黄河三角洲滨海滩涂不同密度柽柳林的土壤盐碱与养分特征
2023-01-13夏江宝王善龙高芳磊赵万里马金昭
陈 萍,夏江宝,王善龙,李 栋,高芳磊,赵万里,马金昭
1 山东农业大学林学院,泰山森林生态系统国家定位观测研究站, 泰安 271018 2 滨州学院,山东省黄河三角洲生态环境重点实验室, 滨州 256603 3 东平县林业保护发展中心, 泰安 271504 4 滨州市城乡水务发展服务中心, 滨州 256603
林分密度是影响林木生长的重要因子,不仅对环境资源分配和种内竞争起到非常重要的作用,而且对土壤理化性质和养分循环等有显著影响[1—2]。密度和植物-土壤反馈有关,植物对密度具有依赖性[3—4],Carson等[4]发现125种植物中有40%表现出密度依赖性死亡。适宜的林分密度可以改善土壤结构,提高生物产量。研究表明,莱州湾南岸滩涂中密度(3600 株/hm2)柽柳林改良土壤、修复植被的效果相对最佳[5]。柽柳生长能对土壤状况产生影响,而改善的立地土壤环境又有利于柽柳生长、更新和扩散,从而土壤与柽柳之间形成了正反馈作用机制,易出现“肥岛”效应[6]。“肥岛”是指土壤养分在生物与非生物过程作用下富集于灌木周围,表现为土壤养分含量由冠内向冠外逐渐递减的现象。“肥岛”效应和植被的生长发展相互促进,对改善柽柳灌丛下的土壤肥力以及植物的更新和扩散具有重要意义[7—8]。土壤盐碱状况是衡量盐碱土理化性质的基础,也是土壤肥力的重要影响因素之一[9]。因黄河三角洲泥质海岸带地下水位浅且矿化度高,降雨量少、蒸发量大和海水入侵等因素影响,导致滨海滩涂地带土壤盐渍化严重,植物不易在中重度盐碱区生存,利用耐盐植物进行生态修复可以有效改善滨海盐渍土壤的结构,抑制土壤盐碱化;另一方面生态环境的改善能为其它植物的定居和生长繁衍提供必要条件,对盐碱地的改良与植被恢复具有重要意义[10—11]。柽柳是泌盐植物,具有富集土壤盐分、改良土壤等作用,柽柳能通过吸收土壤盐分调节自身渗透压,从而适应高盐度环境下的生长生存。种植柽柳后冠下土壤的含盐量低于周围土壤的现象称为“盐谷”效应。“盐谷”效应的出现表明柽柳林对土壤盐分有迁移、疏导作用。但也有研究发现,柽柳林并未对林下草本群落表现出明显的“肥岛”、“盐谷”效应[12]。位于黄河三角洲区域的山东省滨州市泥质海岸带滨州港林场分布着大面积柽柳天然群落,对改善区域生态环境状况和维护海岸带生态系统稳定发挥着重要作用。目前对黄河三角洲柽柳的研究主要集中在滨海湿地柽柳种群空间分布格局[13—15]、柽柳群落的土壤环境特征[16—18]、柽柳林生长动态对密度结构的响应特征[10]、不同密度柽柳林地土壤调蓄水功能[19]以及柽柳林湿地肥岛效应研究[6]等方面。但关于滨海滩涂不同柽柳林分密度条件下是否存在“盐谷”、“肥岛”效应以及柽柳林土壤养分、盐分富集作用强度的差异尚不清楚,导致该区域柽柳林分密度调控以及柽柳林生态保护和抚育提升管理存在一定困难。因此,探明黄河三角洲滨海滩涂不同密度柽柳林的土壤盐碱和养分特征,明确何种密度下土壤降盐抑碱和改善土壤养分效果好,对柽柳林分密度调整、管护和土壤改良等都具有一定指导意义。
