桑树Aux/IAA基因家族鉴定与IBA处理下表达模式分析
2024-01-01冯曦豆浩孙佳佳孙慧娟毕会涛
冯曦, 豆浩, 孙佳佳, 孙慧娟, 毕会涛
(河南农业大学林学院,河南 郑州 450002)
Aux/IAA是诱导生长素响应调节作用的关键转录因子蛋白家族[1],Aux/IAA基因家族通过表达核定位蛋白[2],影响了生长素的早期反应[3]。在高浓度生长素水平下,生长素在TIR1/AFB基因家族所构建的生长素结合域中与Aux/IAA蛋白结合[4],形成TIR1/AFB-Aux/IAA生长素共受体[5],并作为SCF复合体一部分的F-box蛋白(Skp1/Cullin/F-box)与生长素结合[6],形成SCFTIR1-生长素-Aux/IAA复合体,使Aux/IAA蛋白泛素化后降解[7-8]。在低浓度生长素水平下,ARF蛋白与Aux/IAA蛋白互作,促进转录抑制[9]。Aux/IAA蛋白包括4个高度保守的结构域(Domain Ⅰ, Ⅱ, Ⅲ, Ⅳ)[10]:Domian Ⅰ 存在着与乙烯响应因子(ERF)相关的转录抑制子(ERA)结构域[11],转录阻遏因子(TPL)通过该结构域,从而抑制生长素信号下游基因的表达[12];Domian Ⅱ 具有高度保守的特性,受TIR1调控,可以直接结合SCFTIR1控制Aux/IAA蛋白降解[13];Domian Ⅲ、Ⅳ 是与ARF相结合的部位,具有ARFs蛋白的CTDs的同源结构域,负责与其他Aux/IAA蛋白的二聚化和多聚化[14-15],进而调控生长素信号。
Aux/IAA基因家族的表达模式比较复杂,目前在拟南芥 (Arabidopsisthaliana)[16]、番茄 (Lycopersiconesculentum)[17]、玉米 (Zeamays)[18]、龙眼 (DimocarpuslonganLour)[19]中分别发现了29、36、40、18个Aux/IAA基因。拟南芥光敏色素A通过促进Aux/IAA蛋白质稳定性负调避荫反应[20],AtIAA7参与植物的向地性生长[21],AtIAA5、AtIAA6、AtIAA19能够调节拟南芥的耐旱特性[22],AtIAA32能够调控顶端弯钩发育的作用[23];Sl-IAA17调控番茄果实发育过程中的品质参数[24];GRMZM2G115357在高渗透压条件下高表达,可能参与了玉米对高渗透胁迫的抗性过程[18];Dl-IAA基因参与了龙眼花芽分化过程,Dl-IAA2和Dl-IAA17可能在开花诱导中起作用[19]。
桑树(Morusnotabilis)是桑科桑属的落叶乔木,原产于中国[25],是经济生态兼用型树种[26]。桑树可以培育成叶用、果用、材用的乔木林,也可以培育成菜用的灌木林,还能适应多种立地条件[27]。桑树具有优良的产业优势,一年栽植可多年利用[28]。桑树栽植通常采用嫁接育苗[29],嫁接苗具有成活率高的优势,但也存在育苗成本高,技术要求严格等缺陷[30]。扦插因具有育苗周期短,繁育成本低的优势,成为桑树快速产业化的有效途径[31]。前期研究发现,生长素在桑树扦插生根过程中起着重要的调节作用[32],但目前控制桑树扦插生根的基因表达模式尚不清楚,桑树Aux/IAA基因家族的全基因组分析也未见报道。
本研究鉴定了桑树Aux/IAA基因家族成员的蛋白序列,并对其理化性质,系统发育树、基因结构、蛋白互作关联以及启动子顺式作用元件进行了较为全面的生物信息学分析。并研究了在IBA处理下的桑树Aux/IAA基因家族的表达模式,推测了部分MnAux/IAA基因的功能,为后续阐明桑树生根过程的研究奠定了基础。
