DENV-2对HUVECs自噬小体形成和自噬体-溶酶体融合及溶酶体降解途径的影响
2022-12-22胡盼苟小琴程瑶李志阳杨云巧何慧吴宁
胡盼, 苟小琴, 程瑶, 李志阳, 杨云巧, 何慧, 吴宁**
(1.贵州医科大学 基础医学院 化学与生物化学实验室, 贵州 贵阳 550004; 2.贵州医科大学 基础医学院 免疫学教研室, 贵州 贵阳 550004)
登革病毒(dengue virus, DENV)是一种隶属于黄病毒科黄病毒属的单股正链RNA病毒,编码产生3个结构蛋白(衣壳蛋白C、膜蛋白M和包膜蛋白E)和7个非结构蛋白(NS1、NS2a、NS2b、NS3、NS4a、NS4b和NS5),DENV主要通过白纹伊蚊和埃及伊蚊等媒介传播,已经成为世界上分布最广、引起发病最多的病毒之一,主要流行于全球热带和亚热带地区[1-3]。全球DENV感染者每年约3.9亿,DENV感染后尚无特异性治疗药物,大约12 500人因治疗不及时、不当等而死亡[4-5]。自噬是细胞内物质降解途径之一,包括自噬小体形成、自噬小体-溶酶体融合及溶酶体内自噬底物降解,完整自噬反应的发生对细胞内环境稳态的维持有着重要意义[6-7]。在病毒感染中,自噬起到双重作用,既可抑制病毒的复制、又能促进病毒的增殖[8-10]。已有研究证明DENV可诱导自噬的发生,为病毒复制提供有利场所[11],但DENV对自噬体-溶酶体融合以及溶酶体降解途径的影响未见报道。因此,本研究以原代人脐静脉内皮细胞(HUVECs)作为研究对象,探究DENV-2对HUVECs自噬反应途径的影响,在蛋白水平和细胞形态学水平揭示DENV-2对自噬流的作用机制,为进一步研究DENV在自噬水平的致病机制以及自噬的抗病毒反应提供实验依据。
1 材料与方法
1.1 材料
DENV-2 NGC株和C6/36由实验室液氮保存,HUVECs和ECM培养基购自美国ScienCell公司,RIPA裂解缓冲液购自上海碧云天生物技术有限公司,Anti-微管相关蛋白轻链3Ⅱ(LC3-Ⅱ,CST)和 Anti-自噬选择性底物p62购自日本MBL公司,Anti- 突触融合蛋白17(STX17)购自美国GeneTex公司,Anti-突触体相关蛋白29(SNAP29)和Anti-囊泡相关膜蛋白8(VAMP8)购自英国Abcam公司,兔抗β-actin购自武汉爱博泰克生物科技有限公司, ECL化学发光试剂盒购自上海碧云天生物技术有限公司;4 ℃低温高速离心机为美国ThermoFishe公司产品,ABI StepOnePlusTM实时荧光定量PCR仪为美国 ThermoFisher公司产品。