本研究以3种不同林分密度的柽柳林为研究对象,测定分析不同密度柽柳林及柽柳植株周边不同位置的土壤盐碱状况、养分特征等指标,探讨不同密度柽柳林是否具有明显的“盐谷”、“肥岛”效应,明确不同密度柽柳林对土壤的改良作用,研究结果可为泥质海岸带滨海滩涂柽柳灌丛的生态保护以及低效柽柳林的密度调控管理提供科学依据。
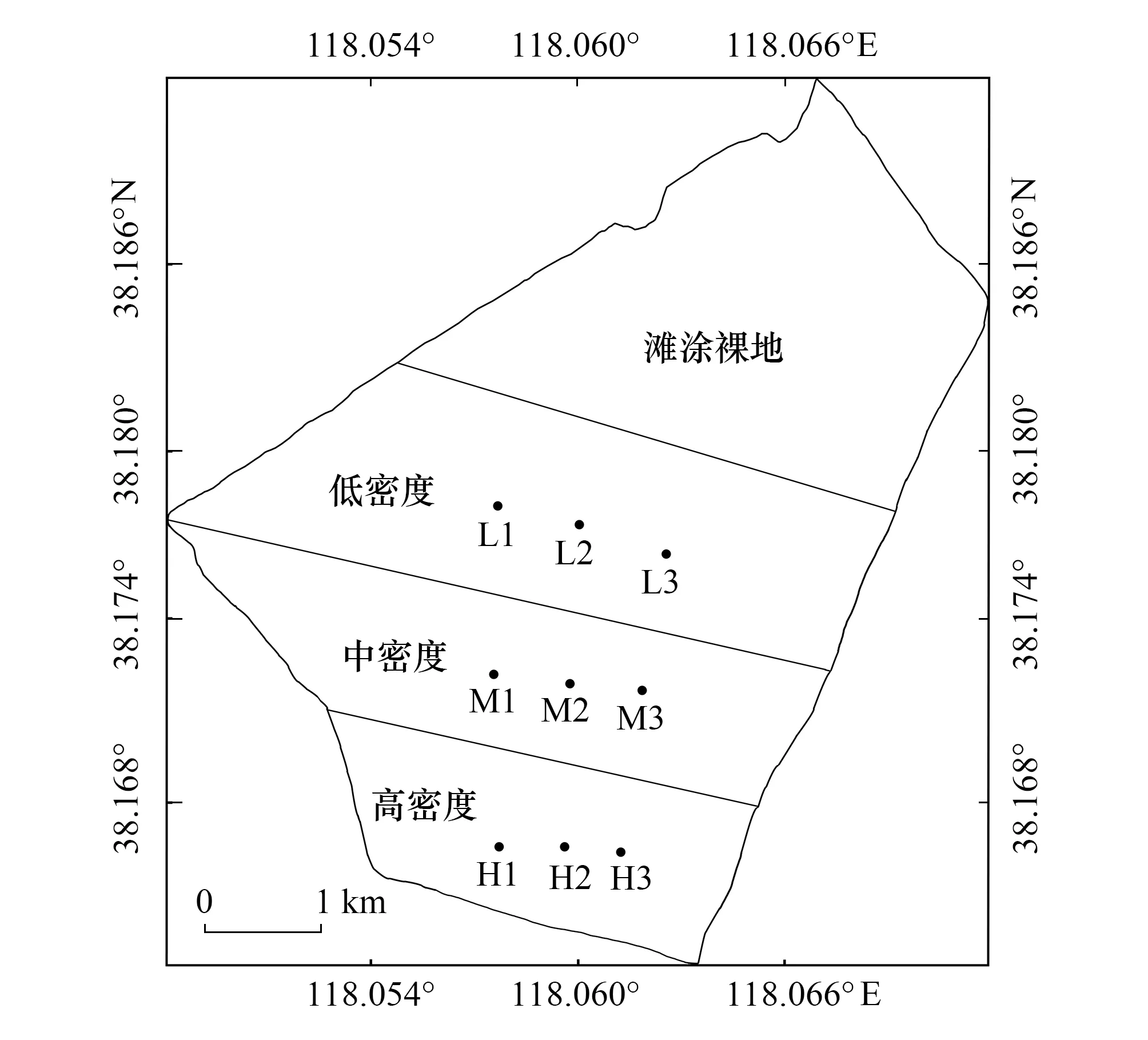
图1 不同密度柽柳林采样点位图 Fig.1 Schematic map of sampling sites of Tamarix chinensis under different density shrubsL: 低密度柽柳林采样点Sampling sites of low-density T. chinensis; M: 中密度柽柳林采样点 Sampling sites of medium-density T. chinensis; H: 高密度柽柳林采样点 Sampling sites of high-density T. chinensis
1 材料与方法
1.1 研究区概况
研究地点位于渤海湾西南岸的山东省滨州市北海新区黄河三角洲东南部的滨州港林场(38°9′51″—38°10′44″N,118°2′15″—118°4′32″E),研究区总面积达5.58 km2。该区域为暖温带半湿润地区,大陆性季风气候,年平均气温为12.6 ℃,年平均无霜期为203 d,年平均日照时间为2850 h,年平均降水量543.2 mm,年平均蒸发量1806 mm,降水集中在6—9月,占年降水量的75%。研究区地下水位浅、矿化度高;地貌属于滨海缓平低地,地形低洼平缓,海拔为0.5—1.0 m,坡度为0.1‰—0.2‰;滩涂土壤为冲积性黄土母质,以粉砂和淤泥质粉砂为主,沙粘相间,易于压实,通气透水性差;滩涂主要分布有柽柳和芦苇(Phragmitesaustralis)、盐地碱蓬(Suaedasalsa)、白茅(Imperatacylindrica)、狗尾草(Setariaviridis)、鹅绒藤(Cynanchumchinense)等植物。该柽柳群落是自然演替形成,主要由盐地碱蓬逐步过渡到柽柳群落,盐地碱蓬的生长起到了降盐改土作用,增加了土壤有机质含量,为柽柳群落生长创造了条件。该区域地貌类型为滨海滩涂地带,土壤来源于黄河沉积物携大量泥沙淤海造陆形成的,即为冲积性黄土母质在海浸母质上沉淀而成。
1.2 样品采集与处理
野外采样时间为2019年8月,在山东省滨州市北海新区的滨州港林场,选取林龄为5 a的柽柳林地作为采样地,根据前期课题组对该区域的调查,针对柽柳生长密度情况,以低密度(1100 株/hm2)、中密度(4100 株/hm2)、高密度(7100 株/hm2)三种林分密度的柽柳林为研究对象,采样点布设见图1,其中低、中、高密度柽柳林面积分别为1.25 km2、1.60 km2、1.17 km2。每种密度柽柳林各设置3个20 m×20 m的标准地(低密度柽柳林采样点分别为L1、L2、L3;中密度柽柳林采样点分别为M1、M2、M3;高密度柽柳林采样点分别为H1、H2、H3),共9个样地,调查样地的生境条件基本一致(坡向、坡度、海拔等),在每块样地内沿对角线设置3个 5 m×5 m的灌木小样方,分别记录样方内每株柽柳株高、冠幅、盖度等指标,并根据株高、冠幅等指标在样方内选择长势均匀、生长良好的3 株柽柳标准木,即每个密度下选择9 株标准木,不同密度柽柳的形态特征见表1。
