不结球白菜BcLBD37基因的克隆及其对硝酸盐吸收的调节
2022-10-30纪雪洁王建军侯喜林许继伟邵嘉轩胡春梅
纪雪洁 王建军 侯喜林 许继伟 邵嘉轩 胡春梅
(南京农业大学园艺学院/作物遗传与种质创新国家重点实验室/农业农村部华东地区园艺作物生物学与种质创新重点实验室,江苏 南京 210095)
氮素作为植物生长发育所必需的营养元素之一,对植物的生长、发育和代谢过程至关重要[1-2],其中NO3-和NH4+作为氮素存在的两种主要形式被植物吸收利用[3-4]。由于植物吸收作用、地表径流和微生物反硝化作用的影响,土壤中硝酸盐的含量逐渐降低,为了提高作物的产量,不得不额外施加氮肥。但长期施用氮肥不仅使作物的产量降低,而且会导致土壤酸化,不利于绿色农业的发展[5-6]。因此,提高氮肥的利用效率和降低施氮量,已成为当今绿色农业可持续发展的必然要求[7-8]。
植物体内对硝酸盐的吸收主要通过高亲和硝酸盐转运系统和低亲和硝酸盐转运系统进行[9]。当外界硝酸盐的浓度超过1 mmol·L-1时,主要通过高亲和硝酸盐转运系统(high affinity transport system, HATS)发挥作用;而当硝酸盐浓度低于1 mmol·L-1时,主要通过低亲和硝酸盐转运系统(low affinity transport system, LATS)发挥作用[10]。有研究表明,NRT1[11]和NRT2[12-13]基因家族分别参与LATS和HATS。在拟南芥(Arabidopsisthaliana)中AtNRT1.1基因编码的蛋白是一种双亲和硝酸盐转运蛋白并充当硝酸盐传感器的角色参与硝酸盐代谢过程,该基因主要在植物的根系中表达[14-15]。AtNRT1.5[16]和AtNRT1.7[17]基因的表达受到硝酸盐的诱导,并参与调控植株根系或地上部分对硝酸盐的吸收、转运和氮素分配等过程。AtNRT2.1[18]、AtNRT2.5[19]主要参与根系对硝态氮的吸收。除了NRT基因家族以外,拟南芥AtANR1通过编码MAD-box转录因子,局部激活硝酸盐代谢途径,进而调控植株根系的生长[20-23]。作为拟南芥NIN-like家族成员之一,NLP7转录因子参与硝酸盐代谢和氮饥饿的响应过程[24]。苹果(MalusdomesticaBorkh)的MdNLP7基因通过激活MdNIA2和MdNRT1.1的表达促进氮的吸收和同化,此外MdNLP7可以通过增加过氧化氢酶活性来调节H2O2含量,进而影响硝酸盐的利用效率[25]。可见NRT基因家族直接参与植物氮代谢的吸收、转运以及固化,ANR1和NLP7转录因子同样可以响应硝态氮的吸收,并参与调控氮代谢过程。
