植物多不饱和脂肪酸的研究进展
2023-10-17蔡璨白玉韩艺郭佳欣沙伟张梅娟彭疑芳马天意
蔡璨,白玉,韩艺,郭佳欣,沙伟,张梅娟,彭疑芳,马天意
植物多不饱和脂肪酸的研究进展
蔡璨,白玉,韩艺,郭佳欣,沙伟,张梅娟,彭疑芳,马天意
(齐齐哈尔大学 1. 生命科学与农林学院,2. 抗性基因工程与寒地生物多样性保护黑龙江重点实验室,黑龙江 齐齐哈尔 161006)
多不饱和脂肪酸(polyunsaturated fatty acids,PUFAs)是一类独特的生物活性物质,在植物中具有调节生长发育和抵御外界胁迫过程的作用.多不饱和脂肪酸在一系列去饱和酶和延长酶的作用下,通过不同代谢途径形成结构各异、功能相关的化合物,这些化合物作为信号分子特异性调控不同生物学功能及相关基因表达,使植物增强对不同环境胁迫的抗性.介绍了植物中多不饱和脂肪酸的来源、分类、合成途径以及在植物抵御外界胁迫中的作用,为深入研究植物中多不饱和脂肪酸的功能奠定基础.
多不饱和脂肪酸;脂肪酸去饱和酶;环境胁迫
植物在生长发育过程中会遭受各类环境胁迫,多不饱和脂肪酸(polyunsaturated fatty acid,PUFAs)是植物细胞膜和角质蜡的关键成分,可以重塑膜的流动性,作为植物体防御基因表达的调节剂,为植物体提供结构屏障[1].PUFAs也可以在果实发育过程中通过脂氧合酶或氧化途径参与香气物质的合成[2],也可以在细胞中经过氧化分解形成CO2和水,为生物体活动提供能量[3].此外,PUFAs在植物体遭遇病菌和生物胁迫时会担当起信号分子的角色[4].近年来,PUFAs作为抵御生物和非生物胁迫的一般防御系统而被关注.PUFAs的合成是一个连续的过程,以饱和脂肪酸为底物,在一系列去饱和酶和延长酶的作用下引入双键形成[5]5-6.脂肪酸去饱和酶(fatty acid desaturase,FAD)在维持细胞膜的稳态中起到关键作用,可将脂肪酸链特定位置的碳碳单键转化成碳碳双键,产生PUFAs来调控植物对外界胁迫的响应[6].
为了更好地研究PUFAs在植物中的作用机制,概括了PUFAs在植物中的来源、分类、合成途径、FAD基因的分离与克隆,为进一步在植物中研究PUFAs提供了理论依据.
1 多不饱和脂肪酸的基本结构及分类
PUFAs在海洋藻类中含量丰富,海洋微藻是PUFAs的初级生产者.在20世纪80年代初期,开始了微藻生产PUFAs的研究.PUFAs又叫做多烯酸,是碳氢化合物长链,由18~22个碳原子组成,主链结构中含有多个碳碳双键[7-8].
PUFAs依据甲基末端第1个双键位置分为2个重要家族:n-3和n-6家族.n-3家族是-亚麻酸(-Linolenic acid,ALA)的衍生物,包括二十二碳六烯酸(Docosahexaenoic Acid,DHA,n3-22:6)和二十碳五烯酸(Eicosapentaenoic acid,EPA,n3-20:5);n-6家族是亚油酸(Linoleic acid,LA)的衍生物,包括花生四烯酸(Arachidonic acid,ARA,n6-20:4)和-亚麻酸(-Linolenic Acid,GLA)[9].其中,ALA和GLA合称为亚麻酸(Linolenic acid,LNA).