在每个样方中,以柽柳灌丛为圆心,在柽柳主根部位、冠幅中心(根部与灌丛边缘之间的中心)、冠幅边缘(灌丛冠幅在地表垂直投影的位置)和株间空地(灌丛之间的裸地)共4个不同位置(图2),分别取不同方位(东、西、南、北) 0—20 cm土壤深度的4个土样,每4个不同方位的土样均匀混合后作为一个样株位置的土壤样品,装入自封袋带回实验室,经自然风干后去除植物残体和石块,测定其土壤盐碱及养分指标。柽柳周边各位置9 (3×3) 个土样,共36 (4×3×3) 个土样。

图2 柽柳土壤采样示意图 Fig.2 Schematic diagram of Tamarix chinensis soil samplingA: 柽柳根部 Root of T. chinensis; B: 冠幅中心 Canopy center; C: 冠幅边缘 Canopy brim; D: 株间空地 Interspaces
1.3 土壤样品的测定
采用电位法测定土壤pH(水土比5∶1浸提);采用电导率仪(HANNA, HI8733)测定土壤浸提液(水土比5∶1)电导率;采用碱解扩散法测定土壤速效氮含量;采用钼锑抗比色法测定土壤样品的速效磷含量;采用醋酸铵-火焰光度计法测定土壤速效钾含量;采用TOC分析仪(VSeriesSSM- 5000A)测定土壤有机碳含量。
1.4 数据处理
采用Excel 2016和SPSS 22.0软件对土壤盐碱及养分指标等进行统计分析,采用Origin 2018软件作图。运用单因素(one-way ANOVA)和Duncan法进行方差分析 (α=0.05),比较柽柳根部、冠幅中心、冠幅边缘、株间空地之间土壤盐碱和养分含量差异,以及不同密度柽柳土壤盐碱和养分含量及其富集率的差异。图表中数据为9次重复的平均值±标准差,各指标相关性采用Pearson相关分析。
富集率可以用来表示土壤养分、盐分富集程度[20]。EA=A/D,表示柽柳根部养分、盐分富集状况;EB=B/D,表示柽柳冠幅中心养分、盐分富集状况;EC=C/D,反映柽柳冠幅边缘土壤养分、盐分富集状况。其中A、B、C、D分别表示根部、冠幅中心、冠幅边缘、株间空地的土壤养分或盐分含量。
2 结果与分析
2.1 不同密度柽柳林及其柽柳植株周边不同位置的土壤盐碱含量
由图3可知,相同密度柽柳林植株周边不同位置的土壤电导率差异显著(P<0.05)。低密度下,柽柳林根部土壤电导率显著高于冠幅中心、冠幅边缘和株间空地,与根部相比,3个位置分别降低54.30%、33.82%、22.71%;中密度下,柽柳林土壤电导率表现为根部<冠幅中心<冠幅边缘<株间空地,与株间空地相比,分别降低63.02%、52.10%、34.26%;高密度下,柽柳林根部土壤电导率与冠幅中心、冠幅边缘相比差异不显著(P>0.05)。中、高密度柽柳林的土壤电导率整体表现出“根部和冠幅中心小、冠幅边缘大、株间空地更大”的特点。
