miR-409-3p在肝癌HepG2细胞中的表达及其在细胞增殖中的作用机制
2023-04-29王长青陈玲徐萍朱晓娟刘政
王长青 陈玲 徐萍 朱晓娟 刘政
摘要:目的本研究旨在明確miR-409-3p在肝癌细胞系中的表达及其意义,并探讨可能的分子机制。方法利用实时荧光定量PCR的方法检测miR-409-3p在正常肝细胞LO2,以及HepG2、BEL-7402、SMMC-7721、MHCC-97H四种肝癌细胞中的表达差异。阳离子脂质体法将miR-409-3p mimics及microRNA mimics control瞬时转染至肝癌细胞株HepG2中,利用CCK8法、平板克隆、流式细胞术检测miR-409-3p对体外癌细胞增殖、细胞周期及凋亡的影响。Western Blot检测过表达miR-409-3p的HepG2细胞中c-Met蛋白的表达变化,荧光素酶报告系统鉴定靶向关系。计量资料两组间比较采用成组t检验,多组间比较采用单因素方差分析,进一步两两比较采用SNK法。结果基于TCGA肝癌microRNA表达谱数据,肝癌组织中miR-409-3p的表达水平显著低于癌旁组织(t=7.752,P<0.05)。与在LO2中的表达水平相比,miR-409-3p在HepG2、SMMC-7721、MHCC-97H、BEL-7402中的表达水平均明显降低(F=31.043,P<0.05)。与miR-con组相比,转染miR-409-3p mimics后的HepG2细胞中miR-409-3p的表达水平上升(t=-8.836,P<0.05),说明干扰有效。CCK8实验结果显示,与miR-con组相比,转染miR-409-3p mimics后的HepG2细胞增殖能力在48、72、96 h明显减弱,差异均具有统计学意义(t值分别为2.876、3.359、3.707,P值均<0.05)。平板克隆形成实验显示,miR-409-3p mimics组的细胞克隆形成率明显低于miR-con组(t=2.846,P=0.047)。流式细胞术结果显示,与miR-con组相比,过表达miR-409-3p后导致HepG2细胞G2期细胞数增多,差异具有统计学意义(t=-3.763,P<0.05);而凋亡率无明显统计学差异(t=0.714,P=0.515)。荧光素酶报告系统鉴定结果显示c-Met为miR-409-3p的靶基因(t=4.970,P=0.007)。与miR-con组相比,转染miR-409-3p mimics的HepG2细胞中,c-Met蛋白表达水平下降(t=-8.509,P=0.001)。结论miR-409-3p通过抑制c-Met蛋白表达,进而调控下游信号通路引起细胞周期G2期阻滞,从而抑制肝癌HepG2细胞的增殖。关键词:癌, 肝细胞; 微RNAs; 原癌基因蛋白质c-met; 细胞增殖
Expression of miR-409-3p in hepatoma HepG2 cells and its mechanism in cell proliferation
WANG Changqing CHEN Ling XU Ping ZHU Xiaojuan LIU Zheng(1. Department of Gastroenterology, Yancheng First Peoples Hospital, Yancheng, Jiangsu 224008, China; 2. Department of Gastroenterology, Nanjing Pukou Peoples Hospital, Nanjing 211899, China; 3. Medical Center for Digestive Diseases, The Second Affiliated Hospital of Nanjing Medical University, Nanjing 210011, China)
Corresponding author:LIU Zheng, liuzheng117@126.com (ORCID:0000-0002-2195-4538)
Abstract:ObjectiveTo investigate the expression and significance of miR-409-3p in hepatoma carcinoma cell lines and possible molecular mechanism. MethodsQuantitative real-time PCR was used to measure the expression of miR-409-3p in normal LO2 hepatocytes and four hepatoma cell line (HepG2, BEL-7402, SMMC-7721, and MHCC-97H). Hepatoma HepG2 cells were transiently transfected with miR-409-3p mimics and microRNA mimics control using the cationic liposome method, and then CCK8, plate colony formation assay, and flow cytometry were used to observe the effect of miR-409-3p on the proliferation, cell cycle, and apoptosis of hepatoma cells in vitro. Western blot was used to measure the change in the expression of c-Met protein in HepG2 cells with overexpression of miR-409-3p, and luciferase reporter gene assay was used to identify targeting relationship. The independent-samples t test was used for comparison between two groups, and a one-way analysis of variance was used for comparison between multiple groups, followed by the SNK test. ResultsBased on the TCGA microRNA expression profile data of liver cancer, the expression level of miR-409-3p in liver cancer tissue was significantly lower than that in adjacent tissue (t=7.752, P<0.05). Compared with the LO2 cells, the HepG2, SMMC-7721, MHCC-97H, and BEL-7402 cells had a significant reduction in the expression level of miR-409-3p (F=31.043, P<0.05). Compared with the miR-con group, the HepG2 cells transfected with miR-409-3p mimics had a significant increase in the expression level of miR-409-3p (t=-8.836, P<0.05), suggesting that the interference was effective. CCK8 assay showed that compared with the miR-con group, the HepG2 cells transfected with miR-409-3p mimics had a significant reduction in proliferative capacity at 48, 72, and 96 hours (t=2.876, 3.359, and 3.707, all P<0.05). Plate colony formation assay showed that the miR-409-3p mimics group had a significantly lower plating efficiency than the miR-con group (t=2.846, P=0.047). Flow cytometry showed that compared with the miR-con group, overexpression of miR-409-3p resulted in the increased number of HepG2 cells in G2 phase (t=-3.763, P<0.05), while there was no significant difference in apoptosis rate (t=0.714, P=0.515). Luciferase reporter gene assay showed that c-Met was a target gene of miR-409-3p (t=4.970, P=0.007). Compared with the miR-con group, the HepG2 cells transfected with miR-409-3p mimics had a significant reduction in the expression of c-Met protein (t=-8.509, P=0.001). ConclusionBy inhibiting the protein expression of c-Met, miR-409-3p regulates downstream signaling pathways to induce cell cycle arrest in G2 phase and thus inhibits the proliferation of hepatoma HepG2 cells.