ALA在紫苏()和亚麻()中含量最高.植物中ALA的合成是以磷脂酰胆碱为最初底物,经磷脂酶(Phospholipase,PLA)和FAD作用合成.DHA和EPA存在于海洋植物中,最初是由海洋微藻合成.LA主要存在于植物油中.2002年,沈继红[10]等运用茵陈蒿()种子中的亚油酸合成了共轭亚油酸,此后人们展开了大量研究.ARA普遍存在于苔藓和蕨类等植物中,种子植物中仅含有微量的ARA.GLA在月见草()种子中含量较高,1919年首次在月见草中发现并命名了GLA[11].
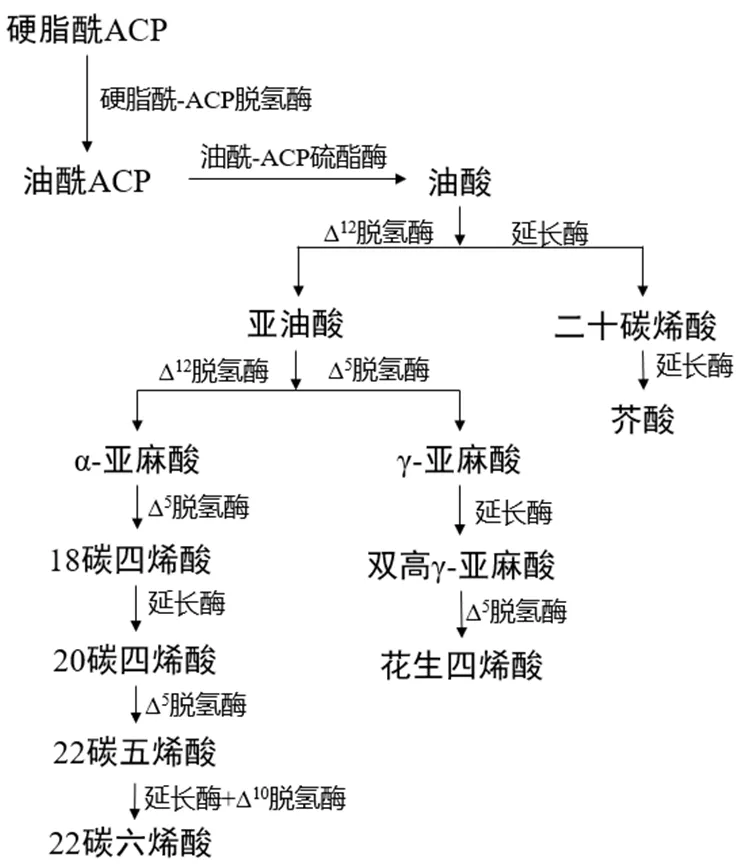
图1 高等植物中PUFAs生物合成途径
2 植物多不饱和脂肪酸的合成途径
在植物中,PUFAs通常是通过修饰饱和脂肪酸前体合成的,饱和脂肪酸前体是脂肪酸合成酶的产物[5]7-8.高等植物中PUFAs的合成途径见图1.根据合成位置和方式不同又分为原核和真核途径.原核途径发生在叶绿体中,饱和脂肪酸在甘油-3-磷酸酰基转移酶和单酰基甘油-3-磷酸酰基转移酶的作用下形成甘油酯;真核途径是指饱和脂肪酸水解后转移至内质网中,通过甘油-3-磷酸酰基转移酶和单酰基甘油-3-磷酸酰基转移酶的催化作用合成甘油酯[12].藻类植物中含有一系列去饱和酶和延长酶,可以从头合成PUFAs.
3 多不饱和脂肪酸去饱和酶的生物学功能
FAD分布广泛,除一些细菌之外,几乎存在于所有生物群中,是植物PUFAs合成过程中的关键酶,在维持生物膜的适当结构和功能方面起关键作用.来自植物膜结合脂肪酸去饱和酶的氨基酸序列有3个高度保守的富含氨基酸序列(组氨酸盒),Δ5脂肪酸去饱和酶(FAD5)含有细胞色素b5样的N-末端延伸,通常伴随着将第3个HiS框改为QXXHH[13].