1 材料与方法
1.1 数据来源
桑树全基因组序列和蛋白序列从本地川桑(Morusnotabilis)基因组数据库中获取,拟南芥全基因组序列和蛋白序列从TAIR数据库(https://www.arabidopsis.org/)下载。
1.2 桑树Aux/IAA基因家族成员提取鉴定
根据LISCUME等[33]提到的29个拟南芥AtAux/IAA基因家族成员,构建本地BLAST数据库(E-value<0.05),获得51条拟南芥的同源蛋白序列;同时从Pfam数据库(http://pfam.xfam.org/)下载具有Aux/IAA蛋白结构域(PF02309)的隐马尔可夫模型文件(hidden markov model, HMM),并使用HMMER3.0软件对川桑全基因组数据库进行检索(E-value<0.05),筛选出51条待定序列。将上述两方法所得到的序列取交集,筛选出具有完整Aux/IAA结构域的蛋白序列。并命名为MnAux/IAA1~MnAux/IAA51。
1.3 桑树Aux/IAA基因家族成员理化性质预测
利用在线软件ExPASy-ProtParam(https://web.expasy.org/protparam/)对筛选得到的51条MnAux/IAA蛋白质的理化性质进行预测。包括氨基酸数、相对分子质量、等电点、平均疏水指数。用在线软件WoLF PSORT(https://wolfpsort.hgc.jp/)预测亚细胞定位。
1.4 桑树Aux/IAA基因家族成员的系统进化树构建
利用MEGA7.0软件的Muscle比对方式,对桑树和拟南芥Aux/IAA基因家族的蛋白序列进行比对,并使用邻接算法(neighbor-joining, NJ)构建系统进化树,Bootstrap重复值设置为1 000。最后用evolview在线网站(https://www.evolgenius.info/evolview/)对系统进化树进行分类和注释。
1.5 桑树Aux/IAA基因家族成员的保守基序与基因结构预测
利用在线工具MEME(http://meme-suite.org/tools/meme)对桑树Aux/IAA基因家族成员的保守基序进行分析,设置motif显示数量为10。再根据川桑基因组的注释文件对MnAux/IAA基因结构进行分析。最后用TBtools软件(https://github.com/CJ-Chen/TBtools)对分析结果进行可视化。
1.6 桑树Aux/IAA基因启动子顺式作用元件预测
将MnAux/IAA基因家族成员的编码区上游的2 000 bp序列作为启动子区域进行提取。用PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)对启动子区域的顺式作用元件分析进行预测分析,最后用TBtools软件进行可视化。
1.7 桑树Aux/IAA基因家族蛋白三级结构和蛋白互作关联预测
利用在线网站SWISS-MODEL(https://swiss-model.expasy.org/interactive/),对鉴定出的MnAux/IAA蛋白序列进行蛋白三级结构预测。随后将MnAux/IAA蛋白序列与已知的AtIAA和AtARF蛋白序列进行比对。然后利用在线网站STRING(https://cn.string-db.org/),将51个MnAux/IAA基因家族蛋白序列作为查询目标并预测蛋白互作(protein-pro-tein interaction, PPI)网络。以AtIAA基因和AtARF基因与MnAux/IAA基因家族的相似度为标准,以置信度为0.7的基因作为PPI网络中的枢纽基因来构建网络图,并用Cytoscape软件进行可视化。
