表油菜素内酯对热胁迫下‘美乐’葡萄着色和抗氧化特性的影响
2023-09-21李嘉佳王磊曹玉早王世平
李嘉佳,王磊,曹玉早,王世平*
(1. 上海交通大学农业与生物学院,上海 200240;2. 东海县石梁河镇金朵种植家庭农场,江苏连云港 222000)
温度是影响植物新陈代谢等生理功能的主要因素之一,在适宜的温度下,植物组织中的不同蛋白质会堆叠成特定的形状,行使不同功能[1]。极端的温度变化会导致具有功能的蛋白质变性失活,最终造成植物新陈代谢速率下降,果实产量及品质降低[2]。据报道,每年全球农作物产量损失有超过半数都是由极端温度造成的, 随着全球气候的日益变暖,温室效应逐渐加剧,高温对农作物的危害更加严重[3]。因此,如何减轻极端气候对植物的伤害,实现高产优质栽培一直是学者们追求的目标。
‘美乐’葡萄是世界范围内种植最广泛的品种之一,具有早熟、多产、口感细腻等优良性状,可用来酿制美味而柔滑的葡萄酒,也可平衡其他葡萄酒的酸涩,极大提高葡萄酒的品质,获得更高的市场效应[4-5]。但是‘美乐’耐热性较差,夏季的高温也成为制约其生长发育最主要的环境因子,若长期处于高温环境下,会抑制树体光合作用,延缓果皮着色,酿制的酒口感变劣,品质变差,影响了该品种的的应用[6]。喷施外源植物激素是快速提高植株抗逆性,缓解热胁迫对细胞膜伤害的有效措施[7-8]。脱落酸(ABA)是促进果皮着色和果实糖分积累的最重要的内源植物激素。已有研究表明,向转色前葡萄果实喷施一定浓度的ABA和乙烯类物质(乙烯利)可以显著缓解高温胁迫下葡萄果实的上色困难,固酸比下降等问题[9-11]。油菜素内酯(BR)具有在极低浓度下促进植株营养生长的功效,并且对果实膨大、糖分积累、果皮着色方面也有显著的促进作用[12]。但遗憾的是,对外源BR及24-表油菜素内酯(EBR)能否通过调控内源BR的合成来缓解热胁迫下‘美乐’葡萄上色困难,从而提高果实品质方面还未有系统的研究,分子机制也仍待进一步揭示。
本研究以盆栽两年生的‘美乐’葡萄为试材,探究EBR对热胁迫下果实次生代谢物合成及抗氧化能力的影响,着重聚焦EBR是否会通过影响内源BR的合成来缓解热胁迫下葡萄果皮上色困难的问题,以期在未来葡萄栽培过程中通过转色前果实浸泡EBR的方式来提高葡萄抗热胁迫的能力,恢复受热胁迫伤害的表型,进而整体提高酿酒葡萄的品质。
1 材料与方法
1.1 材料准备及处理