在较多研究中发现,植物FAD编码基因的表达受环境胁迫影响.1990年,Wada[14]等对蓝藻(sp. PCC6803)进行低温诱导后与等温条件下的蓝藻中脂质含量相比,发现温度降低诱导了FAD的活性,之后在该物种中成功克隆出基因,其过表达提高了PUFAs的积累[15].2002年,反义表达拟南芥()基因,降低了转基因烟草()的耐盐、耐旱性[16].2005年,在烟草植株和细胞中异位过表达了拟南芥中的和,2个基因的过表达增加了烟草植株对干旱胁迫的耐受性,烟草细胞对渗透胁迫的耐受性[17].2014年,运用实时荧光定量PCR技术在竹子()中鉴定了对冷胁迫的动态响应,对研究竹子抗寒性生理提供了理论依据[18].2018年,测定了冷冻和冷敏感橄榄()品种,,基因表达,数据显示这些基因提高了橄榄品种的抗寒性,可作为筛选寒冷气候区域奶油橄榄品种的分子标记[19].近年来,已陆续从拟南芥、水稻()、甘蓝型油菜()等物种中成功克隆出FAD的编码基因[20-22].目前,FAD在植物油基因工程方面已取得一定进展,研究FAD编码基因对不同环境胁迫进行差异表达,获知FADs基因在不同胁迫下的原理和功能,可为不同物种遗传育种计划奠定基础.
4 植物多不饱和脂肪酸的生理功能
PUFAs是植物体细胞膜中的重要成分,具有流动性、柔韧性和选择透性.植物通过对PUFAs合成的调节从而对环境条件进行响应.近年来,在植物中对PUFAs生理功能的研究主要集中在对环境胁迫响应及抵抗功能方面,发现PUFAs影响植物的许多生理生化过程,包括响应干旱胁迫、冷胁迫、盐碱胁迫、重金属胁迫、紫外射线胁迫、离子通道调节、病原体防御等.
4.1 干旱胁迫响应
1985年起,谭裕模[23],王洪春[24]分别将干旱处理后与正常条件下的甘蔗()和玉米()叶片中的PUFAs进行对比,发现在干旱条件下,PUFAs含量的变化与植株的生长代谢密切相关.之后,分别对甘蓝型油菜[25]、豇豆()[26]等植株的叶片进行渐进式胁迫处理,发现在缺水条件下,2种植株叶片中PUFAs的含量均下降.2004年,研究报告了拟南芥在不同程度脱水条件下,PUFAs含量的稳定性对于细胞膜的水分亏缺耐受性十分重要[27].2018年,李东[28]等研究了在水稻种子萌发过程中ALA对萌发和幼苗生长过程中抗旱性的影响,结果表明,在不同浓度聚乙二醇干旱胁迫下,种子萌发和幼苗生长均受到抑制,而25,250 μmol/L的ALA可以缓解干旱对种子萌发和幼苗生长的抑制,且幼苗的根长、苗长、根质量、苗质量、种子酶活、幼根活力都显著提高.2022年,从柑橘()中成功克隆出基因,发现其异位过表达增加了PUFAs的含量,降低了角质层的通透性,增强了转基因植株的干旱耐受性[29].
4.2 冷胁迫响应
1973年,Lyons[30]提出了PUFAs含量与植物抗寒性相关的假说,通过外界的温度变化,生物膜调节膜脂中PUFAs的含量来判断植物的耐寒能力.1993年,在正常和低温条件下研究了拟南芥突变体的生长状况,发现在低温条件下植株的生长状态出现变化,组织中的PUFAs含量减少[31].随后人们将拟南芥中的ω-3脱氢酶在番茄()中表达,发现在转基因番茄中GLA含量增加且比野生型番茄有更强的耐寒性[32].2019年,在香蕉()中鉴定了1个MYB转录因子MaMYB4,发现MaMYB4间接调节ALA/LA比值的升高[33].2022年,对耐冷型和冷敏感型鹰嘴豆()进行低温处理,发现鹰嘴豆的2种植株的PUFAs含量都增加,抗性植株的PUFAs含量增加更显著[34].
