混合振荡神经元网络中反相干共振向相干共振的转迁*
2022-10-29吴媛梦贾雁兵
吴媛梦 贾雁兵
(河南科技大学 数学与统计学院,洛阳 471000)
引言
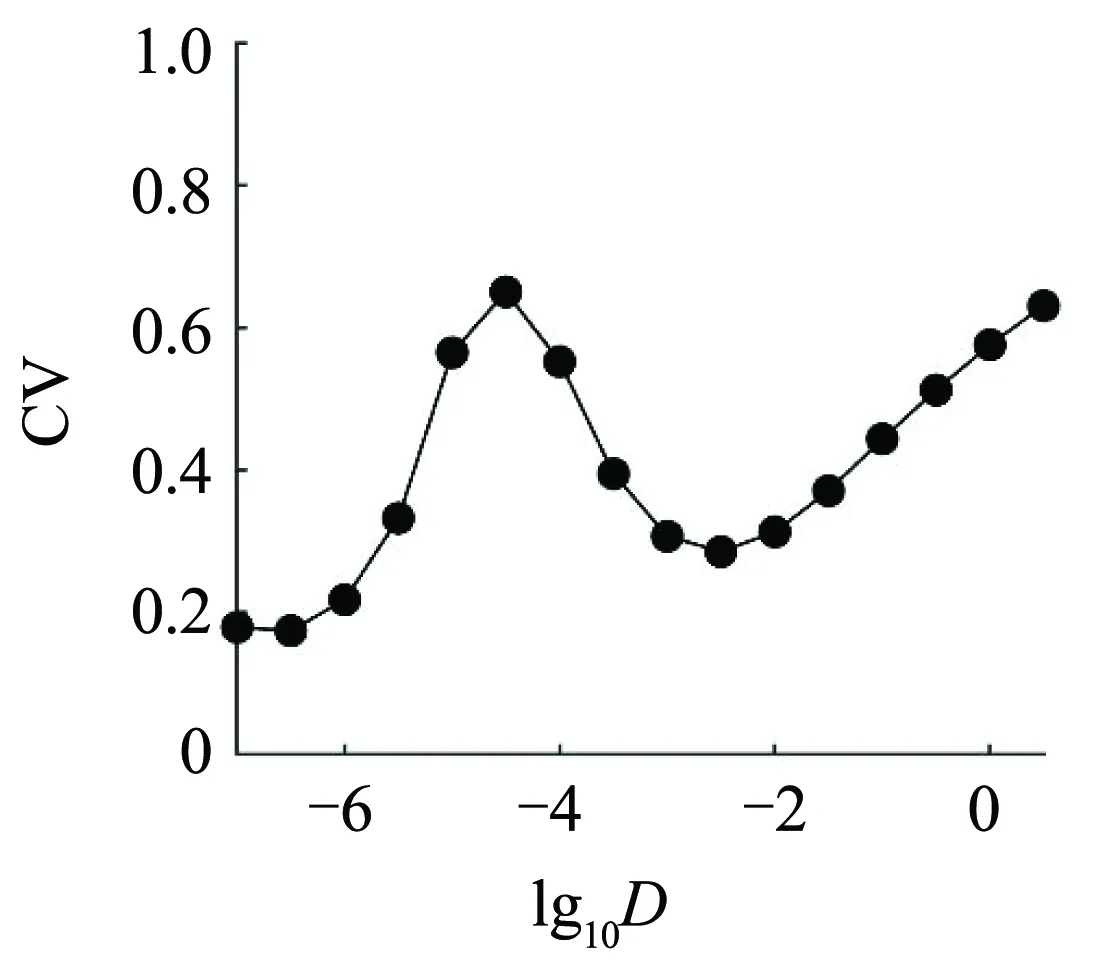
噪声诱导非线性系统的复杂随机动力学是动力学与控制学科的重要内容,如随机共振(stochastic resonance)和相干共振(coherence resonance)[1-8]已经从作为违反直觉行为的代表到如今被广泛接受.其中,随机共振是指含噪声的非线性系统对弱周期信号的响应在中等强度的噪声下得到优化的现象[3],相干共振现象则是指含噪声的非线性系统在无弱周期信号的作用下表现出的类随机共振的现象[3].在神经系统中,噪声源于离子通道的随机开关和神经递质的随机释放等因素,其可以诱导随机共振和相干共振[9-12],在感觉信息处理、运动控制和临床康复中发挥着重要作用.神经系统的相干共振经常用放电峰峰间期(interspike interval,ISI)的变差系数随噪声强度先增加后降低来刻画[3].