植株周边相同位置不同密度柽柳林的土壤电导率差异显著(P<0.05)。从根部来看,中、高密度柽柳林土壤电导率显著低于低密度,与低密度相比,根部土壤电导率分别降低69.45%、75.04%;从冠幅中心来看,低、中密度柽柳林土壤电导率差异不显著(P>0.05),但显著高于高密度;从冠幅边缘来看,低、中密度柽柳林土壤电导率显著高于高密度,与高密度相比,分别增加119.82%、80.28%;从株间空地来看,低密度柽柳林土壤电导率和中密度相比差异不显著(P>0.05),电导率均值大小表现为中密度>低密度>高密度。不同密度柽柳林地土壤电导率均值表现为低密度>中密度>高密度,中、高密度柽柳林土壤电导率分别比低密度降低28.39%、55.74%。
由图3可知,相同密度柽柳林植株周边不同位置的土壤pH差异不显著(P>0.05),高密度柽柳林土壤pH出现根部和冠幅边缘的土壤pH高于株间空地的现象,而低、中密度柽柳林的土壤pH表现出根部和冠幅边缘低于株间空地。
植株周边相同位置不同密度柽柳林土壤pH差异也不显著(P>0.05),不同密度柽柳林土壤pH均值表现为中密度>高密度>低密度。
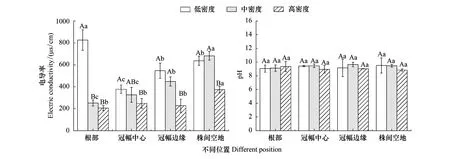
图3 不同密度柽柳林及其柽柳植株周边不同位置的土壤电导率和pH含量Fig.3 Soil electrical conductivity and pH content in different densities of Tamarix chinensis and different positions不同大写字母者表示不同密度柽柳林植株周边相同位置差异显著(P<0.05); 不同小写字母表示相同密度柽柳林植株周边不同位置差异显著(P<0.05)
2.2 不同密度柽柳林的土壤养分含量
由图4可知,植株周边相同位置不同密度柽柳林土壤速效氮含量差异显著(P<0.05)。从根部来看,中、高密度柽柳林土壤速效氮含量显著高于低密度,与低密度相比,分别增加88.59%、65.04%;从冠幅中心来看,不同密度柽柳林土壤速效氮含量差异显著(P<0.05),表现为中密度>高密度>低密度,与低密度相比,中、高密度分别增加109.59%、46.36%;从冠幅边缘来看,中、高密度柽柳林土壤速效氮含量差异不显著(P>0.05),但显著高于低密度;从株间空地来看,低、中密度柽柳林土壤速效氮含量显著高于高密度,与高密度相比,分别增加42.04%、18.03%。不同密度柽柳林地土壤速效氮含量均值表现为中密度>高密度>低密度,中、高密度柽柳林土壤速效氮含量分别比低密度增加49.80%、25.83%。
由图4可知,植株周边相同位置不同密度柽柳林土壤速效磷含量差异显著(P<0.05),且均表现为中密度>高密度>低密度。不同密度柽柳林土壤速效磷含量均值表现为中密度>高密度>低密度,中、高密度柽柳林土壤速效磷含量分别比低密度增加120.05%、58.48%。