4.3 盐碱胁迫响应
高盐度会影响植物光合作用、蛋白质合成、能量代谢等生物过程.2001年,在转基因聚球藻(sp. PCC 7942)中发现,膜脂PUFAs能增强其细胞膜的耐盐性[35].2005年,通过产生外源性过表达脂肪酸去饱和酶的烟草细胞指出了遭受盐胁迫的植物中亚麻酸水平下降,反映了损害机制[36].
2010年,研究了NaCl对格兰氏藻中脂肪酸含量的影响,得出PUFAs含量的增加是植株耐盐性的特征[37].
2023年,通过转录组和代谢组学综合分析了长期盐碱胁迫对水稻的影响,数据表明,LA代谢可以缓解高盐胁迫对细胞膜的损害[38].
4.4 重金属胁迫响应
Jemal[39]和Verdoni[40]等先后报道了在不同浓度重金属胁迫下,富含PUFAs的胡椒()幼苗和西红柿()幼苗分别对重金属的敏感性增强.2007年,Chaffiai[41]等在玉米幼苗中得出同样的结论.2013年,谌金吾[42]在考查镉、铅胁迫三叶鬼针草()的生理指标影响时发现,该植株中的PUFAs含量随着胁迫浓度的增加而增加,并在高浓度胁迫下维持高含量水平.2019年,采用气相色谱法研究了重金属胁迫下番茄叶片脂肪酸组成的变化,与对照植株相比,使用重金属处理的植株中ARA含量显著增加[43].2023年,分析揭示了马齿苋()对重金属胁迫的防御机制,发现在不同重金属胁迫下马齿苋茎部的PUFAs积累量均高于正常植株[44].
4.5 病原体防御
植物与外界环境接触时,其表皮蜡质会抵御外界胁迫.脂肪酸去饱和作用是植物防御反应中的一个重要组成部分,植物通过抗性基因产物识别病原体编码的Avr蛋白,从而对病原体感染作出反应.2000年,分离了马铃薯()块茎中形成的内源性脂氧合酶代谢产物来抵御巴氏丝核菌(Kühn)的伤害.结果表明,PUFAs内的脂氧合酶代谢产物介导防御反应[45].2013年,报道了C18不饱和脂肪酸合成的植物氧化脂质中的脂氧化合酶可将LA进行转化,这些化合物具有抗真菌活性[46].2020年,研究了拟南芥中PUFAs在坏死性病原菌灰霉菌()侵袭过程中的作用,拟南芥突变体对灰霉病菌表现出强抗性[47].
4.6 紫外射线胁迫响应
2019年,孙小琴[48]等通过研究紫外线辐射下小球藻()中PUFAs的含量变化,发现在该条件下小球藻品种的生长情况良好且PUFAs含量达到最大值.2015年,Liu[49]等发现在紫外线辐射下的巴普洛夫微藻()中的DHA和EPA的含量增加.
4.7 调节离子通道
PUFAs能够抑制或增强Ca2+,Na+,K+通道的活性,是电压门控离子通道的调节剂.1994年,报道了ARA和LNA是保卫细胞质膜内外离子电流的调节剂[50].2012年,Gutla[51]等研究报告了PUFAs强烈影响植物TPC1编码的慢液泡(Slow Vacuolar,SV)通道活性.这些研究结果都表明,PUFAs在调节离子通道活动方面起到重要作用.