Key words:Carcinoma, Hepatocellular; MicroRNAs; Proto-Oncogene Proteins c-met; Cell Proliferation
肝细胞癌(HCC)的发生及发展包括一系列复杂的因素及步骤,涉及多个信号通路,探索HCC发生发展的分子机制,将有助于寻找其早期诊断的生物标志物及特异性的靶向药物[1]。c-Met作为一种原癌基因于20世纪80年代被发现,其所编码的c-Met蛋白是受体酪氨酸激酶家族成员[2]。研究[3]表明,在多种肿瘤中c-Met呈过表达,如肝癌、胃癌、结直肠癌、胰腺癌、肺癌、前列腺癌及乳腺癌等。本课题组前期以c-Met为研究对象,通过相关靶向治疗方式抑制肝癌细胞的生长[4-5]。近年来相关研究[6-7]陆续揭示,miR-409-3p可通过下调c-Met信号通路发挥其抑癌基因作用。迄今为止,HCC中有关miR-409-3p对c-Met调控的相关研究未见报道。因此本实验旨在研究miR-409-3p在HCC发生发展中的作用及其与c-Met相关的分子机制,期待从分子水平进一步探讨HCC发生发展的机制,并以此为HCC的临床诊断及靶向治疗提供理论基础。
1材料与方法1.1材料与试剂
1.1.1细胞株人正常肝细胞株LO2以及肝癌细胞株HepG2、BEL-7402、SMMC-7721、MHCC-97H均来自本实验室保存。
1.1.2主要试剂和仪器RPMI 1640培养基、DMEM培养基、Opti-MEM培养基购自美国Gibco公司;胎牛血清(FBS)、胰酶购自美国Hyclone公司;总RNA提取试剂Trizol、Lipofectamine 2000购自美国Invitrogen公司;PrimeScript RT reagent Kit、SYBR Premix Ex Taq購自日本TaKaRa公司;hsa-miR-409-3p和U6引物由上海Invitrogen公司合成;CCK-8试剂盒购自日本同仁公司;吉姆萨工作液、RIPA蛋白裂解液购自上海碧云天生物技术有限公司;细胞周期分析试剂盒购自苏州跃亚生物技术有限公司;Annexin V-EGFP凋亡检测试剂盒、BCA蛋白含量检测试剂盒购自南京凯基生物科技有限公司;蛋白Marker购自美国Thermo Scientific公司;ECL显色剂、β-actin及c-Met抗体、羊抗兔二抗购自美国SAB公司;野生型( wild type,WT)和突变型( mutant,MUT) 荧光素酶报告载体由南京科佰生物科技有限公司构建;StepOnePlus实时荧光定量PCR仪购自美国Life Technologies公司;Clinibio 128c型自动酶标分析仪购自澳大利亚ASYS公司;流式细胞仪购自美国Becton Dickinson公司;聚偏二氟乙烯膜(PVDF膜)购自美国Millpore公司;电泳仪及转膜仪购自美国Bio-Rad公司。
1.2实验方法
1.2.1数据组基因数据来源:TCGA数据集(TCGAcohort)来源于TCGA数据库(https://tcga-data.nci.Nih.gov/tega/),挑选出具有相应癌旁组织microRNA测序数据的肝癌组织microRNA测序数据进行分析,通过配对t检验对miR-409-3P在肝癌和癌旁中的表达进行差异验证。