2022年3月5日将两年生‘美乐’植株移栽于10 L栽培容器中,挑选长势相近且无病虫害的植株,置于上海交通大学农业与生物学院玻璃温室中,栽培土壤由园土和蛭石按1∶1比例混合而成,土壤肥水和病虫害管理一致。待果实转色前2周左右,挑选长势健壮的9株进行试验,调整果穗数和叶片数,使总果粒数与总叶片数比介于1.2~1.5,供后续处理。在果实转色前1周,参考预备试验筛选的处理温度,设置对照(Control,25 ℃/12 h,25 ℃/12 h),热胁迫(HT,50 ℃/12 h,25 ℃/12 h)及热胁迫 + EBR(HT+EBR,50 ℃/12 h,25 ℃/12 h)3组处理,不间断处理14 d。每个处理使用3株冬季修剪完毕的盆栽葡萄,每株保留3~4穗果,每穗保留60粒,并定期浇灌霍格兰营养液。此外,EBR处理在首次高温胁迫后6 h进行,将未转色的葡萄整穗果浸泡在2 L的0.5 mg·L-1EBR溶液(现用现配),每穗浸泡时间持续3 min,待果穗完全没有EBR溶液滴落后,再重复该步骤一次。
1.2 样品采集及指标测定
本试验共设置5个采样时间点,分别为DAT0(处理后0 d,HT及EBR处理后6 h进行采样)、DAT3(处理后3 d,转色前期)、DAT7(处理后7 d,转色中期)、DAT11(处理后11 d,转色完成)、DAT15(处理后15 d,成熟期)。在每个采样时间点,随机在每穗葡萄上中下部各采集3粒,每个处理采果数量不低于20粒。在测定完果实基本品质指标后(粒质量、果实纵径、果实横径、可溶性固形物、可滴定酸),所有果实经液氮冷冻后置于-80 ℃冰箱,用于后续果实花色苷、酚类等次生代谢产物、抗氧化酶活性的测定及RNA的提取。
1.2.1 酚类物质的测定
随机选择5粒完整果实,用液氮研磨成粉末后,称取0.10 g,加入1 mL pH 7.8的PBS缓冲液,4 ℃、12000 r·min-1下离心10 min,取上清液进行果实类黄酮、总酚、抗坏血酸、白藜芦醇的测定,使用由北京索莱宝生物科技有限公司生产的Elisa试剂盒进行检测。每处理重复3次。
1.2.2 抗氧化酶活性的测定
取0.10 g冻干葡萄粉末于1 mL pH 7.8的PBS缓冲液中进行冰浴匀浆处理,4 ℃、12000 r·min-1下离心10 min,取上清液进行果实内超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)和苯丙氨酸解氨酶(PAL)活性的测定,使用北京索莱宝生物科技有限公司生产的Elisa试剂盒进行检测。每处理重复3次。
1.2.3 总花色苷的测定