5 结语
PUFAs是自然界中对氧最敏感的分子,以磷酸脂质和半乳糖脂质结合的形式存在于细胞和细胞器中,植物中FAD是由核基因编码的,主要使内质网和质体中的FAs去饱和,FAD基因的表达受到生物和非生物因子的诱导[52].有大量研究表明,FAD在植物育种和植物油基因工程等方面已取得一定进展.之前的育种工作者致力于提高物种产量和纤维质量,现在逐渐关注物种中油量比例、功能及营养特性.2016年,采用MABC(marker-assisted backcrossing)和MAS(marker-assisted selection)2种分子育种方法来提高花生油的品质,将苏油酸95R的2个突变等位基因转移到ICGV 06110,ICGV 6142,ICGV 6420遗传背景中,油酸含量增加1倍多,亚油酸含量减少一半,筛选出27个高油酸含量的渗入系,28个低油酸含量的渗入系,充分提高了花生油的品质[53].2022年,采用紫外线及氯化锂对深黄被孢霉()AS3.3410进行复合诱变,获得高产PUFAs突变菌株,具有较好的遗传稳定性且PUFAs含量显著提高[54].在植物中成功合成PUFAs之后,开始利用基因工程改良作物,在植物获得更多的PUFAs来弥补市场缺乏的鱼油资源,满足不同人群的需要.2016年,将芝麻()基因预测的叶绿体N端转运肽替换为内质网N端转运肽,并将KDELER保留信号置于C端,以引导产物随后定位于内质网,这样类似的分子策略在向日葵()和转基因烟草中都已做了测试,结果显示,在这些植物种子中的PUFAs含量都增加显著,为提高油料作物种子储存脂质的含量提供了很好的背景[55].2018年,报道了CRISPR/Cas9系统在甘蓝型油菜中的应用,获得2株FAD2_Aa等位基因突变的甘蓝型油菜转基因植株,有效提高了甘蓝型油菜中PUFAs的含量[56].在维管植物中,PUFAs对逆境胁迫作出响应,有大量数据证明,在非维管植物中PUFAs的作用机制还需进一步挖掘,为植物遗传育种提供更多的数据支撑.
[1] Iba K.Acclimative response to temperature stress in higher plants:approaches of gene engineering for temperature tolerance[J].Annual Review of Plant Biology,2002,53:225-245.
[2] 林晓娜.一氧化氮对桃果实脂氧合酶的调控作用[D].泰安:山东农业大学,2017.
[3] 魏雯雯,张波,徐昌杰,等.脂肪酸去饱和酶基因家族与果实成熟衰老研究进展[J].果树学报,2011,28(4):662-667.
[4] 王利民,符真珠,高杰,等.植物不饱和脂肪酸的生物合成及调控[J].基因组学与应用生物学,2020,39(1):254-258.
[5] 周霞.三角褐指藻多不饱和脂肪酸合成相关去饱和酶的功能研究[D].广州:暨南大学,2018.
[6] 刘华,张建涛,陈海燕,等.脂肪酸去饱和酶参与植物对胁迫的响应[J].东北农业大学学报,2013,44(1):154-160.
[7] Khotimchenko S V.Fatty acid composition of sevenspecies[J].Phytochemistry,1991,30(8):2639-2641.
[8] Miyazaki M,Ntambi J M.Fatty acid desaturation and chain elongation in mammals[M].Madison:Biochemistry of Lipids,Lipoproteins and Membranes Elsevier,2008:191-211.
[9] Mariamenatu A H,Abdu E M.Overconsumption of omega-6 polyunsaturated fatty acids(PUFAs)versus deficiency of omega-3 PUFAs in modern-day diets:the disturbing factor for their“balanced antagonistic metabolic functions”in the human body[J].Journal of Lipids,2021(Supply 1):8848161.
[10] 沈继红,石红旗,刘发义,等.耐盐碱植物:茵陈蒿种子营养成分共轭亚油酸的制备研究[J].中国海洋药物,2002(3):28-30.