虽然随机共振和相干共振在神经系统的阈值或分岔点附近是普遍存在的,但是反相干共振(anti-coherence resonance)或逆随机共振(inverse stochastic resonance)也在亚临界Hopf分岔点附近被发现[13-15].与相干共振不同,反相干共振是指适中强度的噪声会压制放电,例如放电峰峰间期随噪声强度的增大而先增大后减小[16].逆随机共振则指放电频率在适中噪声强度下达到最小[13],与相干共振的放电频率一般随着噪声强度的增大而增大不同.反相干共振和逆随机共振扩展了神经系统的复杂随机动力学.
在单神经元模型中,噪声类型、噪声强度、神经元电活动和分岔类型等决定着相干共振或反相干共振的动力学特征.例如,众多研究关注了噪声从平衡点分岔附近的静息态诱发相干共振,以及从亚临近Hopf分岔点附近的共存行为诱发反相干共振[13-15].因为神经系统中的神经元并不是孤立存在的,而是相互耦合构成神经元网络.因此神经元网络模型研究中也关注了随机共振和相干共振[17],而且研究了网络的多种因素(如网络结构、耦合强度和突触时滞等)及神经元的动力学行为对随机共振和相干共振的重要影响[18-24].例如,适中的平均度能增强神经元网络的随机共振[18],相位噪声能诱导神经元网络产生两次相干共振[20],时滞能减弱神经元网络的相干共振[21],抑制性自突触能增强神经元网络的相干共振[24].然而,以上大部分研究关注噪声诱导位于分岔点附近的静息态产生的随机共振或相干共振.
近期,能够产生相干共振和反相干共振的条件得到了扩展.有研究发现簇放电,特别是混合振荡的簇放电也能产生相干共振[25,26],单个峰和阈下振荡交替的混合振荡可以产生反相干共振到相干共振的转迁[16].实际上,该混合振荡之所以能发生反相干共振向相干共振的转迁,是因为该混合振荡相对特殊的动力学行为.该混合振荡的动力学行为是较长的阈下振荡和单个峰的交替,在一定程度上具备“共存”行为的特征,因而能够发生反相干共振;而该混合振荡的阈下振荡远远多于放电峰,在很大程度上具有阈下振荡的特征,接近放电阈值,可以产生相干共振.因此,混合振荡是能够产生复杂随机动力学的重要放电行为.
混合振荡是指具有大幅值和具有小幅值振荡交替出现的一种放电模式,广泛存在于实际神经系统.例如,生理学实验研究发现:垂体催乳素细胞[27]和大鼠内侧隔核中的氨基丁酸能神经元[28]能产生混合振荡;在不同的抗癫痫药物作用下,内嗅皮层中的椎体神经元会产生不同类型的混合振荡[29].此外,混合振荡与神经系统的一些功能有着紧密的联系.例如,脑干中Pre-Bötzinger神经元的混合振荡对于产生规则的呼吸至关重要[30].因此,研究不同混合振荡的单神经元和网络的随机动力学对于动力学和神经科学都具有重要的意义.
本文选用能产生混合振荡的含有自适应电流的改进FitzHugh-Nagumo(FHN)神经元模型,研究了全局电耦合神经元网络的随机动力学.通过计算神经元网络放电ISI的变差系数(coefficient variation,CV)发现:噪声能诱导神经元网络出现反相干共振向相干共振的转迁,而且该转迁现象在较大的耦合强度范围内存在.研究结果丰富了神经元网络的随机动力学,揭示了混合振荡的潜在功能.