由图4可知,植株周边相同位置不同密度柽柳林土壤速效钾含量差异显著(P<0.05)。从根部来看,中、高密度柽柳林土壤速效钾含量显著高于低密度,与低密度相比,分别增加23.49%、40.44%;从冠幅中心来看,中、高密度柽柳林土壤速效钾含量差异不显著(P>0.05),但均显著高于低密度;从冠幅边缘来看,不同密度柽柳林土壤速效钾含量差异显著(P<0.05),表现为高密度>中密度>低密度;从株间空地来看,中、高密度柽柳林土壤速效钾含量显著高于低密度,分别比低密度增加89.31%、67.77%。不同密度柽柳林土壤速效钾含量均值表现为中密度>高密度>低密度,与低密度相比,中、高密度分别增加58.90%、65.34%。
由图4可知,从根部来看,不同密度柽柳林土壤有机碳含量差异显著(P<0.05),表现为中密度>高密度>低密度;从冠幅中心来看,中、高密度柽柳林土壤有机碳含量高于低密度,与低密度相比,分别增加38.96%、16.78%;从冠幅边缘和株间空地来看,不同密度柽柳林土壤有机碳含量差异不显著(P>0.05)。不同密度柽柳林土壤有机碳含量表现为中密度>高密度>低密度,与低密度相比,分别增加65.05%、22.80%。
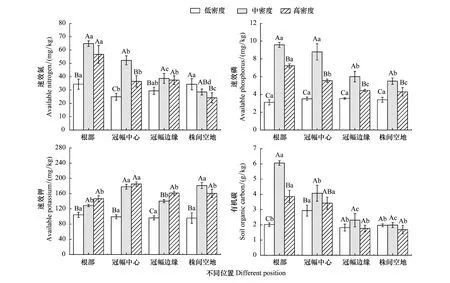
图4 不同密度柽柳林及其柽柳植株周边不同位置的土壤养分含量Fig.4 Soil nutrient content in different densities of Tamarix chinensis and different positions
2.3 不同密度柽柳林植株周边不同位置的土壤养分差异
由图4可知,低密度下,柽柳林根部速效氮、有机碳含量和冠幅边缘、株间空地差异不显著(P>0.05),但均高于冠幅中心;柽柳林根部速效磷、速效钾含量与冠幅中心、冠幅边缘、株间空地均差异不显著(P>0.05)。
中密度下,柽柳植株周边不同位置的土壤速效氮含量差异显著(P<0.05),表现为根部>冠幅中心>冠幅边缘>株间空地,与株间空地相比,3个位置分别增加127.36%、83.22%、35.64%;柽柳林根部速效磷含量显著高于冠幅边缘和株间空地,与冠幅中心差异不显著(P>0.05),表现为根部>冠幅中心>冠幅边缘>株间空地;柽柳林根部速效钾含量显著低于冠幅中心和株间空地,与冠幅边缘相比差异不显著(P>0.05);不同位置柽柳林有机碳含量差异显著(P<0.05),表现为根部>冠幅中心>冠幅边缘>株间空地。
高密度下,柽柳植株周边不同位置的土壤速效氮含量差异显著(P<0.05),表现为根部>冠幅中心>冠幅边缘>株间空地,与株间空地相比,3个位置分别增加134.78%、54.79%、50.97%;柽柳林根部速效磷含量表现为根部>冠幅中心>株间空地>冠幅边缘;柽柳林根部速效钾含量与冠幅边缘、株间空地差异不显著(P>0.05);不同位置柽柳林有机碳含量差异显著(P<0.05),表现为根部>冠幅中心>冠幅边缘>株间空地。
2.4 不同密度柽柳林及其柽柳植株周边不同位置的土壤盐碱及养分的富集率差异