[11] Heiduschka A,Luft K.Fatty oil from the seeds of the evening primrose,and a new linolenic acid[J].Archiv Der Pharmazie,1919,257:33-69.
[12] 刘婉君.微拟球藻多不饱和脂肪酸合成途径及去饱和酶功能的研究[D].广州:暨南大学,2016.
[13] 刘冬梅.三角褐指藻AA和EPA合成的Δ5脂肪酸去饱和酶研究[D].青岛:中国海洋大学,2011.
[14] Wada H,Murata N.Temperature-induced changes in the fatty acid composition of the CyanobacteriumPCC6803[J].Plant Physiology,1990,92(4):1062-1069.
[15] Reddy A S,Nuccio M L,Gross L M,et al.Isolation of a Δ6-desaturase gene from the cyanobacteriumsp. strain PCC6803 by gain-of-function expression insp. strain PCC 7120[J].1993,22:293-300.
[16] Im Y J,Han O,Chung G C,et al.Antisense expression of an Arabidopsis ω-3 fatty acid desaturase gene reduces salt/drought tolerance in transgenic tobacco plants[J].Molecules & Cells,2002,13(2):264-271.
[17] Zhang M,Barg R,Yin M,et al.Modulated fatty acid desaturation via overexpression of two distinctΔ-3 desaturases differentially alters tolerance to various abiotic stresses in transgenic tobacco cells and plants[J].The Plant Journal,2005,44(3):361-371.
[18] Zhang F,Zhu X Q,Guo Y L,et al.Ultrastructural changes and dynamic expressions of FAD7,Cu/Zn-SOD,and Mn-SOD inunder cold stress[J].Russian Journal of Plant Physiology,2014,61:760-767.
[19] Hashempour A,Ghasemnezhad M,Sohani M M,et al.Effects of freezing stress on the expression of fatty acid desaturase (,and)and beta-glucosidase(BGLC)genes in tolerant and sensitive olive cultivars[J].Russian Journal of Plant Physiology,2018,66:214-222.
[20] Van Cam N,Nakamura Y,Kanehara K.Membrane lipid polyunsaturation mediated by FATTY ACID DESATURASE 2 (FAD2)is involved in endoplasmic reticulum stress tolerance in[J].Plant Journal for Cell & Molecular Biology,2019,99(3):478-493.
[21] Birla D S,Malik K,Sainger M,et al.Progress and challenges in improving the nutritional quality of rice()[J].Critical Reviews in Food Science and Nutrition,2015,57(11):2455-2481.
[22] Xu L,Li W,Liu J,et al.Characteristics of membrane-bound fatty acid desaturase(FAD)genes inand their expressions under different cadmium and salinity stresses[J].Environmental and Experimental Botany,2019(162):144-156.
[23] 谭裕模.甘蔗叶片脂肪酸及透性与抗旱性的关系[J].福建农学院学报,1988,17(3):211-215.
[24] 王洪春.植物抗逆性与生物膜结构功能研究的进展[J].植物生理学通讯,1985(1):62-68,31.
[25] Dakhma W S,Zarrouk M,Cherif A.Effects of drought-stress on lipids in rape leaves[J].Phytochemistry,1995,40(5):1383-1386.
[26] Torres-Franklin M L,Repellin A,Huynh V B,et al.Omega-3 fatty acid desaturase(,,)gene expression and linolenic acid content in cowpea leaves submitted to drought and after rehydration[J].Environmental& Experimental Botany,2009,65(2/3):162-169.
[27] Agnes G,Ana-Rita M,Daniel L,et al.Effect of drought stress on lipid metabolism in the leaves of(ecotype Columbia)[J].Annals of Botany,2004(3):345-351.
[28] 李东,甄春燕,陈德富,等.-亚麻酸对干旱胁迫下水稻种子萌发的影响[J].广西植物,2018,38(8):1025-1031.