先将葡萄果实研磨成粉末,准确称取0.10 g,加入6 mL含1%HCl的甲醇溶液,涡旋振荡混匀2 min,4 ℃避光浸提过夜。在加入4 mL ddH2O后,将溶液在3800 r·min-1下离心10 min。收集上清液并加入1 mL氯仿,混匀,然后将混合物在13000 r·min-1下离心5 min。最后,使用分光光度计在530 nm和657 nm下分别测量水相中的花色苷含量,并使用等式 (A530-A657)·g-1得到总花色苷含量。每个处理重复3次。
1.2.4 内源BR含量的测定
取0.50 g果肉组织粉末于5 mL pH 7.8的PBS缓冲液以及5 mL的1 mol Tris-HCl中进行冰浴匀浆处理,经过4 ℃、6000 r·min-1离心15 min 后,取上清液进行果实内BR含量的测定,使用上海研启生物科技有限公司生产的Elisa试剂盒进行检测。每个处理重复3次。
1.2.5 qRT-PCR
RNA提取试剂盒(TaKaRa,大连,中国)用于提取在不同处理的不同采样期的葡萄果实中的总RNA。BIO-RADXR凝胶成像分析系统(美国,加利福尼亚州,BIO-Rad)用于检测提取的RNA的纯度和完整性。使用CFX connect实时PCR检测系统(Bio-Rad,CA,USA)进行qRT-PCR,并遵循以下既定程序:95 ℃(20 s),然后95 ℃(39个循环,15 s),55 ℃(15 s),最后60 ℃(15 s)。基因的相对表达由2-△△Ct表示。所有基因和转录因子序列均从EnsemblPlants(http://plants.ensembl.org/info/about/index.html)获得。 在Primer Premier 5.0和qPrimerDB qPCR引物数据库中设计用于qRT-PCR的引物(https://biodb.swu.edu.cn/qprimerdb/)。
1.3 数据分析
使用SPSS 16.0统计软件包(IBM,Armonk,NY,USA)对本研究中获得的结果进行差异显著性分析。方差齐性分析采用均数±标准差(SE)形式,选择单变量方差分析(one-way ANOVA,P<0.05),每个处理至少选择3组数据。所有图形均使用GraphPad Prism 9.0(GraphPad Software Inc.,圣地亚哥,美国),TBtools(CAN,CHN)和Visio 2020(Microsoft,SEA,美国)绘制。
2 结果与分析
2.1 EBR处理对热胁迫下果实大小及着色的影响
图1为热胁迫及EBR处理对果粒表型及果粒大小的影响。结果表明,热胁迫显著抑制了果粒的膨大和着色,EBR处理后果粒大小和上色程度显著改善,且果实纵横径、粒质量均在转色完成期和果实成熟期高于对照组(图1A、B、C)。以上表型以及果粒大小指标说明,热胁迫可以抑制‘美乐’葡萄的着色及膨大,EBR对恢复果实表型,促进果实膨大有着积极效应。