由表2可知,从根部来看,低、中密度柽柳林土壤pH富集率显著低于高密度,与高密度相比,分别降低8.65%、7.69%;不同密度柽柳林土壤电导率富集率差异显著(P<0.05),表现为低密度>高密度>中密度;中、高密度柽柳林土壤速效氮富集率显著高于低密度,分别比低密度增加1.27倍、1.35倍;中、高密度柽柳林土壤速效磷富集率显著高于低密度,与低密度相比,分别增加0.82倍、1.06倍;不同密度柽柳林土壤速效钾富集率差异显著(P<0.05),表现为低密度>高密度>中密度;不同密度柽柳林有机碳富集率差异显著(P<0.05),表现为中密度>高密度>低密度。
从冠幅中心来看,土壤pH富集率差异不显著(P>0.05);低、高密度柽柳林土壤电导率富集率显著高于中密度,与中密度相比,分别增加31.11%、35.56%;土壤速效氮富集率差异显著(P<0.05),具体表现为中密度>高密度>低密度;中密度柽柳林土壤速效磷富集率显著高于低密度,但与高密度差异不显著(P>0.05);低、中密度柽柳林土壤速效钾富集率显著低于高密度,与高密度相比,分别降低9.56%、14.78%;中、高密度柽柳林有机碳富集率显著高于低密度,表现为中密度>高密度>低密度。
从冠幅边缘来看,中密度柽柳林土壤pH富集率显著高于低密度,但与高密度相比差异不显著(P>0.05);中、高密度柽柳林电导率富集率显著低于低密度,与低密度相比,分别降低23.25%、20.93%;中、高密度柽柳林土壤速效氮富集率显著高于低密度,表现为高密度>中密度>低密度;土壤速效磷富集率差异不显著(P>0.05);低、高密度柽柳林土壤速效钾含量显著高于中密度,与中密度相比,分别增加29.87%、31.17%;中、高密度柽柳林有机碳富集率显著高于低密度,表现为中密度>高密度>低密度。

表2 不同密度柽柳林及其柽柳植株周边不同位置的土壤盐碱和养分富集率
2.5 不同密度柽柳林植株周边不同位置土壤盐碱含量与养分的相关性分析
由表3可知,在柽柳根部,林分密度和电导率呈极显著负相关(P<0.01),与速效钾呈极显著正相关(P<0.01),与速效氮呈显著正相关(P<0.05),与速效磷、有机碳相关性不显著。电导率和速效氮、速效磷、速效钾、有机碳呈极显著负相关(P<0.01),表明土壤盐分与养分关系密切。速效氮和速效磷、有机碳呈极显著正相关(P<0.01),速效磷和有机碳呈极显著正相关(P<0.01)。
在冠幅中心,林分密度与电导率呈极显著负相关(P<0.01),与速效钾呈极显著正相关(P<0.01),与其他指标相关性不显著。电导率和速效钾呈显著负相关(P<0.05)。速效氮和速效磷、有机碳呈极显著正相关(P<0.01),速效磷和速效钾、有机碳呈显著正相关(P<0.05)。
在冠幅边缘,林分密度和电导率呈极显著负相关(P<0.01),与速效钾含量呈极显著正相关(P<0.01),与其他指标相关性不显著。速效氮和速效磷、速效钾呈显著正相关(P<0.05)。
在株间空地,林分密度和电导率、速效氮呈显著负相关(P<0.05),与速效钾呈显著正相关(P<0.05),速效磷和速效钾呈显著正相关(P<0.05)。
在柽柳根部、冠幅中心、冠幅边缘和株间空地,各土壤因子具有一定的相关性,但林分密度与速效磷、有机碳无显著相关性,与电导率呈极显著性负相关(P<0.01),与速效钾呈极显著正相关(P<0.01)。
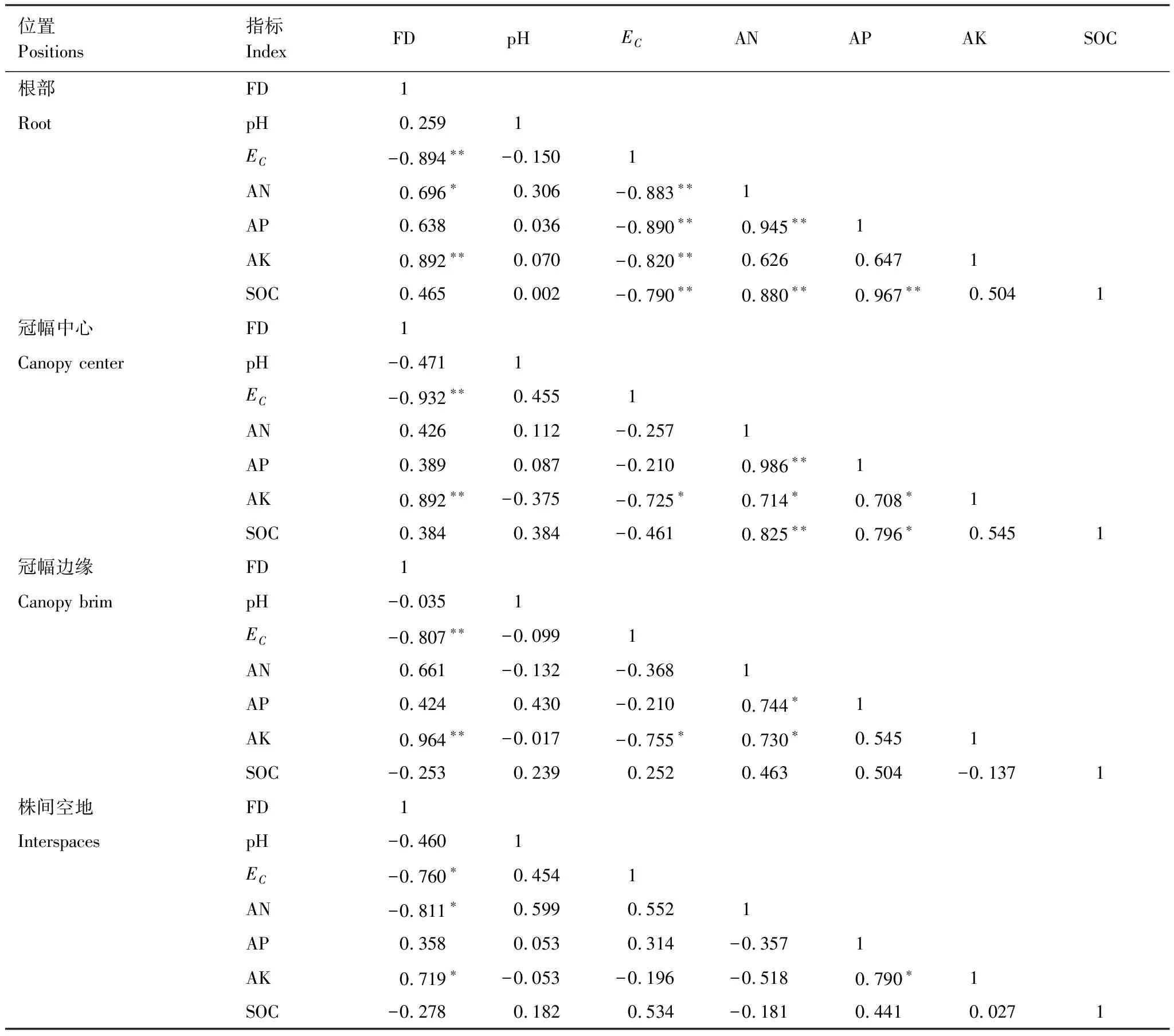
