%)为59.49%,染色体整体较为对称。
2.2 黑壳楠端粒保守序列、5S rDNA和18S rDNA-FISH的定位分析
为进一步确定染色体条数和5S rDNA和18S rDNA在染色体上的分布,以端粒重复序列、5S rDNA和18S rDNA为探针与黑壳楠细胞中期染色体进行杂交。端粒保守重复序列的DAPI染色结果表明,细胞染色体数目为24条,端粒序列位于染色体两端,未发现染色体异常的细胞(图2-A)。黑壳楠5S rDNA和18S rDNA定位结果表明,黑壳楠染色体上具有1对5S rDNA杂交位点(红色信号)和1对18S rDNA杂交位点(绿色信号),4个染色体信号均较强(图2-B)。5S rDNA杂交位点位于第5号染色体的近中心粒区域,而18S rDNA杂交位点位于第9号染色体的短臂近端粒区域,2对杂交信号的位置接近,这也证明黑壳楠为二倍体的结论,且在9号染色体短臂存在1对随体。
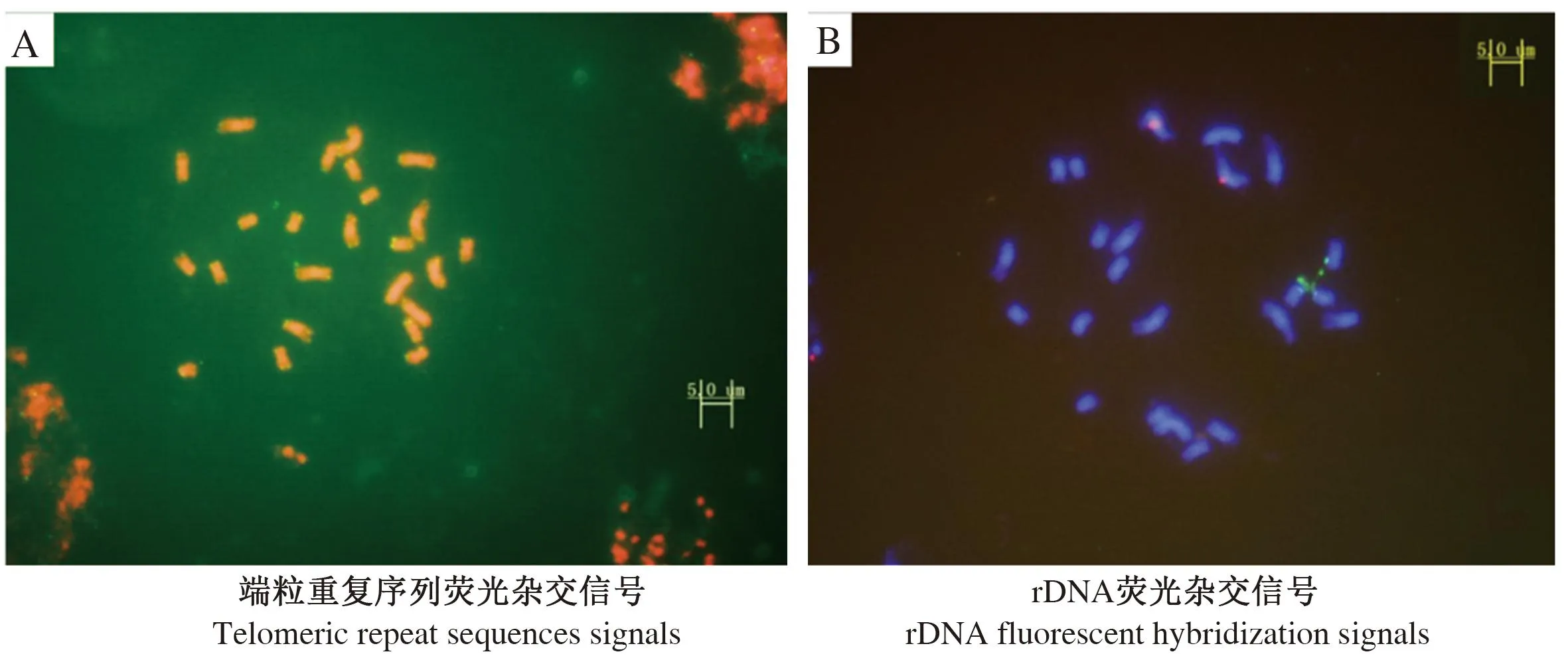
红色信号为5S rDNA荧光杂交信号,绿色信号为18S rDNA荧光杂交信号。
2.3 黑壳楠的基因组预测
基于小片段文库的深度测序,通过K-mer分析,获取了黑壳楠基因组的基本信息(图3、表2、表3)。结果显示,测序共获得50 201 264 Gb的clean data。通过测序质量评估发现,Q30大于93%,测序错误率分布合理,C和G的比例接近,说明测序质量优,符合进一步分析要求。CG含量呈现单峰趋势(图3),NT库对比到黑壳楠的近缘物种DNA上说明取材未有明显的外源污染(表3)。
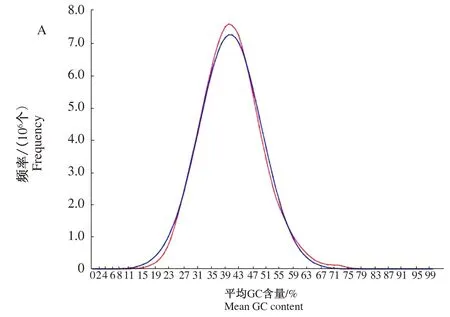
图3 黑壳楠基因组Survey测序数据GC分布图

表2 黑壳楠基因组Survey测序数据统计Table 2 The genome sequencing data of Lindera megaphylla

表3 质控后数据NT库对比结果Table 3 Blast results in NT database of row data
基因组survey结果显示,黑壳楠预估基因组大小为1.38 Gb,测序深度为30倍,杂合率为0.8%,重复序列比例为70%。所以黑壳楠为大基因组,杂合率较高,重复序列多的物种。
3 结论与讨论
不同种源染色体存在多态性。本研究结果显示,南阳西峡的野生黑壳楠的染色体核型为2n=2X=24=18m(2SAT)+6sm,除4/10/11号染色体为近端部着丝点染色体(sm)外,其他均为中部着丝点染色体(m),9号染色体短臂端粒处含1对随体,未观测到异性染色体细胞。这个核型结果与采自都江堰的黑壳楠核型一致,但与采自南充的黑壳楠存在差异[14]。陈成斌等[14]研究发现,采自南充的黑壳楠的染色体核型为2n=18m+6sm+3B,含有3~6个不等的B染色体细胞,这种差异的出现可能是由于不同的种源地黑壳楠B染色体存在多态性。尽管如此,3者染色体核型相同,均属于2B型染色体。根据相对长度系数进行的染色分类,3者存在一定差异。