外侧器官边界域(lateral organ boundaries domain, LBD)转录因子作为高等植物特有的一类转录因子,具有1个高度保守的外侧器官边界(lateral organ boundaries, LOB)结构域[26],根据LOB结构域的差异,将LBD转录因子分为两类,第Ⅰ类LBD转录因子除了含有类锌指结构(CX2CX6CX3C)外,在LOB结构域的末端形成类亮氨酸拉链结构(LX6LX3LX6L),该类转录因子主要参与蛋白质互作过程[27],第Ⅱ类LBD转录因子仅含有1个保守的类锌指结构区,可能与DNA的绑定有关[28]。第Ⅱ类LBD转录因子受硝酸根离子的强烈诱导并影响氮代谢,例如在大麦(HordeumvulgareL.)中,经外源硝酸盐处理后HvLBD5/14基因表达量上调,并且该基因过表达导致拟南芥叶片中硝酸还原酶活性以及硝酸盐含量增加[29]。苹果MdLBD13基因也特异性响应硝酸盐,并参与到氮代谢的调控过程[30]。而在不结球白菜(Brassicarapassp. Chinensis)中,LBD转录因子家族成员是否同样参与硝酸盐代谢的调控仍鲜见报道,其与硝酸转运系统的作用关系仍有待研究。
本研究前期以紫色不结球白菜单株为材料,化学诱变获得其绿色突变体NJZX1-0[31]。该绿色突变体BcLBD37基因的表达水平显著低于紫色品系NJZX3-4,而其硝酸盐代谢水平显著高于紫色品系。为探讨LBD基因与硝酸盐代谢的关系,本研究以紫色不结球白菜NJZX3-4为材料,克隆第Ⅱ类LBD基因家族成员BcLBD37,通过外源NO3-离子处理,分析硝酸盐对该基因表达水平的影响,并通过转基因技术,将该基因导入到拟南芥中,鉴定不结球白菜LBD基因的功能,同时,利用病毒介导的基因沉默技术(virus induced gene silencing, VIGS)沉默紫色不结球白菜NJZX3-4的BcLBD37基因,进一步验证该基因对不结球白菜硝酸盐吸收的影响,旨在为研究该基因对硝酸盐代谢的调节机理提供理论依据。
1 材料与方法
1.1 试验材料
紫色不结球白菜纯系品种NJZX1-3由南京农业大学白菜系统生物学实验室提供。首先将该品种的种子用酒精消毒,无菌水冲洗,室温催芽3 d左右,移至32孔穴盘中,放置于人工气候室中进行培养。培养的环境条件为:光照/黑暗16 h/8 h,温度22℃/18℃。
1.2 不结球白菜BcLBD37基因克隆
以紫色不结球白菜NJZX3-4叶片为材料提取RNA,利用反转录试剂盒(TaKaRa公司,北京)反转录为cDNA,作为克隆的模板。PCR总体系为20 μL:模板1 μL,正反引物各1 μL,2×HiQPCR MIX 10 μL, ddH2O 7 μL。扩增程序为:95℃预变性4 min ;95℃变性30 s,56℃退火30 s,72℃延伸1 min,35个循环;72℃终延伸10 min 。PCR产物利用胶(湖南艾科瑞生物工程有限公司)回收试剂盒回收目的片段,连接PEASY-Blunt载体,经大肠杆菌转化后,挑取单菌落放入含有卡那抗生素的LB液体培养基活化,将菌液送至南京生工生物公司进行测序,获得目的基因片段。
1.3 不结球白菜BcLBD37蛋白的序列分析
利用BioXM 2.7对BcLBD37基因进行开放阅读框(open reading frame, ORF)查找、翻译;利用DNAMAN7程序进行多重序列比对;同时用WebLogo(http://weblogo.berkeley. edu/logo.cgi)在线程序分析LBD37转录因子蛋白结构域的保守位点。