表3 柽柳植株周边不同位置土壤养分指标与盐碱指标相关系数矩阵
3 讨论
3.1不同密度柽柳林的土壤“盐谷”效应
土壤含盐量是影响黄河三角洲莱州湾湿地柽柳灌丛分布的主导因素[14],土壤盐分分布与地下水、植物种类、植物根系和根部分泌物等因素有关[21—22]。柽柳林分密度和土壤电导率呈极显著负相关(P<0.01)(表3),随林分密度增大,柽柳林土壤电导率显著降低。低密度柽柳林土壤电导率最大,可能是因为低密度柽柳林相邻的植株距离较远,林分郁闭度较低,导致地表蒸发作用增强,从而使得土壤盐分整体增大。而对滨海湿地不同密度柽柳林土壤调蓄水功能研究发现[19],土壤含盐量均值大小表现为低密度>高密度>中密度,与本研究低密度>中密度>高密度的结果不一致,可能是因为本研究中中密度林分由于其靠近盐池和海水养殖池,及盐水侧渗等因素,使其电导率值高于高密度。
随着距离柽柳植株根部越远,中、高密度柽柳林土壤电导率逐渐增加,表现为根部<冠幅中心<冠幅边缘<株间空地。柽柳冠下盐分含量低于冠外的现象,表明中、高密度柽柳林出现了“盐谷”效应,这与徐梦辰[6]对黄河三角洲湿地柽柳研究发现的柽柳冠下垂直深度0—10 cm、水平距离 0—150 cm处形成了一个典型的“盐谷”区域的研究结果一致。柽柳冠下的这种“盐谷”效应形成,可能是由于柽柳为泌盐植物,可通过根系较强的吸收作用将大量盐分储存于体内[5];其次,植物荫蔽作用以及柽柳发达根系的穿插作用等有助于土壤脱盐[23—24],所以柽柳冠下土壤含盐量小于株间空地,出现了“盐谷”效应。从富集率来看(表2),中、高密度柽柳林电导率富集率均小于1,说明中、高密度柽柳林对土壤盐分具有降低、迁移和疏导作用。
不同密度柽柳林土壤pH差异不显著(P>0.05),说明在黄河三角洲滨海滩涂研究区,密度对土壤pH并无显著影响。随着距离柽柳植株根部越远,低、中密度柽柳林土壤的pH表现出由根部向株间空地逐渐增加,但无显著差异(P>0.05)。柽柳根部pH较低可能与根部微生物呼吸作用产生CO2、根系分泌有机酸等有关[8,12],这与李从娟等[8]对荒漠灌木梭梭研究发现梭梭主根中心形成了一种高养分、低盐和低pH值的研究结果一致。从富集率来看(表2),低、中密度柽柳林土壤pH富集率更低,但差异不显著(P>0.05)。
3.2 不同密度柽柳林的土壤“肥岛”效应
土壤养分是土壤肥力的重要物质基础,是植物进行生命活动的基本保证[25—26]。董聿森等[5]研究发现,林分密度对黄河三角洲莱州湾柽柳林的土壤铵态氮、硝态氮、速效钾、速效磷影响显著,并提出中密度可以提高土壤肥力,这与本文研究表明的随林分密度增大,柽柳林土壤养分含量表现为先增加后减小的结论一致,说明林分密度可以显著影响土壤养分含量。土壤养分在中密度最高,主要是因为随着林分密度的增大,凋落物增加,同时较多的柽柳根系降低了土壤的紧实程度,土壤孔隙度增大,微生物运动活跃,土壤养分开始表现出增加的趋势[27]。但林分郁闭增加到一定值时,柽柳获取的光照资源减少,种间竞争激烈,凋落物分解速率和养分归还量下降,柽柳生长空间有限,导致柽柳生长受到一定限制,这时土壤养分随着林分密度增加而降低,所以中密度柽柳林土壤养分含量高于高密度。