1.8 桑树Aux/IAA基因在IBA不同时期处理下的表达模式测定
选取桑树品种“强桑一号”的扦插枝为试验材料,进行1 000 mg·L-1的IBA处理和清水处理,处理30 s后扦插至基质中,后续选取10、20、30、40 d的插穗进行转录组测序。统计IBA处理后10、20、30和40 d(IBA-1、IBA-2、IBA-3、IBA-4)以及相应的清水(CK-1、CK-2、CK-3、CK-4)对照组的转录组数据。分别比对到每个MnAux/IAA基因的序列数量,采用RPKM(reads per kilobase of tran-scripts per million mapped reads)对每个基因表达量进行均一化处理,最后使用TBtools软件绘制MnAux/IAA基因家族成员在IBA处理前后的表达热图。
2 结果与分析
2.1 桑树Aux/IAA基因家族成员鉴定及理化性质分析
在桑树全基因组中,共鉴定出51个Aux/IAA基因家族成员,依据基因编号大小对桑树Aux/IAA基因家族成员依次命名为MnAux/IAA1~MnAux/IAA51(表1)。桑树Aux/IAA基因家族成员理化性质分析显示,发现它们在氨基酸数量、等电点、相对分子质量方面存在显著差异。51个MnAux/IAAs蛋白的氨基酸长度在79 (MnAux/IAA51)~1 144 aa (MnAux/IAA3)范围内,其中小于400 aa的有27个,所占比例为52.9%;蛋白质相对分子质量差异较大,为8.99(MnAux/IAA51)~127.70 kD (MnAux/IAA3);72.55%的MnAux/IAAs蛋白等电点(pI)小于7,平均值6.7,说明这些MnAux/IAAs蛋白大部分呈酸性;蛋白质平均疏水指数均为负值,表明MnAux/IAAs均为亲水性蛋白质;亚细胞预测结果显示,有40个成员定位于细胞核上,7个定位于叶绿体,2个定位于过氧化物酶体,1个定位于细胞骨架,1个定位于线粒体,说明该家族基因主要在细胞核中发挥功能。
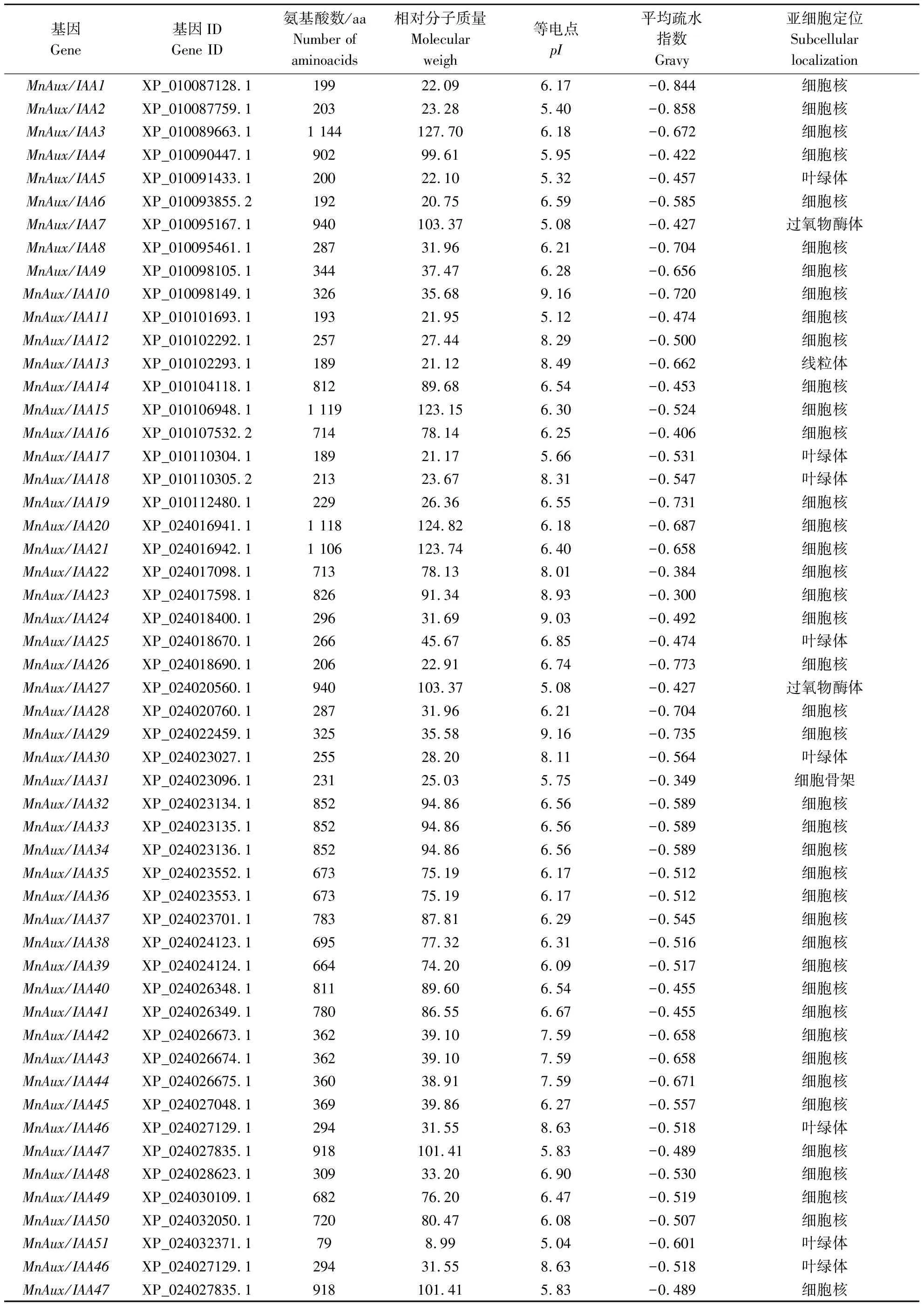
表1 桑树Aux/IAA基因家族基本信息Table 1 Basic information of Aux/IAA gene family in Morus notabilis

表2 桑树MnAux/IAA基因家族蛋白互作预测Table 2 Protein interaction prediction of MnAux/IAA gene family in Morus notabilis
2.2 桑树Aux/IAA基因家族系统进化分析
根据拟南芥AtIAA蛋白序列和鉴定出的桑树MnAux/IAA蛋白序列构建了系统进化树(图1)。根据系统进化分析将51个MnAux/IAA基因家族成员大致分成8个亚家族,8个亚家族分别命名为A1 ~ B4。其中A1、A2、A3、A4、B1、B2、B3和B4亚家族分别包含10、11、4、5、5、4、3和9个MnAux/IAA基因,而且桑树与拟南芥Aux/IAA基因家族成员在6个亚家族中的分布类似。分析结果表明,MnAux/IAA基因和AtIAA基因同源性很高,Aux/IAA基因物种进化比较保守,蛋白序列的相似性。表明2个物种的Aux/IAA基因可能具有相似的生物学功能。从进化树分支上看,桑树与拟南芥的Aux/IAA基因家族成员在各亚家族上的分布数所占比例不同,说明Aux/IAA基因家族在不同植物亚家族中聚类存在差异。