1 模型与方法
1.1 FHN神经元模型
FHN神经元模型是一种简单的但具有代表性的神经脉冲模型,可由如下方程描述:
(1)
式中,v和w分别为膜电位和膜恢复变量,a、τ和I为参数,它们都是无量纲的[31].为保证膜电位的快速演化,参数a和τ在本研究中分别被固定为5和60[31].参数I代表刺激电流,其决定了模型的动力学行为.当I< -4.29时,模型有一个稳定的焦点,对应静息态;当I稍大于-4.29时,模型通过Hopf分岔产生一个稳定的极限环,对应周期的放电态.例如,刺激电流I被固定为-4.2时,模型产生周期1峰放电,如图1(a)所示.
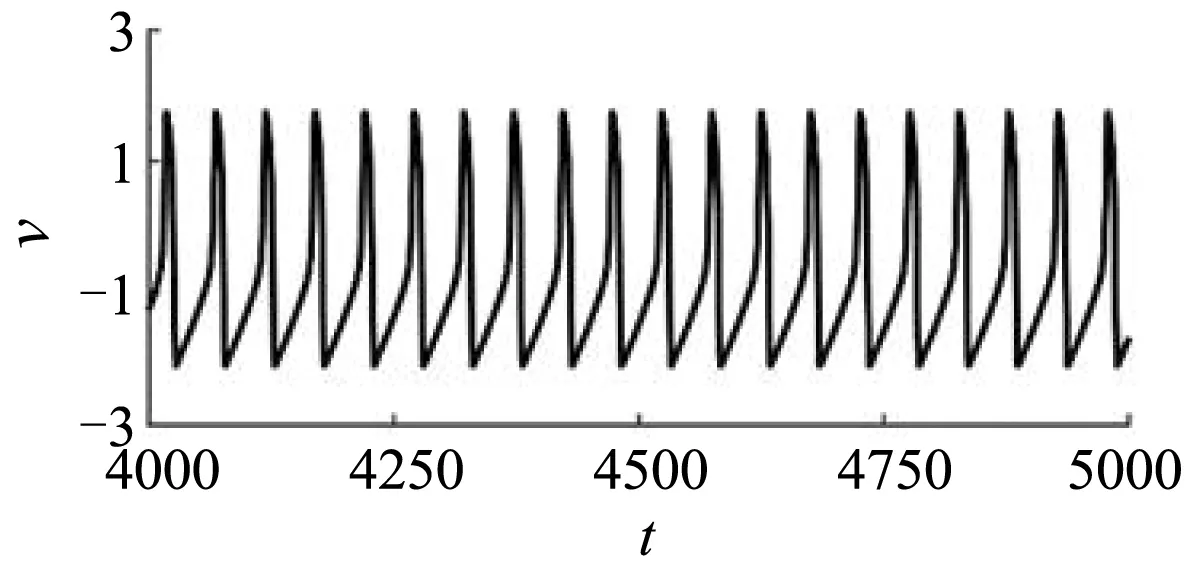
(a)无自适应电流时的周期1峰放电(a)Period-1 spiking with adaptive current absent
1.2 含自适应电流的改进FHN神经元模型
引入自适应电流是使得神经元模型产生混合振荡的常用方法[31],本文在FHN神经元模型中引入自适应电流:
(2)
其中,变量Ia代表自适应电流,参数τa代表自适应电流的衰减时间.自适应电流Ia满足重置条件:当膜电位v正向穿越阈值0时,将自适应电流Ia赋值为δ.在本研究中,衰减时间τa和参数δ分别被固定为150和-0.2[31],模型产生周期5混合振荡,如图1(b)所示.本文以该周期5混合振荡为例进行研究.