1.2.2细胞转染及实验分组LO2、HepG2、SMMC-7721、MHCC-97H、BEL-7402经培养后用于后续试验。收集对数生长期肝癌HepG2细胞,胰蛋白酶消化,调整细胞浓度为1×105/mL,接种于96孔板(100 μL/孔),采用阳离子脂质体法分别将miR-409-3p mimics(miR-409-3p mimics组)、microRNA mimics control (miR-con组)转染至HepG2细胞,严格按照Lipofectamine 2000转染试剂说明书进行操作。
1.2.3实时荧光定量(qRT-PCR)检测细胞中miR-409-3p表达水平采用Trizol法提取细胞总RNA并测定RNA浓度,按照逆转录试剂盒将RNA反转录为cDNA。以cDNA为模板进行qRT-PCR反应。以U6为内参,检测各模板的Ct值。以Folds=2-ΔΔCt表示目的RNA在实验组与对照组中表达的相对倍比关系。实验重复3次,计算平均值。
1.2.4CCK8细胞增殖实验收集各组对数生长期HepG2细胞接种于96孔板,每孔加入2×103个细胞,调整培养液至每孔体积100 μL,每组3个复孔,同时设仅加培养基的空白对照。分别培养24、48、72、96 h后每孔加入CCK8试剂10 μL于培养箱中孵育2 h。以空白孔为对照,酶标仪设置波长450 nm测定各孔吸光度值(OD值),各组取3个复孔平均值,以相对应的平均OD值表示细胞增殖能力的大小,绘制细胞增殖曲线。
1.2.5平板克隆形成实验取对数生长期的各组转染细胞,按每孔300个细胞接种至6孔板中,每组接种3个复孔,调整培养基体积为2 mL/孔后培养2周。当6孔板中出现肉眼可见的克隆时终止培养并洗涤干燥。每孔加入4%多聚甲醛1 mL固定细胞15~30 min,弃去固定液,加入适量吉姆萨工作液染色30 min后洗去染色液,空气干燥。将6孔板倒置并叠加一张带网格的透明薄片,显微镜下计数≥5个细胞的细胞团即为一个克隆数。平板克隆形成率=克隆数/接种细胞数×100%。
1.2.6流式细胞术检测细胞周期及凋亡收集各组HepG2细胞,0.25%胰酶消化(1×106个细胞)离心,细胞沉淀转移至流式管内,按照细胞周期检测试剂盒方法进行操作,用300目尼龙筛网过滤混合液后上机检测。收集流式管中1~5×105个细胞再次离心,弃去上清,按照细胞凋亡检测试剂盒方法进行操作,轻轻混匀后上机检测。
1.2.7蛋白免疫印迹(Western Blot)检测c-Met蛋白表达取对数生长期HepG2细胞,加入适量含PMSF的裂解液,冰上裂解30 min,4 ℃条件下12 000 r/min离心30 min,吸取上清液并测蛋白浓度,取30 g蛋白样品进行SDS-PAGE电泳反应,转膜、封闭,孵育一抗(1∶1 000),4 ℃摇床内孵育过夜,TBST洗膜,加入二抗(1∶2 000),ECL显影,使用软件进行灰度扫描,以相应蛋白与β-actin的灰度比值作为蛋白的相对含量。
1.2.8荧光素酶报告基因检测miR-409-3p的靶基因
构建含有3UTR端结合位点的WT c-Met荧光素酶报告载体,同时构建突变以后不含3UTR端结合位点的MUT c-Met荧光素酶报告载体,将WT c-Met、MUT c-Met分别同miR-409-3p mimics、microRNA mimics control转染到肝癌细胞HepG2中,48 h后,用荧光素酶活性检测试剂盒测定细胞荧光素酶活性变化。
1.3统计学方法采用SPSS 20.0统计软件进行数据分析。计量资料以x±s表示,两组间比较采用成组t检验,多组间比较采用单因素方差分析,进一步两两比较采用SNK法。P<0.05为差异有统计学意义。