1.4 不结球白菜BcLBD37蛋白的亚细胞定位
利用PRI101-BcLBD37-GFP采用冻融法转化到农杆菌GV3101,涂布在含卡那霉素和利福平的抗性的溶菌肉汤(Luria-Bertani, LB)固体培养基上,挑取单克隆活化,获得悬浮液,用1 mL去掉针头的的灭菌注射器注射一个月月龄的烟草叶片背部,3 d后利用LSM 780激光共聚焦显微镜(Zeiss,德国)确定BcLBD37蛋白在植物细胞中的位置并拍照。
1.5 过表达植株的获得
利用PRI101-BcLBD37-GFP悬浮菌液侵染野生型拟南芥花序,每周侵染1次,重复3次,刚侵染完收获的种子记为T0代。放入37℃培养箱中,干燥1周,之后将T0代种子播种在含有卡那霉素、特美汀抗性的固体Murashige and Skoog(MS)培养基上,筛选阳性苗,绿色荧光蛋白标记(green fluorescent protein,GFP)、聚合酶链式反应(polymerase chain reaction, PCR)、实时荧光定量PCR(quantitative real-time PCR, qRT-PCR)以及蛋白免疫印迹法(Western Blot)验证,然后将阳性苗单株收种直至获得纯合的T3代种子。
1.6 BcLBD37基因沉默植株的获取
载体构建参照杨学东等[32]的方法。选择40 bp的序列,5′-C G A A G A C G G T G G C G T G G C C T T G A G C A T C G G C G G T T T C T AT-3′,另一条链反向互补,共80 bp,送至南京金斯瑞生物科技有限公司进行合成。PTY载体经过NdeI酶切后,用T4连接酶将该PTY载体与上述80 bp的DNA片段过夜连接。经过大肠杆菌转化后,挑取单菌落,测序。提取PTY-BcLBD37质粒,使用金粉包裹质粒,利用基因枪法轰击紫色不结球白菜的叶片,并且轰击不含质粒的空载PTY-S作为对照,20 d后,观察植株的发病情况,取0.1 g发病叶提取RNA,用于qRT-PCR检测。
1.7 外源硝酸盐对紫色不结球白菜的处理
将紫色不结球白菜NJZX3-4催芽后,播种在32孔穴盘中,放置于气候室中进行培养,光照/黑暗时间为14 h/10 h,温度为22℃。待植株生长至五叶期,选择生长健壮的幼苗,分成2组,清洗根部,转移至不含有氮元素的霍格兰林营养液中,缓苗2 d。将2组霍格兰林营养液分别配制为5 mmol·L-1的KNO3和5 mmol·L-1的KCl溶液,将2组幼苗分别放置于上述营养液中,在放置后的0、1、3、6、12、24、48、72 h进行取样,放置于-80℃冰箱中保存,每个处理设置3组重复。
1.8 实时荧光定量PCR表达分析