1.3 含自适应电流和高斯白噪声的改进FHN神经元网络模型
含自适应电流和高斯白噪声的FHN神经元网络可由如下方程描述:
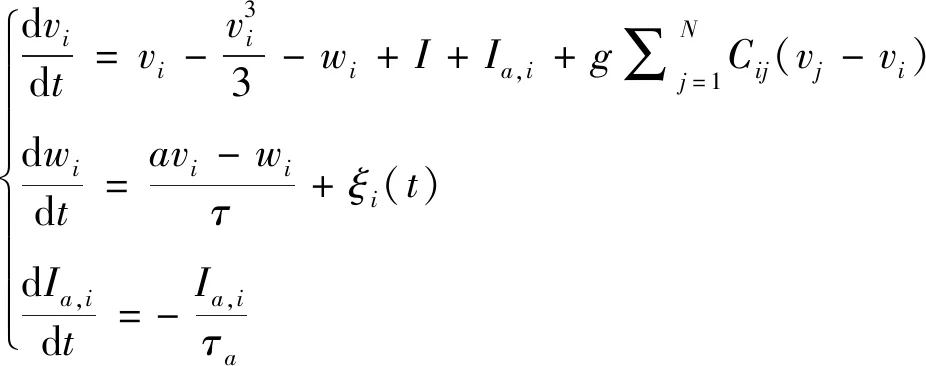
(3)

1.4 方法
本文采用欧拉方法数值求解所有方程,积分步长为0.001.为了刻画神经元网络的相干共振,本文首先计算网络中所有神经元的ISI,然后采用所有神经元的ISI的变差系数CV定量地刻画神经元网络的相干共振.其中,CV定义为所有神经元的ISI的标准差除以所有神经元的ISI的均值[2,3].
此外,本文采用如下指标定量刻画网络中神经元放电的同步[32,33]:
(4)
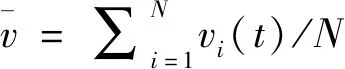
2 结果
2.1 耦合强度对确定性神经元网络放电模式和放电同步的影响
图2刻画了不同耦合强度下所有神经元的放电的时空图(左)和网络中某个神经元的放电图(右).其中,在放电的时空图中,每个黑点都对应于神经元的一次放电.当耦合强度g很小时(例如,当g= 0.00000001或0.000001时),由于初值的随机性神经元放电同步性很弱[图2(a)、(c)],网络中单个神经元的放电为周期5混合振荡[图2(b)、(d)]且与孤立神经元的放电模式类似[图1(b)].当耦合强度g增大为0.0001时,网络中的放电率减小[图2(e)],单个神经元的ISI增大且放电变得不规则[图2(f)].当耦合强度g增大为0.01时,神经元的放电达到完全同步[图2(g)],单个神经元的ISI减小且放电变得规则[图2(h)].当耦合强度g增大为1时,神经元的放电仍为完全同步[图2(i)],单个神经元的ISI减小且放电为规则的峰放电[图2(j)].图2表明耦合强度的增大不仅能增强神经元网络放电的同步,还能改变神经元网络中的放电模式.
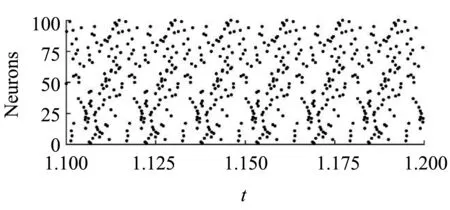
(a)g = 0.00000001时的时空图(a)Space-time plots for g = 0.00000001
为了深入刻画耦合强度对确定性神经元网络放电模式和放电同步的影响,图3刻画了网络中所有神经元的ISI和网络的同步指标S随着耦合强度的g的变化规律.由图3(a)可知,随着耦合强度g的增大,网络中神经元的放电从周期5混合振荡,经过复杂放电,变为周期1峰放电,平均ISI先增大后减小(红色虚线),平均ISI的增大和减小分别是由小幅振荡的个数增加和减少引起的.由图3(b)可知,当耦合强度g较小时(g< 0.0004时),网络的同步性较弱,当g较大时(g≥ 0.0004时),网络的同步为完全同步态.