1.2 研究方法
1.2.1DENV-2感染HUVECs 取HUVECs种植于6孔板(5×104/孔),37 ℃,5% CO2培养箱中过夜,加入DENV-2孵育2 h,每30 min摇晃1次,最后1次弃去上清液,PBS洗涤1次,加入足量的培养液继续培养。
1.2.2透射电镜检测自噬小体 收集36 h空白组(仅培养,未加入DENV-2孵育)和DENV感染组细胞沉淀,缓慢加入0.5%戊二醛固定液,重悬后4 ℃静置10 min。12 000 r/min离心10 min,弃去上清,缓慢加入3%戊二醛固定液预固定,再用1%四氧化锇再固定,使用丙酮溶液逐级脱水,然后利用Ep812包埋剂进行包埋,超薄切片机切片和展片后醋酸铀染色15 min,再用柠檬酸铅室温下染色2 min,最后用JEM-1400FLASH透射电镜进行图像采集。
1.2.3Western blot法检测 LC3Ⅰ/Ⅱ、p62、STX17、SNAP29和VAMP8的蛋白表达量使用RIPA裂解缓冲液从HUVECs细胞中提取总蛋白, 并用BCA蛋白试剂盒测定蛋白浓度, 蛋白样品通过15%SDS-PAGE电泳,并转印到PVDF膜。5%脱脂奶粉封闭2 h。孵育一抗LC3(1 ∶1 000)、p62(1 ∶1 000)、STX17(1 ∶1 000)、SNAP29(1 ∶1 000)、VAMP8(1 ∶1 000)、β-actin(1 ∶50 000),HRP标记的山羊抗兔二抗(1 ∶8 000)孵育2 h,显影底物A液和B液等比例混合避光显影。使用Image J软件进行各蛋白条带灰度值分析。
1.2.4Lysotracker red 染色 根据试剂说明书提前使用完全培养基配制终浓度为75 nmol/L Lysotracker red工作液,37 ℃预处理,对空白组和DENV感染组用预冷的PBS洗涤2次,加入Lysotracker red工作液,37 ℃恒温箱孵育1 h;去除上清液,加入新鲜细胞培养液后置于倒置荧光显微镜下观察。
1.3 统计学处理

2 结果
2.1 DENV-2感染对HUVECs自噬小体形成的影响