2.3 桑树Aux/IAA基因家族成员的保守基序与基因结构分析
保守基序分析结果显示(图2-B)。不同MnAux/IAAs中所包含的motif数量存在一定差异,亲缘关系相近的成员,其所含的motif也大多相似。51个MnAux/IAAs中,有15个MnAux/IAAs包含9种motif。几乎所有家族成员都有motif 1,这说明motif 1是桑树MnAux/IAA基因家族的保守结构域的一部分。比较发现,有较多的蛋白保守基序按照motif 4、motif 9、motif 3、motif 2、motif 5、motif 6、motif 7的顺序排列,该结果可能与MnAux/IAA基因的特定功能有关。结构域分析发现,有23个MnAux/IAAs包含B3DNA结构域,49个MnAux/IAAs包含羧基末端的PB1结构域。
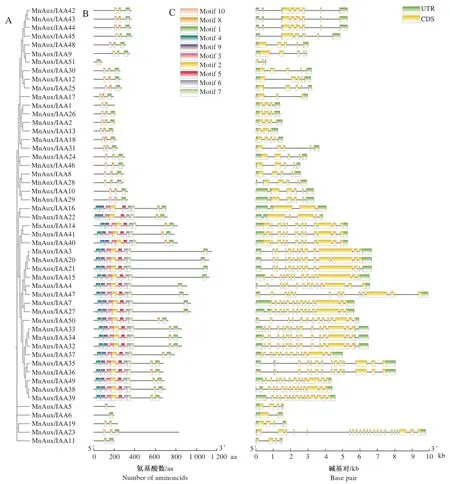
A: MnAux/IAA基因家族系统发育树;B: MnAux/IAA基因家族成员的保守基序;C: MnAux/IAA基因家族成员的基因结构。
基因结构分析结果显示(图2-C),MnAux/IAA基因家族中均含有内含子,除MnAux/IAA6含有1个内含子外,其余均含有2个及以上内含子。不同成员所含有的外显子数量也不同,分布为3~22个,其中MnAux/IAA23含有最多的外显子(22个),MnAux/IAA6的外显子数量最少(2个)。外显子在5~14个的基因数最多,占80.39%。另外各亚家族间的基因长度差异较大,B2、B4两个亚家族的基因长度较短。其中MnAux/IAA51基因长度最短;MnAux/IAA23的基因长度最长。所有的基因家族成员都有外显子和内含子,仅有1个基因家族成员(MnAux/IAA51)无非编译区(untranslated region, UTR),说明MnAux/IAA基因家族成员的功能比较稳定,在家族进化过程中变化较小。
2.4 桑树Aux/IAA基因家族成员的顺式作用元件分析
分析MnAux/IAA基因上游2 000 bp启动子序列区域,得到Aux/IAA基因家族的启动子顺式作用元件预测结果(图3)。在MnAux/IAA基因中共检测到19种顺式作用元件,主要包括4种类型:光响应型、激素响应型、逆境响应型和发育响应型。其中,激素响应型方面。MnAux/IAA基因的启动子上存在5种激素相关的顺式作用元件。5种顺式作用元件的数量存在差异,其中茉莉酸甲酯响应元件占比例最多,达到14.68%,赤霉毒响应元件所占比例为7.01%。生长素响应元件数量最少,仅占3.75%。从整体来看,厌氧感应元件所占比例最大,达到18.10%。整体看来,MnAux/IAA49上的顺式作用元件最多(21个),其余大多数都在11~17个之间。同时发现,同一元件在不同基因上的分布不均衡。生长素响应元件主要分布在MnAux/IAA31、MnAux/IAA38、MnAux/IAA39的启动子上。

图3 桑树MnAux/IAA基因启动子顺式作用元件分布情况Fig.3 Distribution of cis-acting elements of MnAux/IAA gene promoter in Morus notabilis