(a)所有神经元的ISI(黑色点)和ISI的平均(红色虚线)随着耦合强度g的变化规律(a)Dependence of ISIs of all neurons(black dots)and the mean of ISIs(red dashed line)on the coupling strength g
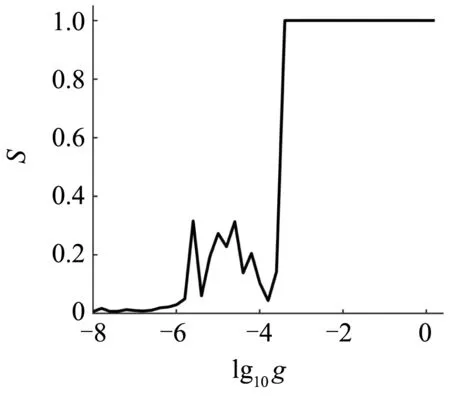
(b)网络的同步指标随着耦合强度g的变化规律(b)Dependence of the synchronization index on the coupling strength g
2.2 弱耦合神经元网络中反相干共振向相干共振的转迁
当网络中神经元间的耦合很弱时(例如,当g=0.0001时),图4刻画了不同噪声强度D下网络中某个神经元的膜电位随时间的演化(左)和所有神经元的ISI统计直方图(右).当D很小时(例如,当D= 0.0000001时),膜电位的振荡为较规则的混合振荡[图4(a)],ISI的分布较集中,在240~330之间[图4(b)].当D增大至0.000032时,一些小的ISI出现[图4(c)],导致ISI的分布变宽,在50-390之间[图4(d)],表明神经元的放电规律性变弱.当D增大至0.0032时,放电的规律性增强[图4(e)],这反映在ISI的分布变窄(在30~130之间)且ISI统计直方图的高度增大[图4(f)].当D进一步增大至D= 3.16时,ISI的分布范围变宽(在20~170之间)且ISI统计直方图的高度降低[图4(h)],表明放电的规律性又变弱[图4(g)].图4提示:随着噪声强度的增大,神经元网络的放电规律性变弱后又变强,可能先出现反相干共振再出现相干共振.
为了定量地验证神经元网络会出现反相干共振向相干共振的转迁,图5(a)描述了当g= 0.0001时变差系数CV随着噪声强度D的变化规律.随着噪声强度D的增大,CV先增大后减小再增大,并在D= 0.000032处达到局部最大值(反相干共振),在D= 0.0032处达到局部最小值(相干共振).图5(a)表明:随着噪声强度D的增大,弱耦合神经元网络会出现反相干共振向相干共振的转迁.图5(b)描述了ISI的标准差(红色圆圈)和均值(蓝色方格)随着噪声强度D的变化规律.变差系数CV的先增大是由ISI的标准差增大而ISI的均值减小引起的;变差系数CV的后减小是由ISI的标准差减小慢于ISI的均值减小引起的;变差系数CV的再增大是由ISI的标准差增大而ISI的均值几乎不变引起的.

(a)D = 0.0000001时的放电图(a)Spike trains for D = 0.0000001

(a)g = 0.0001时CV随着D的变化(a)Dependence of CV on D for g = 0.0001
2.3 中等耦合神经元网络中反相干共振向相干共振的转迁
当耦合中等时(例如,当g= 0.01时),随着噪声强度D的增大,CV先增大后减小再增大,如图5(c)所示,并在D= 0.001处达到局部最大值(反相干共振),在D= 0.01处达到局部最小值(相干共振),还会出现反相干共振向相干共振的转迁.图5(d)描述了ISI的标准差和均值随着噪声强度D的变化规律.与弱耦合强度下不同的是,反相干共振和相干共振出现的噪声强度都增大.
为了更深入地描述该转迁现象,图6刻画了几组关键噪声强度D下,网络中某个神经元的膜电位随时间的演化(左)和所有神经元的ISI统计直方图(右).当D很小时(例如,当D=0.0000001时),膜电位的振荡为规则的混合振荡模式[图6(a)],ISI的分布很集中,在150~160之间[图6(b)].当D增大至0.001时,放电的规律性变弱[图6(c)],ISI的分布变宽,在50-200之间[图6(d)].当D增大至0.01时,放电规律性增强[图6(e)],ISI的分布变窄(在40~100之间)且ISI统计直方图的高度增大[图6(f)].当D进一步增大至1时,放电的规律性又变弱[图6(g)],ISI的分布范围变宽(在20~190之间)且ISI统计直方图的高度降低[图6(h)].当D= 0.001时(反相干共振),放电达到最不规则的状态;当D= 0.01时(相干共振),放电达到最规则的状态.