2结果
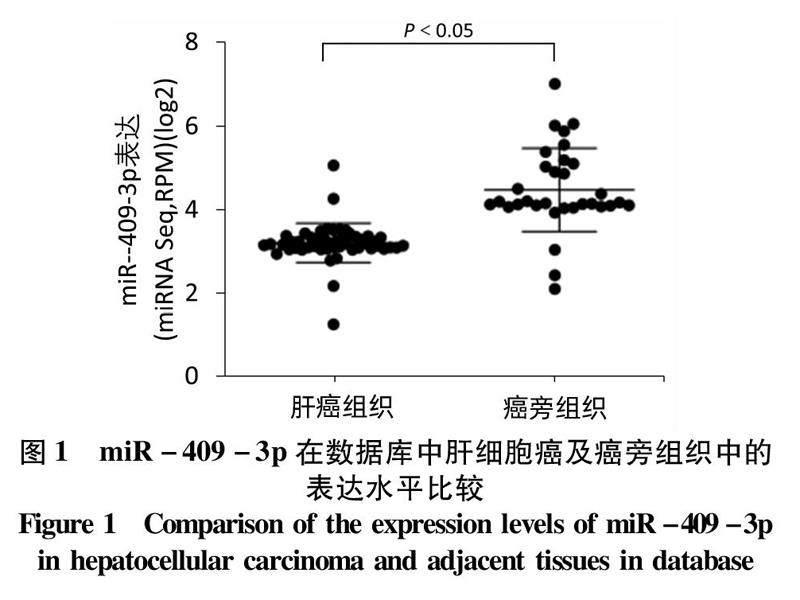
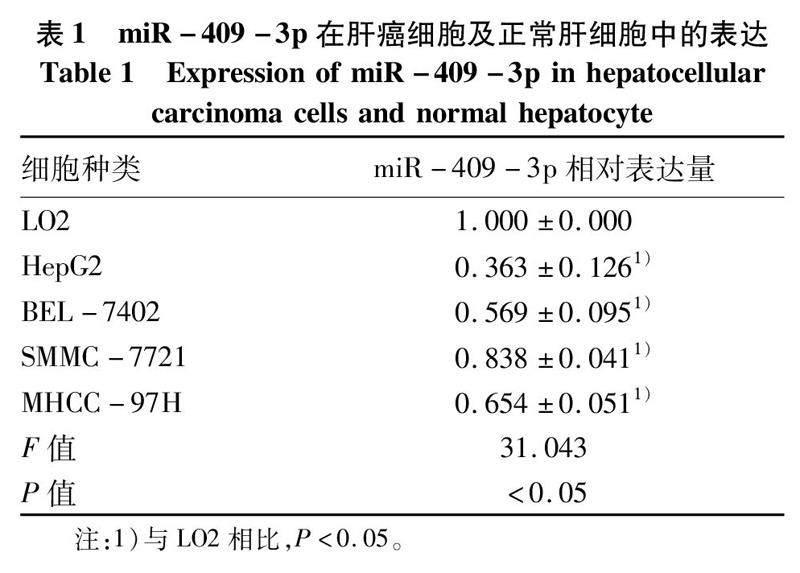
2.1miR-409-3p在肝癌中表达下调基于TCGA肝癌microRNA表达谱数据,分析肝癌及癌旁组织(53对)中miR-409-3p的表达水平发现,肝癌组织中miR-409-3p的表达显著低于其在癌旁组织中的表达(t=7.752,P<0.05)(图1)。qRT-PCR结果发现:以正常肝上皮细胞LO2为对照,miR-409-3p在4种肝癌细胞株中的表达水平均明显低于LO2(P值均<0.05)(表1)。
2.2miR-409-3p mimics转染后抑制HepG2細胞的增殖
2.2.1qRT-PCR鉴定miR-409-3p的表达采用荧光定量RT-PCR检测转染miR-409-3p mimics及miR-con两组HepG2细胞中miR-409-3p的表达水平,结果显示,miR-409-3p在mimics组中的表达水平明显高于miR-con组(8.522±1.474 vs 1.000±0.000,t=-8.836,P<0.05)。
2.2.2CCK8细胞增殖及平板克隆形成实验随着培养时间的推移,与miR-con组相比,miR-409-3p mimics组的增殖速率逐渐下降,两组在48、72及96 h比较差异均有统计学意义(P值均<0.05)(表2,图2)。此外,对两组转染细胞进行平板克隆形成实验(图3)。结果显示,连续培养14 d后,miR-con组和miR-409-3p mimics组的克隆形成率分别为0.650±0.076和0.470±0.079,miR-409-3p mimics组的细胞克隆形成率明显低于miR-con组(t=2.846,P=0.047)。
2.2.3miR-409-3p mimics转染对HepG2细胞周期的影响流式细胞仪分析结果显示,与miR-con组相比,miR-409-3p mimics组细胞G2期百分比明显增多(P<0.05)(图4,表3)。结果表明,过表达miR-409-3p可以诱导肝癌HepG2细胞G2期阻滞。
2.2.4miR-409-3p mimics转染对HepG2细胞凋亡的影响流式细胞仪分析结果显示,miR con组的凋亡率为9.47%±2.26%,miR-409-3p mimics组的凋亡率为8.30%±1.71%(图5),与miR-con组相比,miR-409-3p mimics组细胞凋亡无明显差异(t=0.714,P=0.515)。
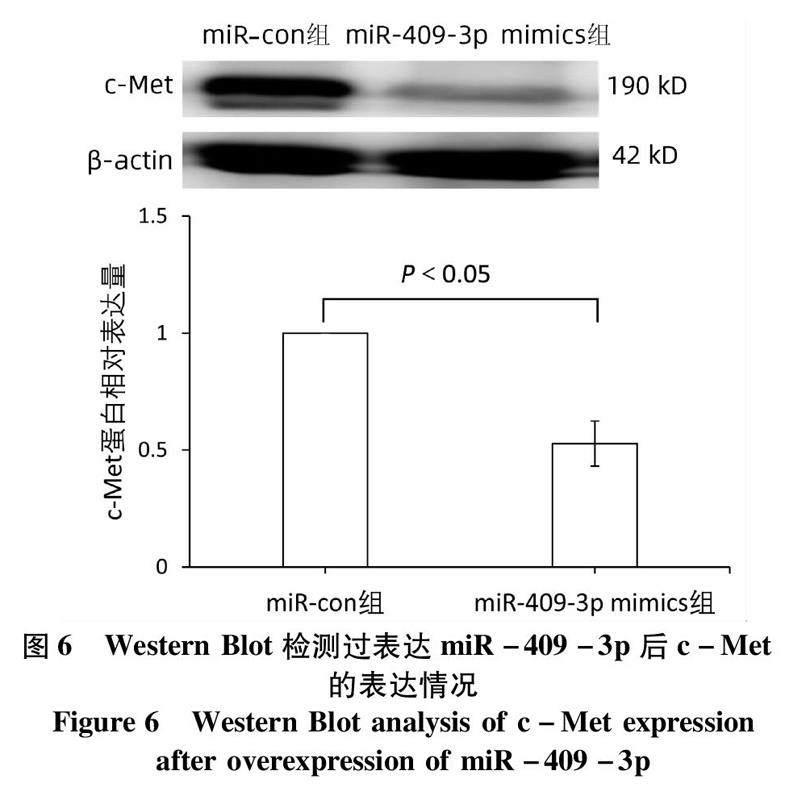
2.3miR-409-3p 抑制肝癌HepG2细胞中c-Met蛋白的表达利用生物信息学软件预测发现miR-409-3p同c-Met的3UTR端有互补结合位点,荧光素酶报告系统鉴定结果显示c-met为miR-409-3p的靶基因(表4)。与miR-con组相比,HepG2细胞转染miR-409-3p mimics后,细胞中c-Met蛋白的表达下调(t=-8.509,P=0.001)(图6)。
3讨论