透射电子显微镜结果显示,DENV-2感染HUVECs 36 h后胞质内形成双层膜包裹着未消化的细胞器的自噬小体(图1),表明DENV-2感染HUVECs后可诱导自噬小体的形成。
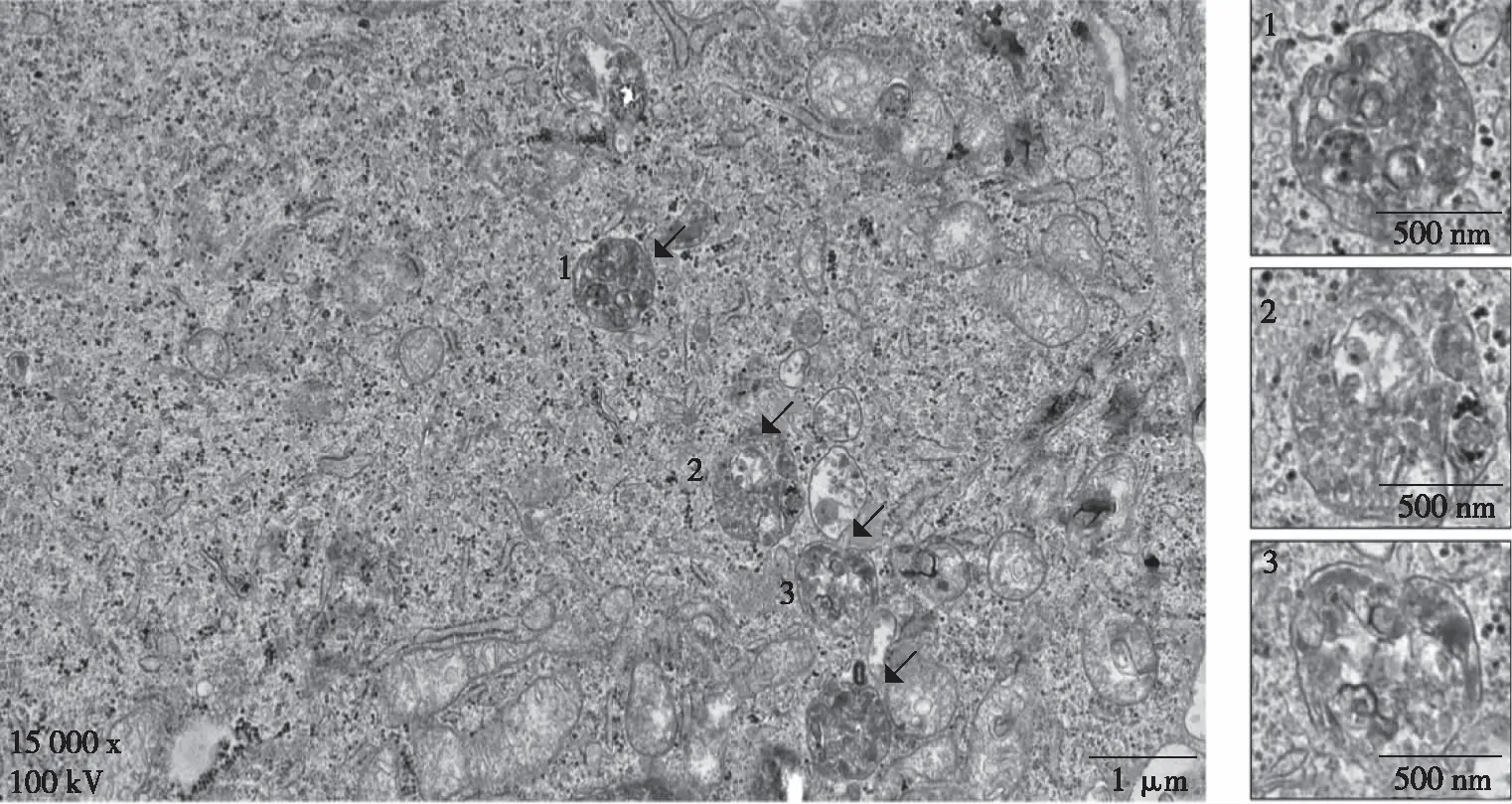
注:箭头所示为自噬体,左图比例尺为1 μm,右图为500 nm。图1 DENV-2感染HUVEC 36 h后的自噬体形成情况(透射电子显微镜)Fig.1 Autophagosome formation in HUVECs at 36 h after DENV-2 infection(transmission electron microscope)
2.2 DENV-2感染对自噬发生的影响
Western blot结果显示(图2),DENV-2感染HUVECs 12、24、36及48 h时,与对照(Control)组比较, DENV-2感染组和自噬激动剂雷帕霉素RAPA组LC3-Ⅱ蛋白表达水平升高,差异有统计学意义(P<0.05),但60 h时3组LC3-Ⅱ蛋白表达水平比较,差异无统计学意义(P>0.05);提示DENV-2感染 HUVECs 12、24、36及48 h均可激活自噬。
2.3 DENV-2对HUVECs自噬流的影响
Western blot检测DENV-2感染HUVECs 2、4、6、8、10、12、18、24、30、36及48 h时的p62蛋白表达水平变化。结果显示(图3),与对照(Control)组比较,DENV感染2、6、12 、30、36及48 h时的p62蛋白表达水平升高,差异有统计学意义(P<0.05),且36 h p62蛋白表达水平到达峰值,随后开始降低。提示DENV感染HUVECs 30 h后,自噬流受到阻碍。
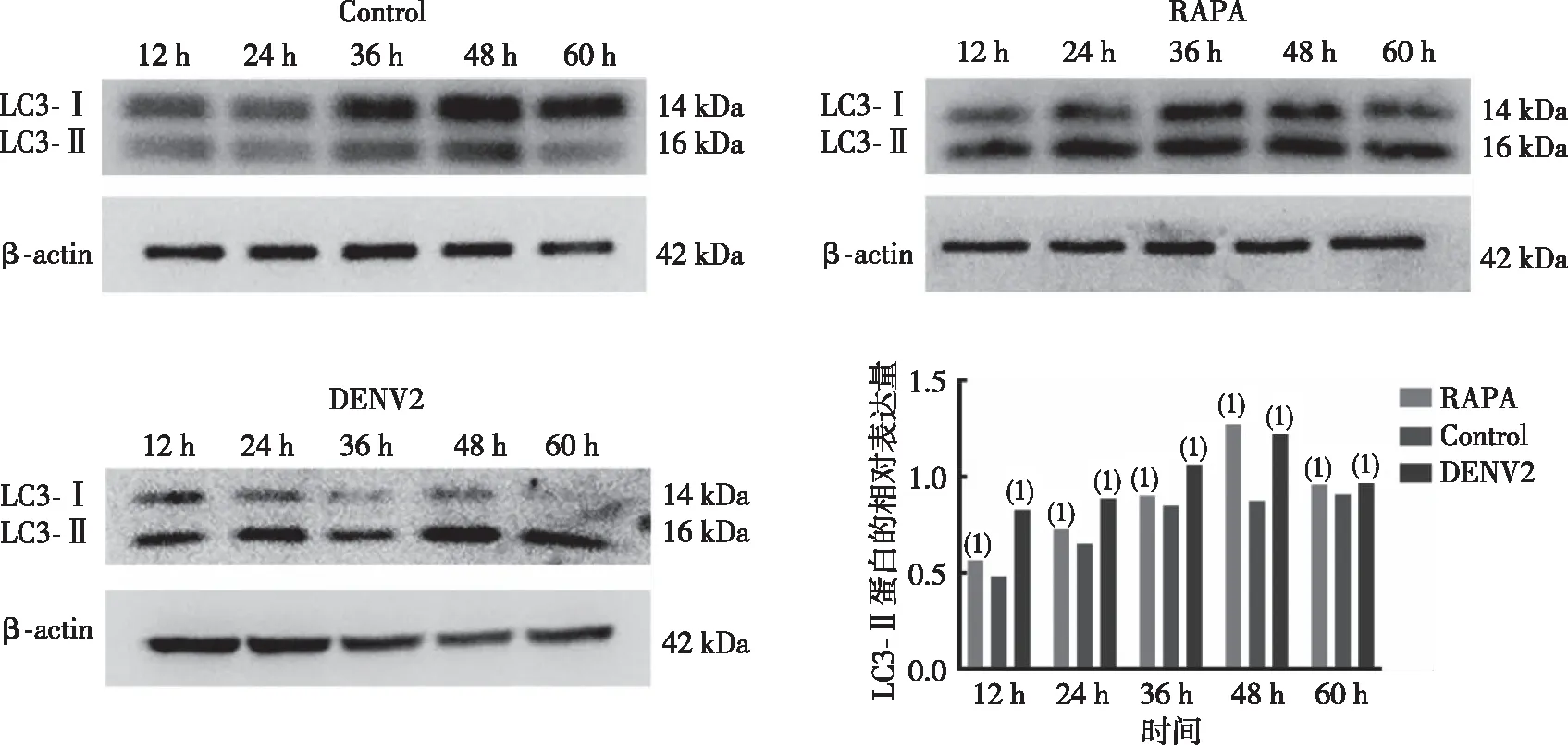
注:(1)与空白组比较,P<0.05。图2 DENV-2感染HUVEC不同时点的LC3蛋白表达Fig.2 Protein expression of LC3 in HUVECs at different time after DENV-2 infection
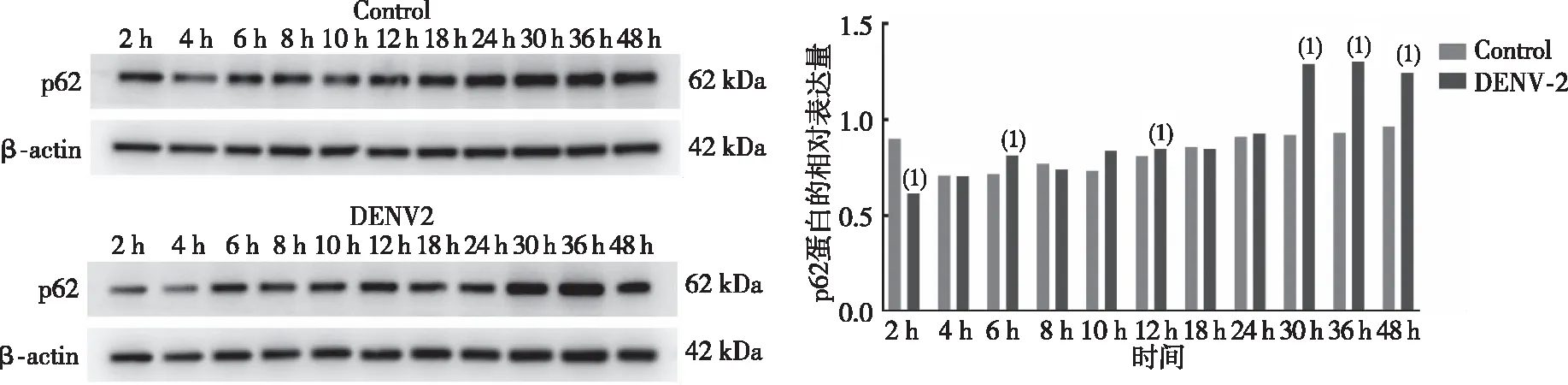
