户太8号葡萄抗寒性初步研究
2023-03-24宋光永厉晓婷栗晓妍党晓文郑旭成周龙
宋光永,厉晓婷,栗晓妍,党晓文,郑旭成,周龙*
(新疆农业大学园艺学院,新疆乌鲁木齐 830052)
新疆是典型的温带大陆性气候,非常适合栽培葡萄[1],栽培历史悠久,2020年产量达305万t,占全国的21.35%[2],是中国最大的葡萄产区。但冬季严寒,葡萄易遭冻害,轻则枝条抽干,重时导致植株死亡[3]。研究葡萄的抗寒性对新疆乃至北方寒冷地区葡萄正常生长结果具有重要意义。
户太8号葡萄是中国自主选育、具有自主知识产权的优良品种[4,5],耐贮运、不裂果、连续结果能力强,有玫瑰香味,比中熟品种巨峰早上市15 d左右。
当前对户太8号葡萄的研究集中在栽培管理[4,5]、果实品质[6]、酿酒[7]等方面,对其抗寒性的研究较少。笔者对户太8号葡萄一年生休眠枝条进行不同的低温处理,观察测定在恢复生长期的形态表现和生理指标表现,分析评判其抗寒性,为户太8号葡萄在新疆地区的栽培推广提供理论参考。
1 试验材料
户太8号一年生休眠枝条采自于新疆生产建设兵团第十二师头屯河农场。植株5年生,树势中庸,篱架栽培,株行距1 m×4 m,常规水肥管理,冬季覆土越冬。
2021年9月下旬植株开始落叶进入休眠期,11月20日采集长势良好一致、无病虫危害的一年生枝条80条,长度15~20 cm,用棉被包裹埋于雪中贮藏。雪中贮藏7 d至11月27日取出,至此,户太8号枝条已基本满足需冷量。枝条用自来水清洗干净,再用蒸馏水冲洗3次,吸水纸吸干水分放置在不同低温条件下处理。
1.1 试验设计与方法
2021年11月27日对枝条进行低温处理,设8个低温梯度:-4 ℃(对照)、-7 ℃、-10 ℃、-13 ℃、-16 ℃、-19 ℃、-23 ℃、-27 ℃,(一般来说,葡萄能抗-6~-8 ℃的低温,参照前人对于植物人工模拟低温胁迫的试验,采用-4 ℃作为对照)。以降温速率4 ℃/h分别降至目的温度,持续10 h,再以升温速率4 ℃/h都升温至25 ℃,解除低温胁迫进入生长条件。枝条自低温处理到恢复生长试验完成(2021年11月27日至2022年1月5日)共计39 d。参照前人对枝条恢复生长的研究方法,每隔3 d观察拍照1次,共10次。
枝条恢复生长条件1个月内可萌芽,所受低温伤害越轻,萌芽越早。30 d后仍不萌芽的枝条,解剖观察内部已干枯。
1.2 指标测定
萌芽率 将不同低温处理后统一升温至25 ℃的枝条,在自来水中浸泡12 h,剪成上端为平口、下端为斜口的单芽茎段,蜡封上端切口,插入三角瓶中。在室温25 ℃条件下进行水培,每个处理重复3次。每3 d观察1次萌芽情况并拍照,观察生长情况,计算萌芽率。

枝蔓电解质渗出率 将低温处理后恢复室温的枝蔓,用蒸馏水冲洗干净,切成薄片,混均取样0.1 g放入25 mL玻璃试管中,加20 mL蒸馏水,在25 ℃下,用频率150 r/min的摇床振荡4 h。用雷磁DDS-307A电导率仪测定其初电导率(E1)、蒸馏水对照电导率(E0),沸水浴30 min,冷却至室温后,再测定其终电导率(E2),每个处理重复3次,取平均值。计算电解质渗出率。