HCC占原发性肝癌的75%~85%,仅美国2020年约3万人死于HCC[8-9]。全球范围内,HCC的临床转归不尽如人意,这主要归因于缺乏早期诊断的可靠指标、治疗抵抗、肿瘤的复发及转移等。目前,针对HCC高危人群所采取的每6~12个月行甲胎蛋白(AFP)联合B超检查最为常用。但是,达到HCC诊断标准的高水平血清AFP(>400 ng/mL)仅出现于少部分HCC患者。除此之外,即使是间隔3个月的B超监测仍无助于小肝癌诊断率的提高[10-11]。一定程度上,AFP与B超的联合应用不仅无益于诊断率的提高,而且会引发对假阳性的怀疑[12]。因此,迫切需要研究新的分子生物标志物来帮助临床医师早期诊断及判断预后,并依此设计新的治疗策略及方案来提高HCC的临床预后。近年来miRNA的深入研究为这一良好愿景实现的可能提供了巨大的希望[13-14]。
miRNA为非编码RNA家族成员,可调控编码蛋白质的信使RNA的稳定性及翻译过程,但其本身并不翻译成为蛋白质[15]。miRNA既可为致癌基因,亦可为抑癌基因,且与多种类型的人类癌症相关[16]。大量的证据[17]表明,miRNA的异常表达与调控细胞凋亡、增殖、细胞周期及转移的相关基因的表达有紧密的关系,可导致HCC的发生及发展。因此,有必要探索其在肿瘤发展中所扮演的重要作用。近年来,有关miR-409-3p与癌症的关系不断引起人们的关注,研究发现其在肺癌[18]、胃癌[19]、宫颈癌[20]、结肠癌[21]、乳腺癌[22]等肿瘤中发挥抑癌作用,但在HCC发生及发展中的作用需要进一步的研究加以阐明。因此,作者通过对TCGA数据库中肝癌组织和癌旁组织miR-409-3p表达水平的测序数据进行分析,并且分析了miR-409-3p在不同肝癌细胞系及正常肝细胞中的表达差异,结果显示其在肝癌中的表达水平较正常肝细胞低。功能学实验表明,转染miR-409-3p mimics可抑制肝癌HepG2细胞增殖。上述结果证明,miR-409-3p在肝癌HepG2细胞中发挥抑癌基因作用。
大量的證据证明,miRNA在功能上通过介导上述蛋白质的表达参与控制细胞周期的调控。在鼠类肝癌细胞系中,周期蛋白D2和E2已被鉴定为miR-26a的直接靶基因;而在人肝细胞癌细胞中,miR-195可调控周期蛋白D1、CDK6和E2F3的表达。因此,miR-26a或miR-195的低表达驱使G1期向S期转换[23-24]。同样地,miR-503通过下调周期蛋白D3和E2F3的表达抑制G1期向S期转换[25]。更重要的是,在HCC细胞中miR-221可负性调控周期蛋白依赖性激酶抑制因子(cyclin dependent kinase inhibitor,CDKⅠ)CDKN1B/p27和CDKN1C/p57的表达[26]。此外,miRNA可以通过抑制某些间接参与细胞周期蛋白的表达从而调控G1期向S期转换。在HCC细胞中,某些miRNA同样可以抑制G2期向M期转换,从而发挥抑癌基因作用。例如,Liu等[27]研究发现miR-517a和miR-517c的异常表达阻滞G2期向M期转换,发挥抑制HCC细胞增殖的作用。本研究发现,转染miR-409-3p mimics后肝癌细胞HepG2的G2期细胞增多,阻滞G2期向M期转换,从而抑制细胞的增殖。上述结果进一步说明,miR-409-3p通过调控细胞周期的转换影响肝癌HepG2细胞的增殖。