根据克隆所得的BcLBD37基因cDNA全长序列,采用在线软件Primer-BLAST设计正、反引物,引物序列见表1,Actin基因作为内参。试验步骤及用量参考SYBR Premix Ex Taq TMNJZX1-0Ⅱ(TaKaRa,北京)试剂盒说明书。反应总体系为20 μL:正、反引物各0.4 μL,DNA模板2 μL,Hieff® qPCR SYBR Green Master Mix 10 μL,ddH2O 7.2 μL。反应程序:95℃预变性 5 min,95℃变性 10 s,60℃退火30 s,共40个循环。每个反应设置3次重复。采用2-ΔΔCT的方法分析数据,利用IBM SPSS Statistics 24软件进行误差分析和差异显著性分析。

表1 本研究所用引物序列Table 1 Primer sequences in this study
1.9 硝酸盐含量和硝酸还原酶活性的测定
使用BC1500硝态氮检测试剂盒(Solarbio,北京),测定硝态氮含量。使用BC0080硝酸还原酶活性检测试剂盒(Solarbio,北京)测定硝酸还原酶活性。
1.10 数据分析
采用Excel 2016和SPSS 21.0统计软件进行数据统计和方差分析,采用2-ΔΔCT的方法进行基因表达分析,用Duncan检验法进行显著性差异(P<0.05)分析;采用Origin 2018、Excel 2016和Photoshop 2018作图。
2 结果与分析
2.1 不结球白菜BcLBD37基因的克隆以及序列分析
提取紫色不结球白菜NJZX3-4叶片RNA,反转录后进行BcLBD37基因PCR扩增,得到1个840 bp的目的片段(图1),含有837个ORF,编码279个氨基酸,其中含有酸性氨基酸44个,碱性氨基酸34个。

注:*表示终止密码子。Note: *represents the stop codon.图1 不结球白菜中BcLBD37基因的核苷酸序列及其编码的氨基酸序列Fig.1 The nucleotide and deduced amino acids sequence of BcLBD37 gene in Brassica campestris ssp. Chinensis
2.2 BcLBD37蛋白的同源性比较及系统进化树分析
将不结球白菜BcLBD37蛋白序列与拟南芥中AtLBD37、AtLBD38、AtLBD39的蛋白序列进行同源比对,结果发现不结球白菜BcLBD37与拟南芥AtLBD37、AtLBD38、AtLBD39蛋白序列都含有高度保守的LOB结构域,仅含有1个C2CX6CX3C基序,不含有类亮氨酸拉链模型(LX6LX3LX6L),并且这4个蛋白序列在C末端都含有1个高度保守的LINLF基序(图2)。
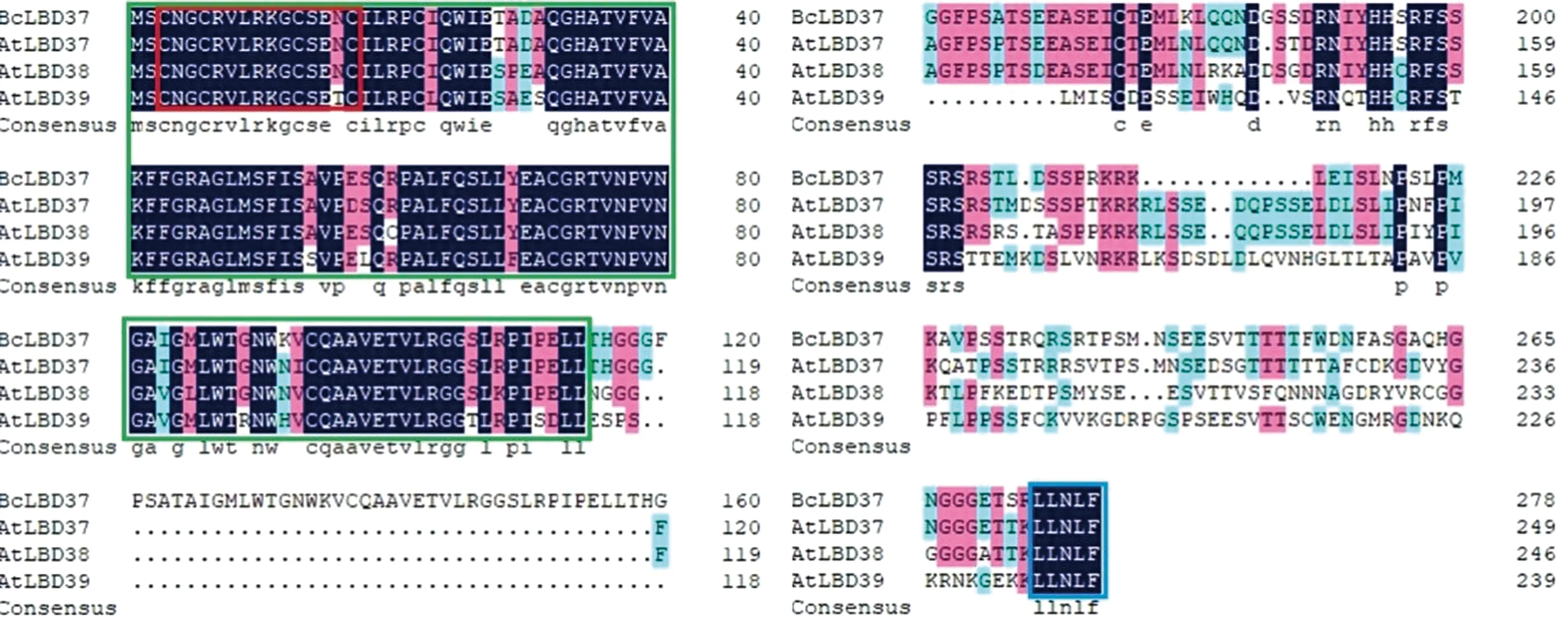