2.2 EBR处理对热胁迫下果实品质的影响
图2为热胁迫及EBR处理对果实糖积累、酸代谢和多种次生代谢产物合成的影响。具体来说,热胁迫显著抑制了果实DAT3、DAT7、DAT11、DAT15时期的可溶性固形物的积累。在DAT3和DAT15时期,EBR浸泡果实后则显著抑制了可溶性固形物在热胁迫下的减少,但其值仍显著低于对照组(图2A)。此外,热胁迫也可以显著的降低葡萄果实内可滴定酸的含量,EBR浸泡可恢复热胁迫下其值的降低,但其含量仍显著低于对照组(图2B)。对于类黄酮含量以及总酚含量(图2C、2D),热胁迫处理同样显著抑制了这两种物质的积累。在EBR处理组中,总酚含量和类黄酮的含量较热胁迫处理组显著提升。此外,EBR对于缓解热胁迫处理下抗坏血酸和白藜芦醇这两种次生代谢产物含量降低效果显著,尤其在转色期和成熟期(图2E、2F)。
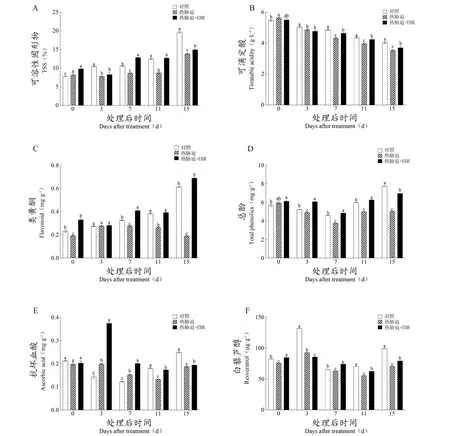
图2 EBR对‘美乐’葡萄果实多种次生代谢物含量的影响Figure 2 Effect of EBR on the content of multiple secondary metabolites in 'Merlot' grape
2.3 EBR处理对热胁迫下果实抗氧化能力的影响
如图3所示,SOD、POD、CAT、PAL活性变化一致,呈现出总体先上升后下降的趋势,热胁迫显著促进了果实DAT0和DAT15时期SOD活性的提高,显著促进几乎所有采样时期POD和CAT的活性,而抑制了果实转色期PAL的活性。此外,EBR在促进热胁迫下DAT3、DAT7和DAT11时期的SOD、POD以及所有采样时期PAL的活性提高方面发挥着重要作用。此外,EBR较热胁迫处理组显著降低了热胁迫下CAT活性。

图3 EBR对‘美乐’葡萄果实抗氧化物质积累的影响Figure 3 Effect of EBR on the accumulation of antioxidant substances in 'Merlot' grape fruit
进一步在转录水平上探究EBR对促进热胁迫下抗氧化相关基因表达的作用,如图4所示。SOD,POD、CAT和PAL1的表达总体呈现出先上升后下降的趋势,以及热休克基因HSFA1、HSP70呈现出总体不断下降的趋势。在DAT7、DAT11和DAT15时期,热胁迫显著促进了SOD、POD、CAT的表达,抑制了PAL1的表达。HSFA1和HSP70在DAT0和DAT15时期明显上调,其余时间未见显著差异,且EBR处理并不能促进这些基因在热胁迫下表达的显著提高。