肝细胞生长因子受体,即c-Met,是一种致癌性的受体蛋白酪氨酸激酶,在40%~70%的HCC患者中呈高表达[28-29]。据报道,HCC中c-Met的表达可被miR-206[30]、miR-101-3p[31]和miR-128-3p[32]负性调控。在不同肿瘤中,miR-409-3p亦可通过调控c-Met的表达发挥抑癌基因作用。Xu等[6]研究表明,miR-409-3p通过靶向调控c-Met抑制膀胱癌细胞的迁移和侵袭;同样,Wan等[7]研究发现,miR-409-3p通过靶向调控c-Met发挥抑制人肺癌的作用。Huynh等[33]研究发现,采用c-Met抑制剂——Foretinib处理HCC细胞后,细胞周期蛋白B表达减少,导致G2/M期阻滞,从而抑制HCC细胞增殖及集落形成。本研究发现,转染miR-409-3p mimics后肝癌细胞HepG2中c-Met蛋白表达降低。上述结果进一步说明,miR-409-3p通过调控c-Met的表达,进一步影响下游信号通路,通过对细胞周期分布的影响抑制肝癌HepG2细胞的增殖。
然而,本实验仍有一些不足之处。首先,本实验未研究肝癌组织与正常肝组织中miR-409-3p表达差异,仅通过数据库加以分析,因此,后续实验需要收集一定数量的配对组织标本进一步明确miR-409-3p的临床意义。其次,本试验所选取的4种肝癌细胞系中,miR-409-3p的表达均显著低于正常肝细胞系,在肝癌细胞系中转染miR-409-3p 抑制剂后未出现miR-409-3p的差异性低表达,因此无法进行miR-409-3p inhibitor组的体外试验。
总之,本研究表明miR-409-3p在肝癌细胞系中呈低表达,且可以通过调控c-Met蛋白的表达抑制肝癌HepG2细胞的增殖。此miR-409-3p/c-Met轴为探索HCC的发病机制提供了一个新的见解,且为HCC的干预和治疗提供了一个更广阔的视角。但其详尽的作用机制及临床应用价值仍需要进一步更为深入的研究。
利益冲突声明:本文不存在任何利益冲突。作者贡献声明:王长青负责课题设计,实验操作,资料分析,撰写论文;陈玲、徐萍参与实验操作,收集分析数据,修改论文;朱晓娟、刘政负责拟定研究思路,指导撰写文章并最后定稿。
参考文献:
[1]RUIZ-MANRIQUEZ LM, CARRASCO-MORALES O, SANCHEZ Z EA, et al. MicroRNA-mediated regulation of key signaling pathways in hepatocellular carcinoma: A mechanistic insight[J]. Front Genet, 2022, 13: 910733. DOI: 10.3389/fgene.2022.910733.
[2]COOPER CS, PARK M, BLAIR DG, et al. Molecular cloning of a new transforming gene from a chemically transformed human cell line[J]. Nature, 1984, 311(5981): 29-33. DOI: 10.1038/311029a0.
[3]FAIELLA A, RICCARDI F, CARTEN G, et al. The emerging role of c-Met in carcinogenesis and clinical implications as a possible therapeutic target[J]. J Oncol, 2022, 2022: 5179182. DOI: 10.1155/2022/5179182.