注:(1)与空白组比较,P<0.05。图3 DENV-2感染HUVEC不同时点的p62表达Fig.3 Expression of p62 in HUVECs at different time after DENV-2 infection
2.4 DENV-2对自噬小体-溶酶体融合的影响
为了进一步探究DENV-2阻断HUVECs晚期自噬流,利用Western blot检测DENV-2感染HUVECs 24、30、36及48 h时的STX17、SNAP29 、VAMP8蛋白表达水平,结果显示(图4),DENV-2感染组各时点的STX17蛋白表达水平低于对照(Control)组,差异有统计学意义(P<0.05);DENV-2感染24 h时的SNAP29、VAMP8蛋白水平高于空白组 (P<0.05),但30 h后SNAP29、VAMP8蛋白水平均低于空白组,差异有统计学意义(P<0.05)。提示DENV-2感染HUVECs可阻碍自噬体和溶酶体的融合。
2.5 DENV-2影响溶酶体正常酸化阻断自噬流
为了更进一步探究DENV-2阻碍自噬流的作用过程,采用Lysotracker red对DENV-2感染的HUVECs进行染色,倒置荧光显微镜观察细胞内溶酶体pH的变化。结果显示(图5),对照(Control)组细胞质中红色亮点较多,且染色明亮,而DENV-2感染组颜色黯淡,红色亮点明显减少。提示DENV-2感阻碍了HUVECs溶酶体正常酸化,从而阻断自噬流。进一步利用自噬晚期抑制剂CQ处理DENV-2感染的HUVECs,结果显示,DENV-2感染24、36及48 h时,DENV-2感染组和CQ组LC3-Ⅱ蛋白表达水平均高于对照(Control)组(P<0.05),且DENV-2感染组和CQ处理组的LC3-Ⅱ蛋白表达水平相近。提示DENV-2感染后期可以改变溶酶体内部的酸性环境,提高溶酶体的pH值。
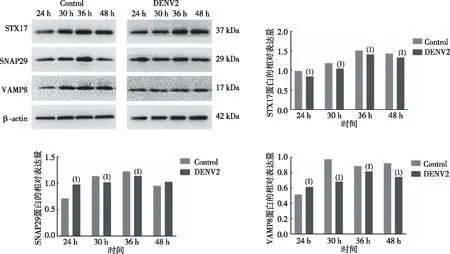
注:(1)与空白组比较,P<0.05。图4 关键自溶体融合蛋白STX17、SNAP29、VAMP8的相对表达Fig.4 Relative expression of key autolysosome fusion proteins STX17, SNAP29, and VAMP8
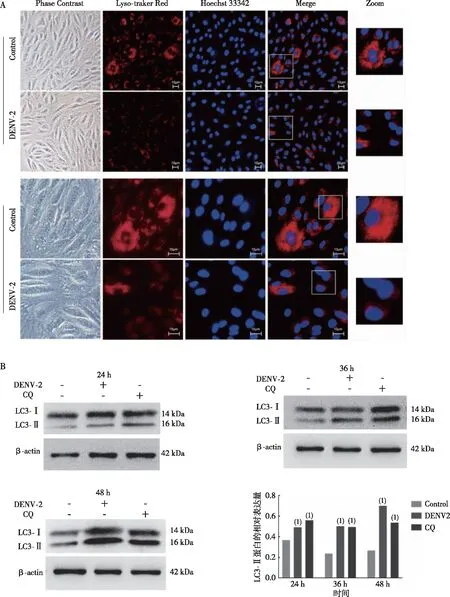