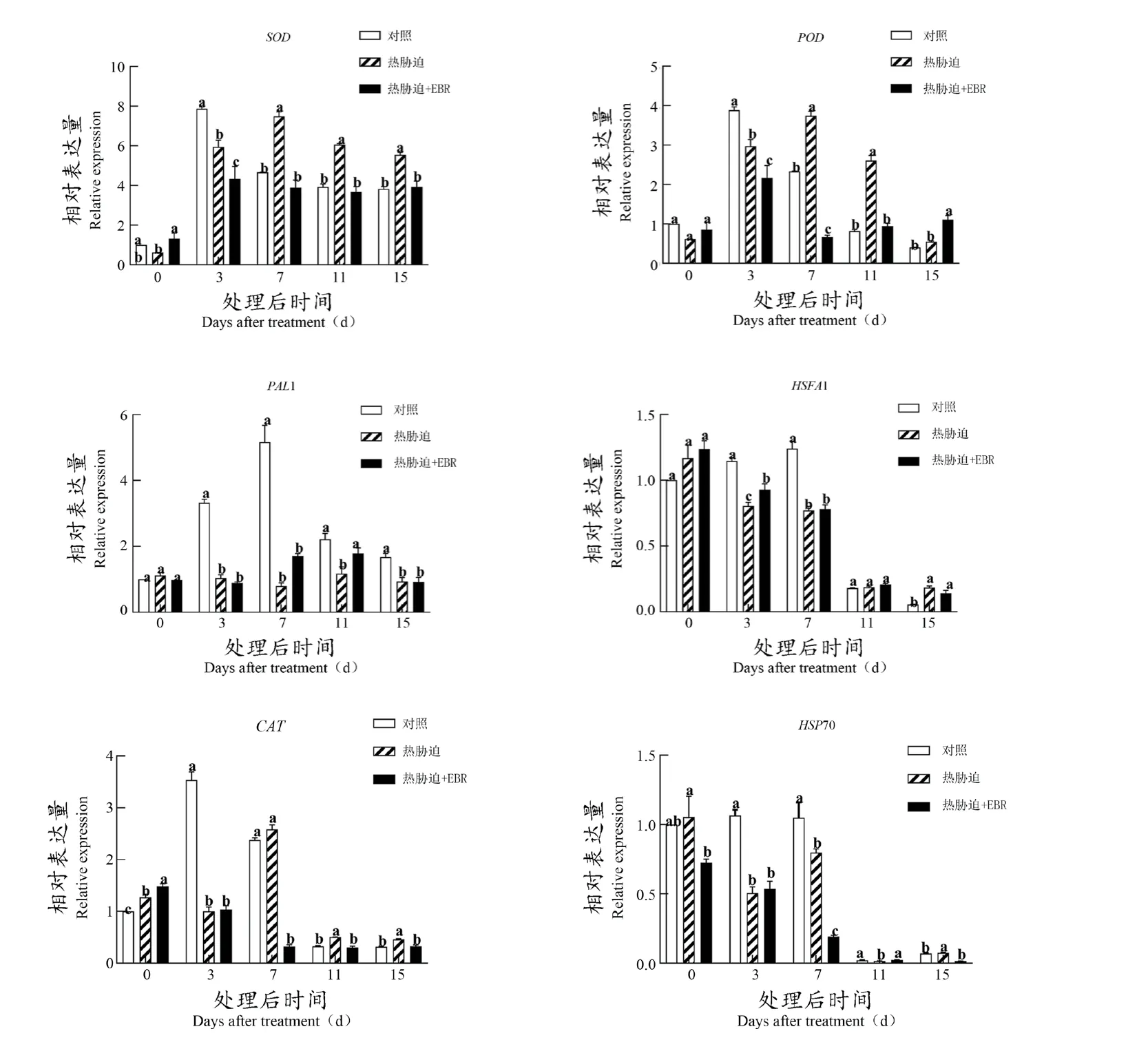
图4 EBR对‘美乐’葡萄果实抗氧化物质相关基因表达量的影响Figure 4 Effect of EBR on the expression of related genes of antioxidant substances in 'Merlot' grape fruit
2.4 EBR处理对热胁迫下果实花色苷含量及其合成代谢及信号转导的影响
如图5A所示,随着果实的不断成熟,总花色苷含量不断提高,热胁迫处理在转色期和成熟期时期均显著抑制了总花色苷的积累,EBR有效地逆转热胁迫对花色苷积累的抑制作用。
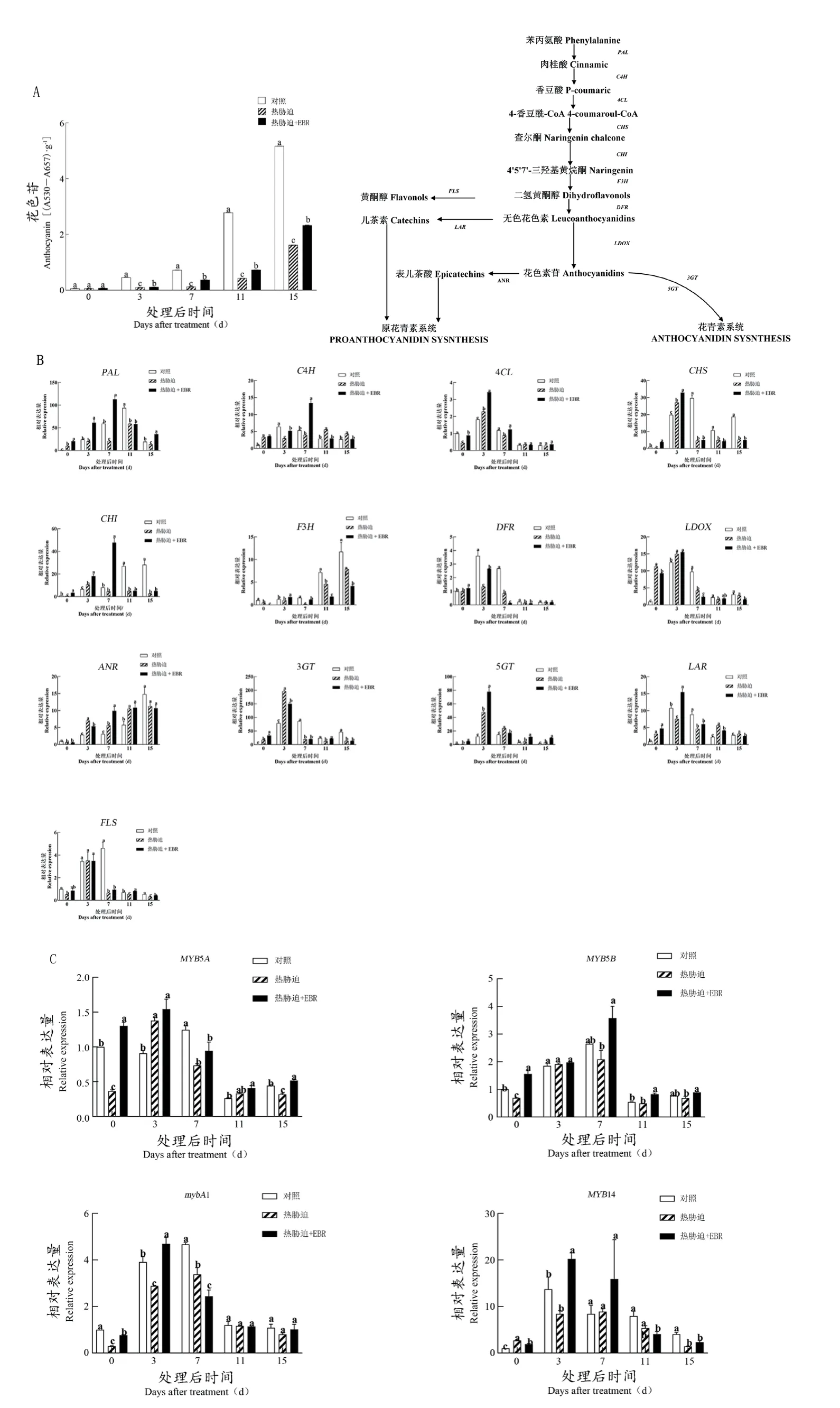
图5 EBR对‘美乐’葡萄果皮花色苷含量及花色苷合成代谢、信号转导的影响Figure 5 Effect of EBR on anthocyanin content, anthocyanin biosynthesis and metabolism, and signal transduction of 'Merlot' grape
进一步在转录水平上研究热胁迫及EBR处理对‘美乐’葡萄花色苷合成代谢及信号转导的影响,如图5B所示。结果发现,花色苷合成的上游基因PAL、C4H、CHS、CHI变化趋势较为一致,即随着果实成熟,它们的表达量先增加后减少。随着葡萄果实的不断成熟,负责调控花色苷合成的下游基因的F3H不断上调,下游基因LDOX和DFR先上调后下调。此外,在转色期DAT15时期,热胁迫较对照组降低了上述大部分基因的表达。而在DAT0后及DAT3时期,EBR对热胁迫下这些基因表达量的上调起促进作用。在大部分的采样时间点,热胁迫促进了5GT的表达,而抑制了3GT的表达,EBR在显著上调它们的表达方面做出贡献。同时发现,负责调控原花青素(儿茶素)合成的基因LAR呈现先上升后下降的趋势,以及负责调控原花青素(表儿茶素)的基因ANR随着果实的成熟呈现上调的趋势,且在大多数采样时间,热胁迫处理均可以促进它们的表达。经过EBR浸泡果实后,DAT3时期的LAR、DAT7时期的ANR上调趋势更显著。此外,诱导二氢黄酮醇生成黄酮醇的基因FLS的表达量先上升后下降;在大部分采样时间,热胁迫处理显著下调了它的表达量;在EBR浸泡果实后,表达量虽上调,但整体水平仍低于对照组。