[4]CHEN L, SHI Y, ZHU X, et al. IL-10 secreted by cancer-associated macrophages regulates proliferation and invasion in gastric cancer cells via c-Met/STAT3 signaling[J]. Oncol Rep, 2019, 42(2): 595-604. DOI: 10.3892/or.2019.7206.
[5]MA Y, ZHANG M, WANG J, et al. High-affinity human anti-c-Met IgG conjugated to oxaliplatin as targeted chemotherapy for hepatocellular carcinoma[J]. Front Oncol, 2019, 9: 717. DOI: 10.3389/fonc.2019.00717.
[6]XU X, CHEN H, LIN Y, et al. MicroRNA-409-3p inhibits migration and invasion of bladder cancer cells via targeting c-Met[J]. Mol Cells, 2013, 36(1): 62-68. DOI: 10.1007/s10059-013-0044-7.
[7]WAN L, ZHU L, XU J, et al. MicroRNA-409-3p functions as a tumor suppressor in human lung adenocarcinoma by targeting c-Met[J]. Cell Physiol Biochem, 2014, 34(4): 1273-1290. DOI: 10.1159/000366337.
[8]General Office of National Health Commission. Standard for diagnosis and treatment of primary liver cancer (2022 edition)[J]. J Clin Hepatol, 2022, 38(2): 288-303. DOI: 10.3969/j.issn.1001-5256.2022.02.009.國家卫生健康委办公厅. 原发性肝癌诊疗指南(2022年版)[J]. 临床肝胆病杂志, 2022, 38(2): 288-303. DOI: 10.3969/j.issn.1001-5256.2022.02.009.
[9]SIEGEL RL, MILLER KD, GODING SAUER A, et al. Colorectal cancer statistics, 2020[J]. CA Cancer J Clin, 2020, 70(3): 145-164. DOI: 10.3322/caac.21601.
[10]FORNER A, LLOVET JM, BRUIX J. Hepatocellular carcinoma[J]. Lancet, 2012, 379(9822): 1245-1255. DOI: 10.1016/S0140-6736(11)61347-0.
[11]RAY K. Liver cancer: The promise of new approaches in the management of hepatocellular carcinoma--adding to the toolbox?[J]. Nat Rev Gastroenterol Hepatol, 2013, 10(4): 195. DOI: 10.1038/nrgastro.2013.52.
[12]TRINCHET JC, CHAFFAUT C, BOURCIER V, et al. Ultrasonographic surveillance of hepatocellular carcinoma in cirrhosis: a randomized trial comparing 3- and 6-month periodicities[J]. Hepatology, 2011, 54(6): 1987-1997. DOI: 10.1002/hep.24545.
[13]ZHOU Q, SHAO JG. Research progress of miRNA in HBV-related hepatocellular carcinoma[J]. J Nantong Univ(Med Sci) , 2022, 42(3): 257-261. DOI: 10.16424/j.cnki.cn32- 1807/r.2022.03.013.周倩, 邵建国. MiRNA在HBV相关肝癌中的研究进展[J]. 南通大学学报(医学版), 2022, 42(3): 257-261. DOI: 10.16424/j.cnki.cn32-1807/r.2022.03.013.
[14]XIE HJ, RASHED N, NING Y, et al. Current status of research on circulating microRNAs as diagnostic markers for hepatocellular carcinoma [J]. J Clin Hepatol, 2021, 37(2): 448-451. DOI: 10.3969/j.issn.1001-5256.2021.02.042.谢惠君, Rashed Nasot, 宁勇, 等. 循环miRNA作为肝细胞癌标志物的研究现状[J]. 临床肝胆病杂志, 2021, 37(2): 448-451. DOI: 10.3969/j.issn.1001-5256.2021.02.042.
[15]HUSSEN BM, HIDAYAT HJ, SALIHI A, et al. MicroRNA: A signature for cancer progression[J]. Biomed Pharmacother, 2021, 138: 111528. DOI: 10.1016/j.biopha.2021.111528.
[16]PIEROULI K, PAPAKONSTANTINOU E, PAPAGEORGIOU L, et al. Long non-coding RNAs and microRNAs as regulators of stress in cancer (Review)[J]. Mol Med Rep, 2022, 26(6): 361. DOI: 10.3892/mmr.2022.12878.
[17]HUANG S, HE X. The role of microRNAs in liver cancer progression[J]. Br J Cancer, 2011, 104(2): 235-240. DOI: 10.1038/sj.bjc.6606010.
[18]LIU S, LI B, XU J, et al. SOD1 Promotes cell proliferation and metastasis in non-small cell lung cancer via an miR-409-3p/SOD1/SETDB1 epigenetic regulatory feedforward loop[J]. Front Cell Dev Biol, 2020, 8: 213. DOI: 10.3389/fcell.2020.00213.