[29] Liu D,Guo W,Guo X,et al.Ectopic over expression of CsECR from navel orange increases cuticular wax accumulation in tomato and enhances its tolerance to drought stress[J].Frontiers in Plant Science,2022,13:924552.
[30] Lyons J M.Chilling Injury in Plants[J].Annual Review of Plant Biology Physiol,1973,24(1):445-466.
[31] Miquel M,James D,Dooner H,et al.requires polyunsaturated lipids for low-temperature survival[J].Proceedings of the National Academy of Sciences of the United States of America,1993,90(13):6208-6212.
[32] Nishiuchi T,Iba K.Roles of plastid ε-3 fatty acid desaturases in defense response of higher plants[J].Journal of Plant Research,1998,111(4):481-486.
[33] Chunbo S,Yingying Y,Tianwei Y,et al.MaMYB4 Recruits histone deacetylase MaHDA2 and modulates the expression of ω-3 fatty acid desaturase genes during cold stress response in banana fruit[J].Plant& Cell Physiology,2019,60(11):2410-2422.
[34] Javidi M R,Maali A R,Poormazaheri H,et al.Cold stress-induced changes in metabolism of carbonyl compounds and membrane fatty acid composition in chickpea[J].Plant Physiology and Biochemistry,2022,192:10-19.
[35] Allakhverdiev S I,Kinoshita M,Inaba M,et al.Unsaturated fatty acids in membrane lipids protect the photosynthetic machinery against salt-induced damage in1[J].Plant Physiology,2001,125(4):1842-1853.
[36] Zhang M,Barg R,Yin M,et al.Modulated fatty acid desaturation via overexpression of two distinct ε-3 desaturases differentially alters tolerance to various abiotic stresses in transgenic tobacco cells and plants[J].The Plant Journal,2005,44:361-371.
[37] Fujii S,Uenaka M,Nakayama S,et al.Effects of sodium chloride on the fatty acids composition in(Ochromonadales,Chrysophyceae)[J].Phycological Research,2010,49(1):73-77.
[38] Qian G,Wang M,Wang X,et al.Integrated transcriptome and metabolome analysis of rice leaves response to high saline-alkali stress[J].International Journal of Molecular Sciences,2023,24(4):4062.
[39] Jemal F,Zarrouk M,Ghorbal M H.Effect of cadmium on lipid composition of pepper[J].Biochemical Society Transactions,2000,28(6):907-910.
[40] Verdoni N,Mench M,Cassagne C,et al.Fatty acid composition of tomato leaves as biomarkers of metal-contaminated soils[J].Environmental Toxicology and Chemistry,2001,20(2):382-388.
[41] Chaffai R,Elhammadi M A,Seybou T N,et al.Altered fatty acid profile of polar lipids in maize seedlings in response to excess copper[J].Journal of Agronomy and Crop Science,2007,193(3):207-217.
[42] 谌金吾.三叶鬼针草(L.)对重金属Cd,Pb胁迫的响应与修复潜能研究[D].重庆:西南大学,2013.
[43] Kisa D.Responses of the fatty acid composition ofexposed to heavy metal stress[J].2019,9(1):88-96.
[44] Alsherif E A,Yaghoubi K M,Crecchio C,et al.Understanding the active mechanisms of plant()against heavy metal toxicity[J].Plants,2023,12(3):676.
[45] Reddy P S,Kumar T C,Reddy M N,et al.Differential formation of octadecadienoic acid and octadecatrienoic acid products in control and injured/infected potato tubers[J].BBA-Molecular and Cell Biology of Lipids,2000,1483(2):294-300.
[46] Kim K R,Oh D K.Production of hydroxy fatty acids by microbial fatty acid-hydroxylation enzymes[J].Biotechnology Advances,2013,31(8):1473-1485.
[47] Dubey O,Dubey S,Schnee S,et al.Plant surface metabolites as potent antifungal agents[J].Plant Physiology and Biochemistry,2020,150:39-48.