注:绿色框线内为LOB结构域;红色框线内为C2CX6CX3C基序;蓝色框线内LINLF基序。Note: LOB domain is in the green box. C2CX6CX3C motif is in the red box. LINLF motif is in the blue box.图2 不结球白菜BcLBD37与拟南芥Class Ⅱ 类LBD蛋白序列比对Fig.2 Sequence alignment of class Ⅱ LBD protein between non-heading Chinese cabbage BcLBD37 and Arabidopsis thaliana
2.3 BcLBD37蛋白的亚细胞定位检测
PRI101-BcLBD37荧光表达载体在绿色荧光蛋白(green fluorescent protein,GFP)通道下可以被激发,发出明显的绿色荧光信号。细胞核maker用来标识细胞核的位置,在红色荧光蛋白(red fluorescent protein,RFP)通道下发出明显的红色荧光信号。利用激光共聚焦显微镜观察注射3 d后的烟草叶片,发现绿色荧光信号和红色荧光信号重叠细胞核位置(图3)。由上述结果可知,BcLBD37蛋白定位于细胞核。
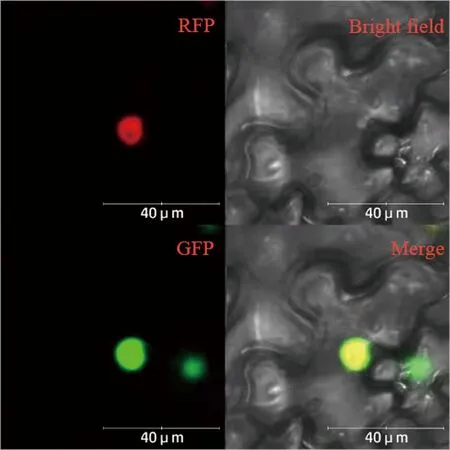
注:GFP:GFP 激光下的绿色荧光信号;RFP:RFP激光下的红色荧光信号;Bright field:明场;Merge:GFP、RFP和明场的叠加。Note: GFP: The green fluorescence signals. RFP: The red fluorescence signals. Merge: The merge of GFP, RFP and bright field.图3 BcLBD37蛋白在本氏烟草叶片细胞中的定位Fig.3 Subcellular localization of BcLBD37 in leaves of Nicotiana benthamiana
2.4 BcLBD37基因的组织特异性表达以及对外源硝酸盐的响应
提取紫色不结球白菜NJZX3-4植株的根茎叶,通过qRT-PCR分析BcLBD37基因在不同组织中的表达差异,结果显示BcLBD37在不结球白菜的各个部位均有表达,且在根中的表达量最高,在叶中的表达量最低(图4-A)。紫色不结球白菜NJZX3-4分别用 5 mmol·L-1KCl和5 mmol·L-1KNO3处理不同时间,结果显示,当用KNO3处理3 h时,根系中BcLBD37基因的表达量达到最大值,约为0 h时的10.6倍(图4-B)。而叶片中BcLBD37基因的表达量在用KNO3处理24 h时达到最大值,约为0 h时的8.6倍(图4-C)。用KCl处理后,BcLBD37基因在根和叶中的表达量均无显著差异。以上结果表明,NO3-对不结球白菜BcLBD37基因的表达具有诱导效应。
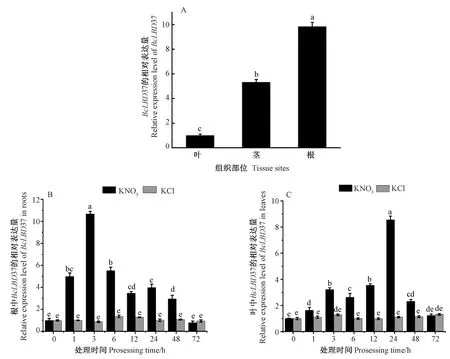