西峡黑壳楠染色体相对长度组成为2n=6M+6M2+10M1+2S,而南充黑壳楠为2n=6M+4M2+8M1+6S,都江堰黑壳楠为2n=6M+4M2+10M1+4S。这种差异的存在可能是由于染色体制片方法的不同而产生的。本研究采用的是酶解去壁滴片法制片,后2者采用的是卡宝品红压片法制片。与传统的压片法相比,酶解去壁滴片法呈现的染色更加清晰,染色体形态也更加优质。这种方法也广泛应用在向日葵、蔷薇、毛竹等物种上,是更利于荧光定位分析的方法[15-21]。另外,在取样时间和处理上与3者均不同。可能拍照时所选择的细胞染色体回缩的程度存在差异,从而展现在长度上就会不同。另外,后两者在计算染色体长度时使用的是裁剪法,显微镜清晰度较差,精度相对较低;而前者采用的是软件绘制,高清晰度显微摄像,精度较高。虽然3者相对长度存在差异,但最终计算获得的染色体核型不对称系数却十分接近,西峡、南充和都江堰黑壳楠分别为59.49%、59.36%和60.35%[14],数值接近50%为较对称染色体,这也标志着在樟科的进化中,黑壳楠为较为原始的物种。
荧光原位杂交技术以其快速、稳定、特异度高的优点广泛应用于植物细胞遗传学研究中。它可以用来构建物种的物理图谱,鉴定转基因植株、阐明基因组的结构和进化过程或分析物种亲缘关系[22-25]。rDNA是一种精准的分子遗传学标记,常作为探针应用于荧光原位杂交中。利用rDNA为标记多用于研究植物多拷贝基因的形成和演化、鉴定植物染色体组型、多倍体的起源与非整倍体,还可以研究基因组的进化等问题[26-28]。常见的rDNA有5S rDNA和45S rDNA。45S rDNA与核仁相连,伴随着随体的出现,所以其位点的个数也代表了随体的个数[29]。45S rDNA杂交信号的强度可以间接反应基因拷贝数的多少。5S rDNA标记往往用来鉴定植物的倍性情况[30]。本研究发现,黑壳楠在9号染色体短臂端粒含有1对45S rDNA强杂交信号,位置相同,强度相近。在5号染色体含有1对5S rDNA杂交信号,这也说明黑壳楠45S rDNA在9号染色体上存在1对随体,基因的拷贝数相近,且染色体未发生重组、变异或者种内杂交等现象,且所检测野生黑壳楠为二倍体。
基因组survey是在高通量测序的基础上预测基因组的手段。对基因组的大小、杂合度和重复序列预测对精确的植物基因组测序有重要的指导意义,这种手段已经广泛应用在樟、四合木、西番莲等多种植物基因组测序中[31-32]。目前所报道的植物基因组的大小约为63 Mb~127 Gb[33]。在已报道的樟科植物中,樟属樟树基因组大小为723.12 Mb,牛樟732.7 Mb,樟科山胡椒属木姜子1.3 Gb,樟科鳄梨属为2.24~2.46 Gb[34-37]。基因组最大是鳄梨属植物,最小是樟属植物。本研究发现,黑壳楠属山胡椒属,基因组大小为1.38 Gb,基本与同属木姜子基因组大小相同,属较大基因组。黑壳楠基因组的杂合率为0.8%,重复序列比例为70%。说明其属于高杂合率和高重复率的基因组,这也为下一步基因组测序指明了方向。