对与花色苷信号转导密切相关的4个转录因子的表达进行测定(图5C)。结果表明,MYB5A、MYB5B、mybA1、MYB14有一致的变化趋势,呈现先上升后下降。在大多数的采样时间点,热胁迫下调了这些基因的表达,EBR则有效地逆转了热胁迫对它们表达的抑制作用,尤其是在DAT3和DAT7。
2.5 EBR处理对热胁迫下果实内源BR含量及其合成代谢的影响
如图6所示,葡萄果实内源BR含量随着果实成熟而不断增加,在DAT7和DAT15时期,热胁迫显著提高了内源BR的含量。而在大部分的采样点,EBR处理组相较于热胁迫处理组,内源BR含量无明显提高,但其含量仍显著高于对照组。
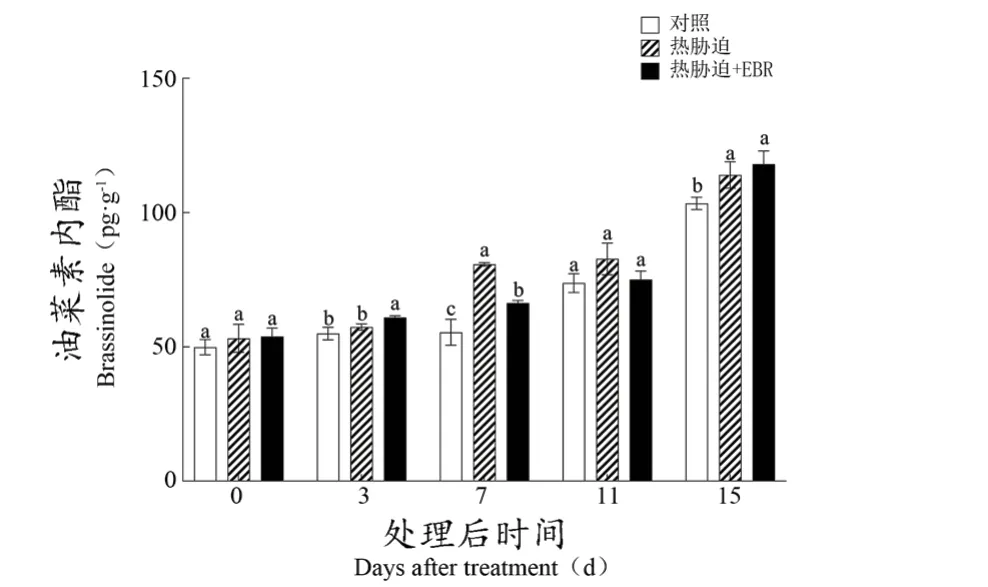
图6 EBR对‘美乐’葡萄果实内源BR含量的影响Figure 6 Effect of EBR on endogenous BR content of 'Merlot' grape
进一步在转录水平上分析热胁迫和EBR对BR合成代谢关键基因表达的影响。如图7所示,上游基因DWF4和CPD表现出较为一致的变化趋势,在大多数采样时间点,热胁迫都显著提高了它们的表达量,且在果实DAT11和DAT15时期,EBR对于上调这两个基因表达量的作用效果也是显著的。而BR合成上游基因DET2则表现出先略微下降,后逐步提升的趋势。在大多数采样点,热胁迫下它的表达都显著高于对照组,在果实DAT0和DAT11时期,EBR显著上调它的表达。下游基因ROT3,BR6OX1,BR6OX2的表达量在热胁迫处理组和EBR处理组的转色期较对照组上调。调控BR代谢的BAS1会随着果实不断成熟先上调后下调,在大多数的采样时间点,热胁迫显著上调了其表达,而EBR处理在绝大多数的采样时期较对照组上调了它们的表达。
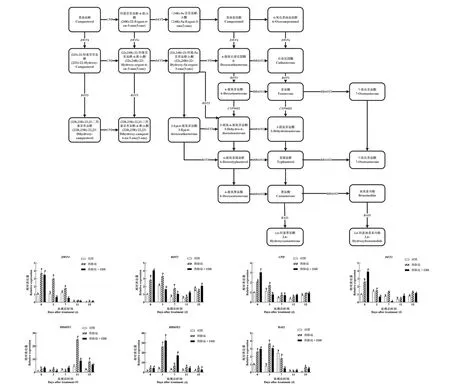
图7 EBR对‘美乐’葡萄果实内源BR合成代谢的影响Figure 7 Effect of EBR on endogenous BR biosynthesis of 'Merlot' grape fruit
2.6 内源BR的合成与缓解热胁迫下果实品质降低的关联程度
通过相关性试验进一步分析发现,随着‘美乐’葡萄果实的不断发育成熟,内源BR的积累与果实的膨大,糖分的积累和花色苷的富集关系最为密切(图8)。令人意外的是,内源BR的积累与多种抗氧化物质的积累呈负相关,遂推测EBR缓解了热胁迫对葡萄果实的伤害,提高其抗氧化能力并不是通过促进内源BR合成这条途径完成的,具体的验证仍待进一步开展。
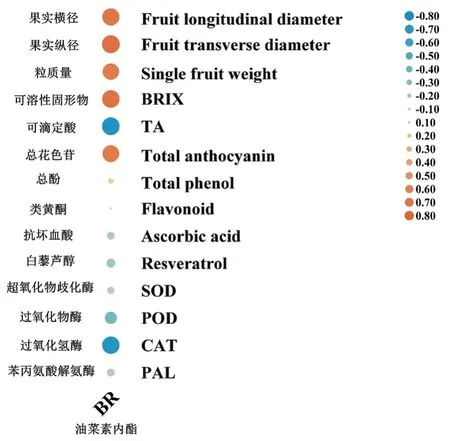