注:不同字母表示不同植株中基因表达量在P<0.05水平差异显著。下同。Note: Different letters have shown significant differences in gene expression in different plants at 0.05 level. The same as following.图4 不结球白菜BcLBD37基因的组织特异性表达及对外源硝酸盐的响应Fig.4 Tissue-specific expression of BcLBD37 gene in non-heading Chinese cabbage and its response to nitrate
2.5 BcLBD37基因过表达拟南芥植株的鉴定
为了验证BcLBD37基因的功能,利用农杆菌介导法对拟南芥进行遗传转化。利用卡那霉素和利福平抗生素筛选出阳性苗,提取DNA用35 S-F/BcLBD37-R进行PCR验证,初步鉴定出7个过表达阳性苗(图5)。通过qRT-PCR对其中5个转基因株系进行表达量测定,结果显示与野生型相比,转基因株系OE-1#、OE-3#、OE-5#的表达量显著上调,同时将这3个株系进行蛋白水平的检测,均检测到了GFP标签。因此选择OE-1#、OE-3#、OE-5#株系进行功能验证。
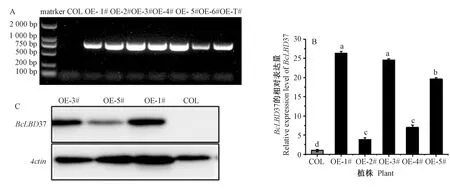
注:A:基因组qPCR;B:qRT-PCR检测;C:蛋白免疫印迹。COL:野生型拟南芥;OE-1~7#:转基因拟南芥。下同。Note: A: Genome PCR. B: The qRT-PCR test. C: Western blotting. COL: Wild-type Arabidopsis thaliana. OE-1~7#: Transgenic Arabidopsis thaliana. The same as following.图5 BcLBD37过表达植株的验证Fig.5 Validation of BcLBD37 overexpressed plants
2.6 转基因拟南芥硝态氮以及硝酸还原酶活性的测定
测量转基因拟南芥与野生型拟南芥中硝态氮的含量以及硝酸还原酶活性,结果显示,与野生型相比,OE-1#、OE-3#、OE-5#株系硝态氮含量(图6-A)和硝酸还原酶活性(图6-B)均显著降低,其中OE-1#株系硝态氮含量最低,为野生型的57%,OE-3#株系硝酸还原酶活性最低,为野生型的55%,可知BcLBD37基因过表达抑制硝酸盐的吸收、转运和同化。
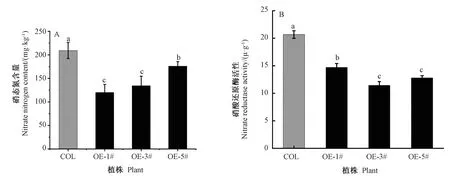
图6 转基因拟南芥与野生型拟南芥硝态氮含量与硝酸还原酶活性对比Fig.6 Comparison of nitrate nitrogen content and nitrate reductase activity between transgenic Arabidopsis thaliana and wild-type Arabidopsis thaliana
2.7 转基因拟南芥硝酸盐代谢相关基因的表达量