将求得的电解质渗出率(y),和处理温度(t)用Logistic方程进行拟合。根据试验得出的电解质渗出率与处理温度画出散点图,采用目前研究中常用的Logistic方程即拟合度最高的曲线,可在SPSS、Origin、R等常用软件拟合。利用电解质渗出率求半致死温度的公式,计算出拐点温度即为低温半致死温度(LT50)。
式中y为上一个公式所求出的电解质渗出率;a、b为Logistic方程中的参数;t为处理温度;k为枝条细胞受到低温伤害的饱和容量。
组织褐变率 组织褐变率为枝条横切面发生褐变面积(S1)与整个横切面面积(S2)之比。参照牛立新的方法[8],将低温处理恢复室温的枝条,用蒸馏水冲洗干净,切成薄片,放置在舜宇SZ45解剖镜下,观察枝条横切面的受冻害褐变程度及木质部受冻害产生的变化。每个处理重复3次,取平均值。
丙二醛(MDA)、脯氨酸(Pro)的含量 采用武汉伊莱瑞特生物科技公司试剂盒测定。
1.3 数据处理
使用Microsoft Office Home and Student 2019进行数据计算,采用OriginPro 2022 SR1(学习版)分析数据。
2 结果与分析
2.1 枝蔓恢复生长情况
冬芽的萌动程度,参照李华编著的《葡萄栽培学》相关指标进行描述[9]。从图1可看出,随着处理温度的降低,户太8号葡萄枝条恢复生长的能力渐弱。-4~-7 ℃处理后的冬芽在30 d内进入展叶期,全部萌发;-13 ℃处理的冬芽在30 d内只进入绿尖期,个别芽未萌发;当-16 ℃或更低温处理时,冬芽一直未萌动。

图 1 不同低温处理的户太8号葡萄一年生枝条在30 d的生长情况
2.2 枝条组织的生理指标
葡萄枝条组织褐变情况。表1显示,随着处理户太8号枝条的低温的降低,枝条组织褐变程度渐重。-4~-7 ℃处理的枝条无组织褐变发生;-10 ℃处理的枝条有部分褐变发生,褐变率为35%;-13 ℃处理的枝条褐变率超过50%;-16 ℃处理的枝条肉眼可见褐变,褐变率达90%;-19 ℃处理的枝条褐变率达100%。
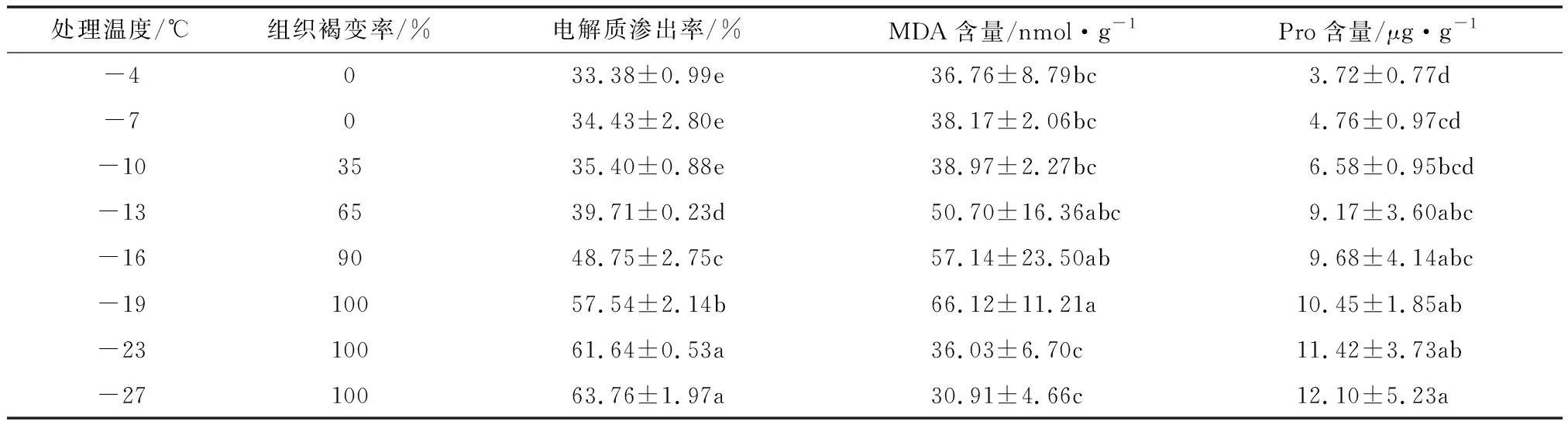
表1 户太8号一年生枝条不同低温处理的生理指标变化
枝条电解质渗出率的变化。随着枝条低温处理的下降,户太8号枝条电解质渗出率逐渐上升,大体上呈现“慢—快—慢”的增长趋势,-4~-10 ℃处理时,电解质渗出率缓慢上升;-10~-19 ℃处理时,电解质渗出率上升加快;-19 ℃处理时,电解质渗出率超过50%;-27 ℃时处理时,电解质渗出率达到63.76%。
枝条中MDA含量的变化。随着枝条处理低温的降低,户太8号枝条中MDA含量呈先升后降的变化趋势。-4~-10 ℃处理时,MDA含量从36.76 nmol/g开始缓慢上升;-10~-19 ℃处理时,MDA含量上升趋势变快,达到峰值66.12 nmol/g;-23~-27 ℃处理时则急剧下降至30.91 nmol/g。