图8 内源BR与品质相关指标的关联程度Figure 8 Correlation between endogenous BR and quality-related parameters
3 讨论与结论
热胁迫亦称高温胁迫,是指环境温度升高到植物适宜温度以上,对植物的能量代谢、生长发育产生的胁迫现象[13]。随着温室效应的日趋严重,热胁迫是葡萄植株生长发育面临的最大问题之一,轻则会导致植株的氧化受伤,代谢失调,严重时会影响植株的生长发育及果实品质[3]。综合本试验结果可知,热胁迫确实会显著抑制酿酒葡萄‘美乐’的果实膨大、果皮着色、糖分积累、抗氧化酶活性提高。EBR的应用可以通过促进与花色苷合成密切相关的结构基因以及转录因子的表达,减轻热胁迫造成的果皮着色困难的程度,通过上调部分抗氧化基因的表达量,缓解热胁迫对细胞膜造成的伤害,提高抗氧化能力。此外,EBR促进果实内源BR的积累,上调负责内源BR合成的大部分下游基因的表达,以应对葡萄果实代谢失衡、品质下降的现象。最后,相关性分析进一步佐证了我们的推测:BR与葡萄果实多种次生代谢产物的合成密切相关,尤其体现在果实膨大、果皮着色与糖分积累方面。
首先,高温主要引起生物膜的物理化学状态和蛋白质分子构型的可逆变化,由于类囊体膜对热特别敏感,光合作用失调是热胁迫的最初指标[14]。高温造成植物光合作用受到抑制,最后导致细胞和个体的死亡[15]。前人研究证实,EBR处理葡萄叶片后,受热胁迫(42 ℃)损害严重的叶片表型慢慢恢复,光合速率提高,并伴随着抗氧化相关的SOD、POD、CAT等物质含量的增加,植株对抗热胁迫的能力显著增强[16]。本试验结果表明,热胁迫处理下果实内的多种次生代谢产物(类黄酮、总酚、白藜芦醇)含量显著降低,果实内与抗逆能力相关的SOD、POD及CAT含量在一定时期内显著增加,果实清除自由基的能力相应提高。EBR对于减轻果实品质下降及抗氧化能力提高方面发挥着重要作用,在果实转色期和成熟期,EBR显著促进了热胁迫下可溶性固性物的积累。在转色完成和成熟期,EBR减轻了热胁迫果实内抗坏血酸和白藜芦醇减少的表型。在大部分采样时期,EBR进一步促进了葡萄果实内SOD和POD的积累。此外,我们发现EBR对促进总酚以及类黄酮的积累在转色期和成熟期较明显。基于此,推测热胁迫处理加速了转色期和成熟期‘美乐’葡萄膜脂过氧化,且与清除自由基相关的抗氧化酶活性也相应提高。EBR不仅促进了多种反映果实品质的次生代谢产物的合成,而且更进一步的加速了多种抗氧化物质的富集,虽然与抗氧化能力提高有关基因的表达量较热胁迫处理组没有明显的上调,但热胁迫对果实表型及品质的损害还是在EBR处理下有所减轻。