2.5 桑树Aux/IAA基因家族蛋白三级结构及蛋白互作关联分析
桑树MnAux/IAA蛋白三级结构预测如图4所示,桑树MnAux/IAA蛋白包含α-螺旋、β-折叠、β-转角以及无规则卷曲等空间构象。对比发现,蛋白质整体相似度水平不高,但同一个亚家族的蛋白质的空间结构比较接近。例如A1亚家族的MnAux/IAA35、MnAux/IAA36、MnAux/IAA38和MnAux/IAA39;B4亚家族的MnAux/IAA42、MnAux/IAA43、MnAux/IAA44。表明蛋白结构与物种进化同源性有的一定关联。

图4 桑树MnAux/IAA基因家族蛋白三级结构预测Fig.4 Prediction of tertiary structure of MnAux/IAA gene family proteins in Morus notabilis
蛋白互作关联预测如图5所示,将筛选出的MnAux/IAA蛋白,与拟南芥中基因结构相似的AtIAAs和AtARFs进行比对,利用AtIAAs和AtARFs所构建的蛋白互作PPI网络,推测MnAux/IAA蛋白之间的调控关系。对比发现,MnAux/IAA蛋白之间存在多种调控通路。IAA3、IAA4、IAA16、IAA17、IAA19、IAA26具有较多的网络交叉联系,说明这些蛋白对植物某些生理功能和生长发育等一系列过程可能具有协同调控作用。AtARF1和AtARF2能调控叶片的衰老、花器官的发育与凋谢[34],AtARF5主要参与维管组织的形成及胚的发育[35],AtIAA3,AtIAA14,AtIAA28,AtIAA19与侧根的生长发育有关[16]。说明与其同源的MnAux/IAA蛋白有相似的功能。

图5 桑树MnAux/IAA基因家族蛋白互作Fig.5 Protein interaction of MnAux/IAA gene family in Morus notabilis
2.6 桑树Aux/IAA基因在IBA处理下的表达模式分析
以“强桑1号”扦插苗为试验材料,对其采用质量浓度为1 000 mg·L-1的IBA处理30 S,并选取10、20、30、40 d 4个不同时期的扦插苗进行转录组测序,从测序数据中筛选出51个MnAux/IAA基因的表达量数据并绘制热图(图6)。
51个Aux/IAA基因在两种处理条件下均有不同程度的表达。在清水处理下,有6个基因随着处理时间的延长,其表达量保持上调趋势;有2个基因表达量保持下调趋势。经过IBA处理后,有13个基因随处理时间的延长,其表达量保持上调趋势;有8个基因表达量保持下调趋势。在IBA-1的处理条件下,B2亚家族中所有的MnAux/IAA基因的表达量显著高于其他处理时期;在CK-A的处理条件下,处于B4亚家族的MnAux/IAA23和MnAux/IAA48两个基因表达量尤为突出,这说明同一亚家族中的基因表达模式具有一致性和相似性。
在IBA-3和IBA-4的处理条件下,MnAux/IAA的整体表达水平不如IBA-1和IBA-2,猜测是由于处理时间过长导致IBA对MnAux/IAA表达的影响逐渐降低。整体来看,92.2%的基因在经过IBA处理后,基因表达程度上会有不同程度的提升,这说明IBA处理对MnAux/IAA基因的表达具有良好的促进作用。MnAux/IAA34、MnAux/IAA33、MnAux/IAA32在经过IBA处理后在表达量上有较大程度的下降,推测IBA抑制了这3种基因的正常表达。
3 结论与讨论