[48] 孙小琴,孙昕,李鹏飞,等.紫外辐射对小球藻光合性能及油脂积累的影响[J].中国油脂,2019,44(12):114-119.
[49] Liu S,Zhao Y,Liu L,et al.Improving cell growth and lipid accumulation in green microalgaesp. via UV irradiation[J].Applied Biochemistry and Biotechnology,2015,175:3507-3518.
[50] Lee Y,Lee H J,Crain R C,et al.Polyunsaturated fatty acids modulates stomatal aperture and two distinct K+channel currents in guard cells[J].Cellular Signalling,1994,6(2):181-186.
[51] Gutla P V K,Boccaccio A,Angeli A D,et al.Modulation of plant TPC channels by polyunsaturated fatty acids[J].Journal of Experimental Botany,2012,63(17):6187-6197.
[52] Berestovoy M A,Pavlenko O S,Goldenkova-Pavlova I V.Plant Fatty Acid Desaturases:Role in the Life of Plants and Biotechnological Potential[J].Biology Bulletin Reviews,2020,10(2):127-139.
[53] Janlia P,Pandey M K,Shasidhar Y,et al.Molecular breeding for introgression of fatty acid desaturase mutant alleles (and)enhances oil quality in high and low oil containing peanut genotypes[J].Plant Science,2016,242:203-213.
[54] 宋遥遥,卢晓霆,贾路遥.多不饱和脂肪酸高产菌株的诱变选育[J].中国酿造,2022,41(5):73-77.
[55] Bhunia R K,Chakraborty A,Kaur R,et al.Enhancement of-linolenic acid content in transgenic tobacco seeds by targeting a plastidial ω-3 fatty acid desaturase()gene of Sesamum indicum to ER[J].Plant Cell Reports,2016,35:213-226.
[56] Okuzaki A,Ogawa T,Koizuka C,et al.CRISPR/Cas9-mediated genome editing of the fatty acid desaturase 2 gene in[J].Plant Physiology and Biochemistry,2018,131:63-69.
Research progress of plant polyunsaturated fatty acids
CAI Can,BAI Yu,HAN Yi,GUO Jiaxin,SHA Wei,ZHANG Meijuan,PENG Yifang,MA Tianyi
(1. School of Life Sciences,Agriculture and Forestry,2. Heilongjiang Provincial Key Laboratory of Resistance Gene Engineering and Protection of Biodiversity in Cold Areas,Qiqihar University,Qiqihar 161006,China)
Polyunsaturated fatty acids(PUFAs)are a particular class of bioactive substances,which regulate growth,development and external stresses resistance progresses of plants.Under the actions of a series of desaturases and elongases,polyunsaturated fatty acids form compounds with different structures and functions through different metabolic pathways,which the compounds act as signal molecules regulating different biological functions and the expression of related genes,so as to enhance the resistance of plants to different environmental stresses.The sources,classification,synthetic pathways of polyunsaturated fatty acids in plants and their roles in resisting external stresses were introduced,which laid the foundation for further studies of polyunsaturated fatty acids′ functions in plants to some extents.
polyunsaturated fatty acid;fatty acid desaturase;environmental stress
1007-9831(2023)09-0064-06
Q943
A
10.3969/j.issn.1007-9831.2023.09.014
2023-05-03
齐齐哈尔大学学位与研究生教育教学改革项目(JGXM_QUC_2021009);黑龙江省省属高等学校基本科研业务费科研项目青年创新人才项目(145109212);黑龙江省省属高等学校基本科研业务费科研项目(145109311)
蔡璨(2000-),女,甘肃金昌人,在读硕士研究生,从事植物逆境分子遗传学研究.E-mail:1549920836@qq.com
马天意(1989-),男,黑龙江齐齐哈尔人,副教授,博士,从事植物逆境分子遗传学研究.E-mail:tyma@qqhru.edu.cn