测定转基因植株中硝酸盐吸收转运和同化相关基因的表达量,结果显示转基因植株中编码硝酸还原酶活性的NIA1、NIA2基因,以及编码硝酸盐代谢途径相关基因NRT1.1、NRT1.7、NRT2.1、NRT2.5的表达量与野生型相比均显著下调(图7),由此得出BcLBD37基因可通过抑制硝酸盐吸收同化相关基因的表达,进而降低硝酸盐的吸收效率。
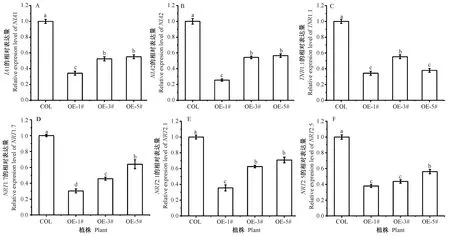
图7 转基因拟南芥与野生型拟南芥硝酸盐代谢相关基因的表达量Fig.7 Expression levels of genes related to nitrate metabolism in transgenic Arabidopsis thaliana and wild-type Arabidopsis thaliana
2.8 BcLBD37基因沉默对不结球白菜硝酸还原酶活性和硝酸盐含量的影响
qRT-PCR结果显示,该基因的表达量与空载PTY-S相比,明显降低,说明不结球白菜中BcLBD37基因被沉默(图8-A)。测定基因沉默植株PTY-2/4和空载植株PTY-S中硝酸还原酶活性和硝态氮含量,发现PTY-2/4植株硝酸还原酶活性为空载植株的1.94、1.69倍(图8-B),硝态氮含量为空载植株的1.30、1.37倍(图8-C)。BcLBD37基因表达量降低,硝态氮含量以及硝酸还原酶活性增加,说明BcLBD37基因的表达对硝态氮合成以及硝酸还原酶活性产生影响。
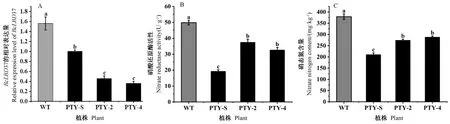
图8 基因沉默处理后BcLBD37基因的表达量以及沉默植株中硝酸还原酶活性与硝态氮的含量的测定Fig.8 The expression of BcLBD37 gene after gene silencing treatment and the determination of nitrate reductase activity and nitrate nitrogen content in silenced plants
2.9 BcLBD37基因沉默对不结球白菜硝酸盐代谢相关基因表达量的影响
测定BcLBD37基因沉默植株PTY-2/4中NIA1、NIA2、NRT1.1、NRT1.7、NRT2.1、NRT2.5基因的表达量。结果显示,与空载植株PTY-S相比,沉默植株NIA1、NIA2、NRT1.1、NRT1.7、NRT2.1、NRT2.5的表达量显著上调,分别是空载植株的1.25、1.19、1.50、1.21、1.25、1.43倍(图9)。BcLBD37基因沉默,硝酸盐代谢相关基因表达量增加,表明BcLBD37基因抑制了硝酸盐代谢相关基因的表达。
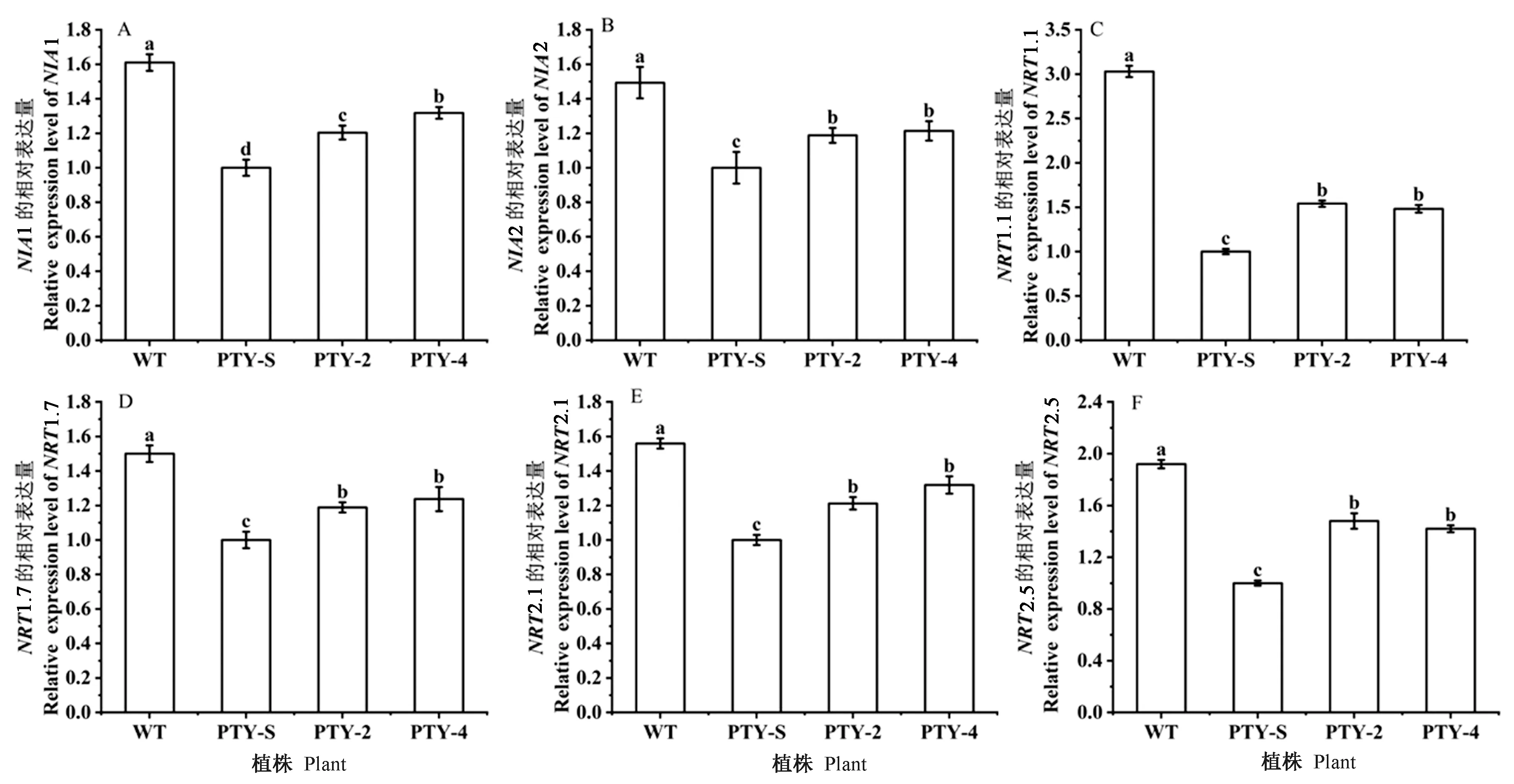
图9 BcLBD37基因沉默对硝酸盐代谢相关基因表达量的影响Fig.9 Effect of BcLBD37 gene silencing of nitrate metabolism related gene expression