枝条中Pro含量的变化。随着枝条处理低温的降低,户太8号枝条中Pro含量呈上升趋势。-4~-7 ℃处理时,Pro含量从3.72 μg/g缓慢上升;-7~-13 ℃处理时,Pro含量上升趋势加快;-13~-27 ℃处理时,Pro含量上升趋势变慢,最终达到峰值12.10 μg/g。
2.3 低温半致死温度
随着枝条处理低温的下降,户太8号枝条电解质渗出率逐渐增加,温度与电解质渗出率之间的关系大致呈S型曲线,基本符合Logistic方程。将不同低温处理的枝条所测得的电解质渗出率和Logistic方程拟合,求得低温半致死温度LT50(表2),拟合度为0.94417、参数 a为3.02651、b为0.06544、k为100,拟合度较好,结果可信,户太8号枝条的半致死温度为-16.92 ℃。

表2 户太8号葡萄枝条的低温半致死温度
3 小结与讨论
在模拟梯度低温胁迫下,户太8号葡萄枝条的恢复生长能力越来越弱,直到无法恢复;组织褐变面积逐渐变大,直到全部褐变;电解质渗出率逐渐升高,用Logistic方程拟合,求得低温半致死温度为-16.92 ℃;丙二醛含量先升后降,游离脯氨酸含量逐渐升高。
植物遇到低温伤害,具有一定的自我恢复能力,研究其遭遇低温伤害后的恢复生长能力,可用来评价其抗寒性的强弱[10]。植物遭遇低温伤害后是否存活,主要通过芽对低温的敏感性反映,芽在低温下能否存活是鉴定植物抗寒性最直接和有效的方法之一[11]。本研究发现,户太8号一年生枝条经-13 ℃低温处理,30 d内冬芽只能进入绿尖期,个别冬芽无法萌动。-16 ℃或更低温处理,冬芽一直未萌动。因为当植物受到冻害程度严重时,会影响其恢复生长,甚至恢复生长受阻[12]。户太8号葡萄在低温处理时,造成冬芽内细胞间和细胞内水分先后结冰[13,14],细胞亚显微结构破坏、细胞生理代谢失调;另一方面在温度恢复过程中,造成细胞变形、细胞内原生质体破裂,细胞受到机械应力损伤。两方面综合作用,使得冬芽内部细胞部分死亡,幸存的细胞勉强支撑冬芽萌动。
在外界低温下,细胞膜中积累的自由基诱导膜脂中不饱和脂肪酸发生脂质过氧化,膜脂过氧化最终产物是MDA[15]。植物体内MDA含量可以反映其面对低温时的细胞膜受损程度以及膜脂过氧化水平[16]。本研究发现,当进行-4~-27 ℃梯度低温处理时,MDA含量出现先升后降,-16~-19 ℃时出现峰值。这与向导等[17]、贾金辉等[18]研究葡萄抗寒性的结果相似。分析认为:在低温处理下,植物细胞内清除和产生的自由基动态平衡遭到破坏,自由基逐渐积累,诱导脂质过氧化作用逐渐增强MDA的过量积累,一方面与枝条内的蛋白质、核酸等大分子反应,使其丧失生理功能,另一方面使得枝条内纤维素分子间的桥键松驰[19]。两方面综合作用,使得枝条内部部分细胞受损,甚至死亡,丧失了生理功能。
本研究为户太8号葡萄的抗寒性初步研究,未涉及与其他品种的对比,只以-4 ℃为对照,初步评判其抗寒性,为后期进行深入研究奠定了基础。