注:A为倒置荧光显微镜(100×)观察未感染或感染DENV-2的HUVEC,B为DENV-2感染24、36及48 h时,HUVEC中LC3-Ⅱ蛋白表达水平结果,(1)与空白组比较,P<0.05。图5 DENV-2影响HUVECS溶酶体的酸性环境Fig.5 Effect of DENV-2 on the acidic environment of HUVECS lysosomes
3 讨论
DENV感染引起的DHF、DSS是致命的主要因素,主要表现为内皮细胞损伤导致血浆渗漏,目前认为血浆渗漏和多种机制相关,如细胞自噬抗病毒反应、炎性介质的释放、内皮屏障蛋白缺失等[12-13]。细胞自噬作为维持细胞内环境稳态的重要途径,能够通过溶酶体降解胞内有害蛋白累积物、衰老的细胞器以及有害病原微生物等[14-16]。越来越多研究证明病毒感染和自噬各阶段相互作用,尤其在诱导自噬发生的自噬-溶酶体融合和溶酶体降解阶段[17-19]。DENV感染已证实能够促进自噬,但其阻碍自噬体和溶酶体融合的分子机制并未阐明,也不清楚DENV对完整自噬反应后续阶段溶酶体降解途径的影响[20]。因此,本研究旨在探讨DENV-2感染HUVECs后对自噬-溶酶体融合和溶酶体降解的影响。
Graybill等[18]的研究表明,水痘带状疱疹病毒(Varicella-zoster virus, VZV)介导自噬小体的形成,但阻断了晚期自噬流和自噬体-溶酶体融合。类似的,本研究通过透射电子显微镜可观察DENV-2感染后HUVECs自噬小体的形成,且Western blot结果显示DENV-2感染组LC3-Ⅱ表达水平增加,p62蛋白在DENV-2感染早期无明显变化,30 h后表达水平明显增加,提示DENV-2感染能够诱导自噬小体的形成,且能够抑制晚期自噬流的发生。而自噬流通畅与否和完整自噬过程的自噬体-溶酶体融合、溶酶体降解途径密切相关。自噬小体-溶酶体融合过程中,可溶性 N-乙基马来酰亚胺敏感因子附着蛋白受体(SNARE)发挥重要作用,SNARE系列蛋白中的STX17、SNAP29、VAMP8在自噬小体-溶酶体融合过程中紧密相连,三者缺一不可[21-22]。首先STX17被募集到成熟的自噬小体膜上,是融合的首要关键步骤,紧接着STX17的SNARE结构域和SNAP29相互作用,最后和VAMP8形成STX17-SNAP29-VAMP8复合物,促进自噬小体和溶酶体的融合,复合物中3个基因之一的敲低或敲除都会阻碍自噬小体和溶酶体的融合,从而阻断自噬流[23]。