其次,高温必然会引起植物内源激素含量的变化[17],诸如脱落酸(ABA)和茉莉酸(JA)这两种已被广泛报道与抗逆性提高相关的植物激素,它们的含量在经过热胁迫后会显著提高[18-20]。而与植株生长发育相关的生长素(IAA)和细胞分裂素(CTK)在经过热胁迫后会显著降低[21-22]。虽然BR也被证实与提高植株抗逆性密切相关,但由于BR在果实内含量极低,很少有研究系统地讨论EBR对热胁迫处理后葡萄内源BR含量变化的影响。本试验结果证实了热胁迫处理的确可以显著提高大部分采样时期果实内源BR的含量。EBR处理过后,在转色前期和成熟期,内源BR含量较热胁迫处理组提高,但两个处理组并没有显著的差异。进一步在转录水平上研究热胁迫和EBR处理对内源BR合成代谢关键基因表达量的影响发现,热胁迫促进了BR合成最关键的两个下游基因(BR6OX1与BR6XO2)在转色完成和成熟期的表达,且促进了BR代谢相关基因(BAS1)表达量在大部分采样时期的上调,推测虽然BR合成和分解的速率在经过热胁迫后都有了提高,但是整体上BR合成的速率还是要快于BR分解的速率,最终导致热胁迫促进葡萄成熟期果实内源BR积累的表型。
众所周知,葡萄果皮着色困难是热胁迫后最明显的表型之一,持续高温会严重阻碍多种有机营养物质的积累,从而造成果皮花色苷含量的减少[23-25]。本研究结果不仅从表型上提供了热胁迫严重阻碍‘美乐’葡萄果皮着色的理论依据,而且也得出了经过热胁迫后,果皮花色苷在转色期和成熟期显著降低的结论,并且在转录水平上进一步解释了这个现象,即在果实转色期和成熟期,热胁迫显著抑制了花青素合成下游基因3GT的表达,而促进了原花青素合成关键基因ANR和LAR的表达,EBR的应用则可以有效缓解大部分时期热胁迫对葡萄着色的抑制作用。根据上述结果推测,热胁迫会更倾向促进原花青素的合成,而抑制与果皮着色有关的花青素的合成,最终产生相较对照组着色不良的表型,而EBR则可以通过上调大部分与花青素合成相关的基因来缓解热胁迫处理造成果皮着色困难的现象。
综上所述,本试验在转色前对酿酒葡萄‘美乐’实施热胁迫处理(最高54 ℃,最低46 ℃,平均50 ℃)基础上,并于当天用EBR浸泡葡萄果穗,发现热胁迫显著抑制葡萄膨大、糖分积累、果皮着色及多种抗氧化酶活性增强。EBR处理整体上通过促进多种次生代谢产物的积累缓解了热胁迫对葡萄着色的抑制,通过提高几种抗氧化酶的活性减轻热胁迫对葡萄果实细胞膜的伤害。此外,我们推测EBR可通过调控内源BR的合成促进果皮糖及花色苷的积累,改善热胁迫下果皮着色困难的情况,但具体的分子机制仍待进一步探索。