[19]WANG Y, ZHANG J, CHEN X, et al. Circ_0001023 promotes proliferation and metastasis of gastric cancer cells thro ugh miR-409-3p/PHF10 axis[J]. Onco Targets Ther, 2020, 13: 4533-4544. DOI: 10.2147/OTT.S244358.
[20]CUI X, CHEN J, ZHENG Y, et al. Circ_0000745 promotes the progression of cervical cancer by regulating miR-409-3p/ATF1 axis[J]. Cancer Biother Radiopharm, 2022, 37(9): 766-778. DOI: 10.1089/cbr.2019.3392.
[21]CHEN J, WANG R, LU E, et al. LINC00630 as a miR-409-3p sponge promotes apoptosis and glycolysis of colon carcinoma cells via regulating HK2[J]. Am J Transl Res, 2022, 14(2): 863-875.
[22]YANG S, ZOU C, LI Y, et al. Knockdown circTRIM28 enhances tamoxifen sensitivity via the miR-409-3p/HMGA2 axis in breast cancer[J]. Reprod Biol Endocrinol, 2022, 20(1): 146. DOI: 10.1186/s12958-022-01011-3.
[23]KOTA J, CHIVUKULA RR, ODONNELL KA, et al. Therapeutic microRNA delivery suppresses tumorigenesis in a murine liver cancer model[J]. Cell, 2009, 137(6): 1005-1017. DOI: 10.1016/j.cell.2009.04.021.
[24]XU T, ZHU Y, XIONG Y, et al. MicroRNA-195 suppresses tumorigenicity and regulates G1/S transition of human hepatocellular carcinoma cells[J]. Hepatology, 2009, 50(1): 113-121. DOI: 10.1002/hep.22919.
[25]XIAO F, ZHANG W, CHEN L, et al. MicroRNA-503 inhibits the G1/S transition by downregulating cyclin D3 and E2F3 in hepatocellular carcinoma[J]. J Transl Med, 2013, 11: 195. DOI: 10.1186/1479-5876-11-195.
[26]FORNARI F, GRAMANTIERI L, FERRACIN M, et al. MiR-221 controls CDKN1C/p57 and CDKN1B/p27 expression in human hepatocellular carcinoma[J]. Oncogene, 2008, 27(43): 5651-5661. DOI: 10.1038/onc.2008.178.
[27]LIU RF, XU X, HUANG J, et al. Down-regulation of miR-517a and miR-517c promotes proliferation of hepatocellular carcinoma cells via targeting Pyk2[J]. Cancer Lett, 2013, 329(2): 164-173. DOI: 10.1016/j.canlet.2012.10.027.
[28]UEKI T, FUJIMOTO J, SUZUKI T, et al. Expression of hepatocyte growth factor and its receptor, the c-met proto-oncogene, in hepatocellular carcinoma[J]. Hepatology, 1997, 25(3): 619-623. DOI: 10.1002/hep.510250321.
[29]GIORDANO S, COLUMBANO A. Met as a therapeutic target in HCC: facts and hopes[J]. J Hepatol, 2014, 60(2): 442-452. DOI: 10.1016/j.jhep.2013.09.009.
[30]WANG Y, TAI Q, ZHANG J, et al. MiRNA-206 inhibits hepatocellular carcinoma cell proliferation and migration but promotes apoptosis by modulating cMET expression[J]. Acta Biochim Biophys Sin (Shanghai), 2019, 51(3): 243-253. DOI: 10.1093/abbs/gmy119.
[31]LIU Y, TAN J, OU S, et al. MicroRNA-101-3p suppresses proliferation and migration in hepatocellular carcinoma by targeting the HGF/c-Met pathway[J]. Invest New Drugs, 2020, 38(1): 60-69. DOI: 10.1007/s10637-019-00766-8.
[32]XU X, JIANG W, HAN P, et al. MicroRNA-128-3p Mediates Lenvatinib Resistance of Hepatocellular Carcinoma Cells by Downregulating c-Met[J]. J Hepatocell Carcinoma, 2022, 9: 113-126. DOI: 10.2147/JHC.S349369.
[33]HUYNH H, ONG R, SOO KC. Foretinib demonstrates anti-tumor activity and improves overall survival in preclinical models of hepatocellular carcinoma[J]. Angiogenesis, 2012, 15(1): 59-70. DOI: 10.1007/s10456-011-9243-z.
收稿日期:2022-11-05;錄用日期:2023-02-27
本文编辑:王莹
引证本文:WANG CQ, CHEN L, XU P, et al. Expression of miR-409-3p in hepatoma HepG2 cells and its mechanism in cell proliferation[J]. J Clin Hepatol, 2023, 39(8): 1895-1902.