本研究利用Western blot检测DENV-2感染后期融合关键蛋白STX17、SNAP29、VAMP8的表达水平变化,结果显示,作为首要融合关键蛋白的STX17 在DENV-2感染后表达水平明显降低,而 SNAP29、VAMP8的表达水平在DENV-2感染24 h时高于对照(Control)组,但 24 h后SNAP29、VAMP8的表达水平均低与对照(Control)组,提示DENV-2感染晚期可阻断自噬-溶酶体融合,这与STX17、SNAP29、VAMP8蛋白低表达水平有关,且阻断融合和可能与DENV-2感染时间相关。DENV-2除了可抑制自噬-溶酶体融合,还可抑制溶酶体降解途径。溶酶体降解途径中溶酶体的酸化至关重要,一方面维持细胞的正常代谢与生长,另一方面有助于自噬对病原微生物的清除。当溶酶体功能紊乱时,即使自噬小体-溶酶体正常融合,自噬小体内底物却无法降解,反而为病毒的复制提供更多膜结构,同时使得定位于自噬小体膜上的病毒进一步抑制自噬小体底物降解[24-25]。本研究利用Lysotracker red对DENV-2感染的HUVECs进行染色,结果发现DENV-2可以抑制溶酶体的正常酸化,提示DENV-2感染可以抑制溶酶体自噬性降解。自噬晚期抑制剂CQ可以提高溶酶体的pH值从而改变溶酶体内部的酸性环境,导致自噬体的降解,引起LC3-Ⅱ表达增加。本研究通过使用CQ进一步处理对照(Control)组和DENV-2感染组,结果发现,与对照(Control)组相比,DENV-2感染组与CQ组的LC3-Ⅱ蛋白表达水平增加,且DENV-2感染组与CQ组的LC3-Ⅱ表达水平相近,提示DENV-2感染可以影响溶酶体酸性环境。
综上所述,本研究发现 DENV-2感染早期可激活HUVECs自噬,诱导自噬小体形成,感染晚期促使p62积聚,从而阻碍自噬流。DENV-2感染还可降低STX17、SNAP29、VAMP8的表达,证明DENV-2感染阻碍自噬小体-溶酶体融合过程,同时能够抑制溶酶体正常酸化从而抑制溶酶体自噬性降解。本研究虽揭示了DENV-2感染对HUVECs完整自噬过程的影响,但DENV-2感染如何干扰溶酶体酸化的分子机制尚不清楚,因此,本研究下一步计划从溶酶体酸化着手,进一步探究DENV-2感染对自噬的影响,从而为自噬抗病毒反应提供有力证据。