Aux/IAA基因家族是植物生长素在表达过程中参与反应的重要基因家族,在不同浓度的生长素下具有相应的调控作用[36],也是生长素最早进行信号转导以及基因表达的3个基因家族之一。现如今Aux/IAA蛋白功能已经被人所知,但对桑树Aux/IAA基因家族的研究还未见报道。本研究利用本地川桑(Morusnotabilis)基因组数据,最终鉴定出51个MnAux/IAA基因家族成员,高于拟南芥29个[16]、番茄36个[17]、玉米40个[18]、龙眼18个[19]。理化性质分析发现,MnAux/IAA蛋白的氨基酸个数与相对分子质量差异较大,大部分MnAux/IAA的等电点小于7,平均值6.7,显酸性;系统进化树分析将MnAux/IAA家族分为8个亚家族,各亚家族内的基因结构基本相似表明,Aux/IAA基因在物种进化的过程中具有较高的保守性;保守基序与基因结构分析发现,含有PB1结构域基因成员较多,且仅有1个基因家族成员无UTR,说明MnAux/IAA基因家族成员的功能稳定。蛋白三级结构分析表明,同一个亚家族的蛋白质的空间结构比较接近。蛋白互作分析发现,MnAux/IAA与AtIAA在部分基因上具有相似的功能。
本研究中,有47个基因经过IBA处理后,4个时期的平均表达量高于清水对照组,说明IBA具有促进Aux/IAA基因表达的作用。经过IBA处理后,基因MnAux/IAA34、MnAux/IAA33、MnAux/IAA32的表达量持续显著降低,蛋白互作分析结果表明,MnAux/IAA34、MnAux/IAA33、MnAux/IAA32与AtARF2同源性较高。调查发现,AtARF2具有促进调控叶片衰老和花器官脱落的作用[37],推测MnAux/IAA34、MnAux/IAA33、MnAux/IAA32可能与叶片生长发育的功能相关。桑树扦插苗在IBA处理10 d后,MnAux/IAA18、MnAux/IAA31基因表达量显著,且与AtIAA17有较高的同源性,调查发现AtIAA17具有调控植物下胚轴长度、根毛数量及保证叶形正常的功能[38]。因此,推测MnAux/IAA18和MnAux/IAA31具有促进生根、调控叶形的功能。
生长素在植物早期的调控作用与Aux/IAA基因的表达密切相关。在拟南芥中,AtAux/IAA家族成员AXR3的突变体AXR3(IAA17)具有下胚轴明显变短,叶向上卷曲且呈黑色及无根毛等表型[39];王婧[40]利用酵母双杂交试验证明,AtlAA8与ARF6和ARF8互作,表明Aux/IAA参与调节花器官的发育;在水稻中,NAKAMURA等[41]将OsIAA31过表达,引起株系叶片变短,不定根减少。本研究中,B2、A1、A2亚家族内部的基因表达量相似,证明同一亚家族内基因表达模式大致相同。随着IBA处理时间的增加,MnAux/IAA基因的表达量普遍下降,证明IBA作用效果随时间增加而出现缓慢下降的趋势。
结合桑树扦插苗受IBA处理后的基因表达量变化,发现IBA具有促进Aux/IAA基因表达的作用;推测MnAux/IAA34、MnAux/IAA33、MnAux/IAA32具有调控叶片衰老和花凋落的功能;推测MnAux/IAA18和MnAux/IAA31与根毛数量的维持和叶片正常舒展密切相关,但其具体功能仍需进一步实验验证。通过本研究,为桑树Mux/IAA基因家族在调控扦插生根过程中提供了候选基因。探究了与植物激素相关的基因家族对扦插生根的表达作用,为深入研究Aux/IAA基因家族的功能提供了基础。为桑树扦插生长发育在分子水平上提供了参考条件。