3 讨论
LBD转录因子广泛存在于高等植物中,在植物的生长发育、胁迫应答以及次生代谢过程中发挥着至关重要的作用[33]。在拟南芥中,LBD转录因子AtLBD38/39属于第Ⅱ类LBD转录因子,已经被证实参与氮素介导的硝酸盐代谢过程[34]。本研究的同源比对结果显示,BcLBD37蛋白与拟南芥AtLBD37/38/39蛋白同源性较高,且BcLBD37基因仅含CX2CX6CX3C类锌指结构,不含类亮氨酸拉链模型,说明BcLBD37与拟南芥AtLBD37/38/39同属第Ⅱ类LBD转录因子家族,由此推测BcLBD37基因可能与拟南芥AtLBD38/39基因的生物学功能相似。
硝酸盐作为一种信号分子参与植物的生长发育与次生代谢过程[35]。有研究结果显示外源硝酸盐诱导大麦(HordeumvulgareL.)基因HvLBD5/14[29]、MdLBD13[30]的表达。为了探究硝酸盐对不结球白菜BcLBD37基因的表达是否具有诱导效应,本研究用5 mmol·L-1的KCl与5 mmol·L-1的KNO3处理紫色不结球白菜,结果发现BcLBD37在根和叶中的表达量均显著上调,而KCl处理基因的表达量无显著变化。表明BcLBD37基因特异性响应硝酸盐,硝酸盐诱导不结球白菜BcLBD37基因的表达。
前人研究发现,HvLBD5/14基因在外源硝酸盐处理后基因的表达量上升,HvLBD5/14转基因拟南芥苗期叶片中硝酸盐和蛋白质的含量增加[29],苹果MdLBD13基因过表达后硝酸还原酶活性和硝态氮含量却显著降低[30],拟南芥AtLBD38/39基因同样表现出对氮代谢的抑制作用[34]。通过对水稻(OryzasativaL.)OsLBD37基因的代谢组学和转录组学进行分析发现,OsLBD37基因与水稻氮代谢过程相关,与AtLBD37基因在氮代谢调控上具有相同的生物学功能[36]。HvLBD5、HvLBD14、MdLBD13、OsLBD37同属于第Ⅱ类LBD转录因子家族,但是对氮代谢的调控效应不同。本研究通过将不结球白菜BcLBD37基因转入拟南芥,发现转基因植株中硝态氮含量与硝酸还原酶活性均显著降低,同时编码硝酸盐代谢相关基因的表达量也显著下调。而与空载植株相比,不结球白菜的BcLBD37基因沉默植株硝酸还原酶活性以及硝态氮含量均显著升高,且编码硝酸盐吸收转运与同化相关基因的表达量显著上调。根据以上结果可知,BcLBD37基因属于第Ⅱ类LBD转录因子家族成员,与拟南芥AtLBD38/39、HvLBD5/14调节氮代谢功能上具有相似的效应。因此说明,BcLBD37基因可通过抑制硝酸盐代谢相关基因的表达,进而对硝态氮的合成与硝酸还原酶的活性产生影响。
4 结论
本研究从紫色不结球白菜纯系NJZX3-4中克隆得到基因BcLBD37,该基因含有837个开放阅读区,编码279个氨基酸;亚细胞定位结果显示,该基因编码的蛋白定位于细胞核;硝酸盐处理的结果显示,硝酸根离子对于BcLBD37基因的表达具有诱导效应;表达结果显示,BcLBD37基因对硝酸盐吸收具有抑制作用。