随着距离柽柳植株根部越远,中、高密度柽柳林土壤速效氮、速效磷含量表现为逐渐减小,即土壤养分含量由冠外向冠内汇集,这与干旱区河岸-荒漠过渡带梭梭(Haloxylonammodendron)与胡杨(Populuseuphratica)冠下“肥岛”效应[28]、南疆绿洲-荒漠过渡带胡杨“肥岛”的养分特征[29]研究结果一致。而低密度柽柳林植株周边不同位置的土壤养分含量差异不显著(P>0.05),表明中、高密度柽柳林出现“肥岛”效应,而低密度柽柳林未出现“肥岛”效应。低密度柽柳林未出现“肥岛”效应,主要是因为低密度柽柳林地表植被覆盖度低,冠幅较小(表1),使空气中的大气沉降颗粒、土壤表层风蚀物质和枯枝落叶等不易富集在冠下,所以导致其产生的养分在不易土壤中聚集[30—31];同时受海风吹蚀等影响,易将枯落物等吹散,使土壤养分不易在冠下聚集[32]。
中、高密度柽柳林养分富集率大于1(表2),表明中、高密度柽柳林对土壤养分具有富集作用,且中密度柽柳林养分富集率均大于高密度,说明中密度的“肥岛”效应强于高密度,即中密度柽柳林富集土壤养分的能力最高。不同密度柽柳林土壤养分富集率差异的原因可能与柽柳所处的分布位置、土壤环境、柽柳形态特征以及冠下有无低矮植物、生物量的空间格局等有关[11,33—35]。中密度柽柳林生长较好,柽柳基径和冠幅生长最大(表1),具有适宜的多枝半球状冠幅,能使枯枝落叶等富集在冠下,易导致养分在土壤中聚集较高[31];其次,土壤微生物数量、酶活性和土壤养分关系密切,中密度林分条件下,林下植被物种多样性较高[36—37],土壤变得疏松透气,有利于土壤中微生物和其他动物的活动,进一步促进“肥岛”的发育[21],所以中密度柽柳林“肥岛”效应强于高密度。不同密度柽柳林植株周边不同位置对土壤养分具有富集作用(表2),其中土壤养分富集率表现为根部>冠幅中心>冠幅边缘,说明柽柳根部“肥岛”效应最强,冠幅中心次之,冠幅边缘最差,这与黑河中游荒漠绿洲人工梭梭土壤养分特征[38]、北疆荒漠植被梭梭立地土壤养分“肥岛”特征[30]的研究结果一致。主要是因为随着到距离柽柳植株根部越远,其根系生物量逐渐减少[39],植物的吸收累积作用减弱,所以富集率逐渐减小。
4 结论
林分密度可显著影响柽柳林土壤盐分及养分含量。随林分密度增大,柽柳林土壤电导率显著降低,土壤速效氮、速效磷、速效钾、有机碳含量表现出先增大后减小。随距离柽柳植株根部越远,中、高密度柽柳林土壤电导率逐渐增加,即中密度、高密度柽柳林出现“盐谷”效应,低密度柽柳林未出现“盐谷”效应。不同密度柽柳林降盐作用表现为中、高密度较好,低密度较差。在中、高密度柽柳林,柽柳植株周边不同位置的土壤养分存在显著差异(P<0.05),低密度差异不显著(P>0.05),即中密度、高密度柽柳林出现“肥岛”效应,低密度柽柳林未出现“肥岛”效应。中密度柽柳林在土壤养分含量、富集率方面高于高密度,具有更强的“肥岛”和“盐谷”效应。
中密度柽柳林具有较好的提高土壤肥力作用,高密度次之,而低密度较差,不同密度柽柳林的降盐作用表现为中、高密度较好,低密度较差。从改善土壤养分和降盐抑碱的角度,建议黄河三角洲滨海滩涂柽柳合理的初植密度为4100 株/hm2。