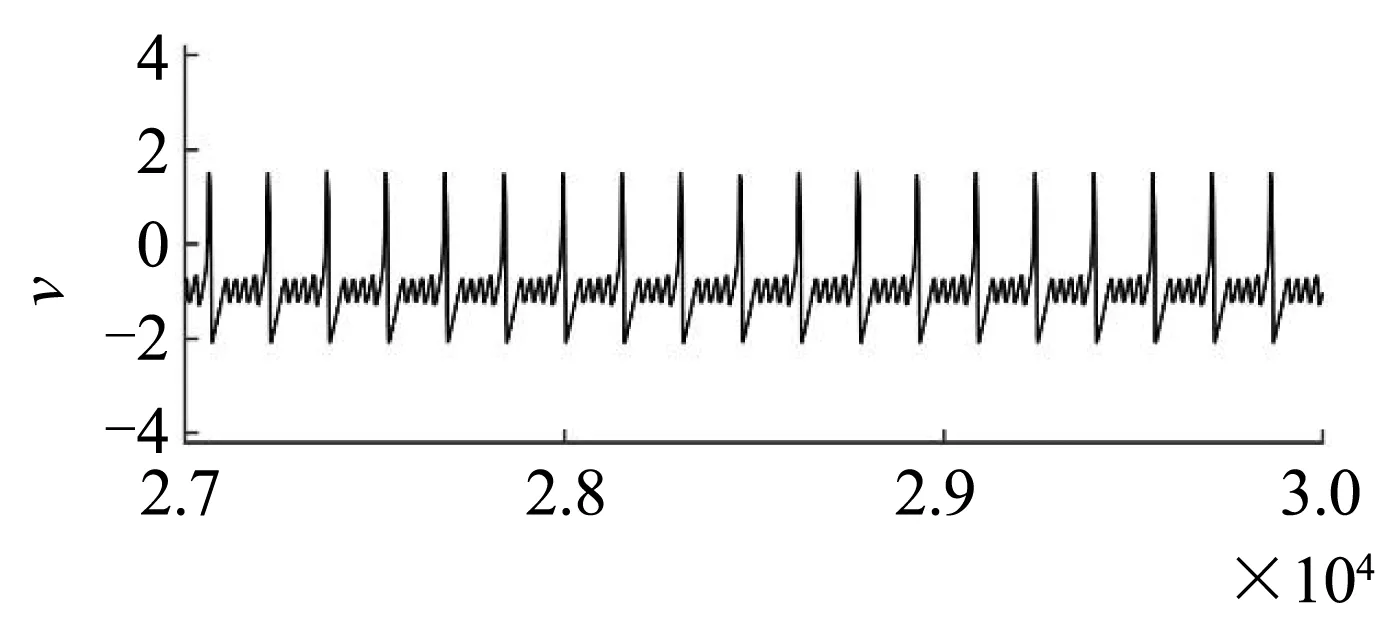
(a)D = 0.0000001时的放电图(a)Spike trains for D = 0.0000001
2.4 强耦合神经元网络中反相干共振向相干共振的转迁
当耦合很强时(例如,当g= 1时),网络还会出现反相干共向相干共振的转迁,如图5(e)所示.变差系数CV在D= 0.001处达到局部最大值(反相干共振),在D= 0.1处达到局部最小值(相干共振).图5(f)描述了ISI的标准差和均值随着噪声强度D的变化规律.变差系数CV的先增大是由ISI的标准差增大快于ISI的均值增大引起的;变差系数CV的后减小是由于ISI的标准差减小快于ISI的均值减小引起的;变差系数CV的再增大是由ISI的标准差增大而均值减小引起的.相比于弱和中耦合强度,ISI的均值变化范围相对较小.
图7刻画了不同噪声强度D下网络中某个神经元的膜电位随时间的演化(左)和所有神经元的ISI统计直方图(右).当D很小时(例如,当D= 0.0000001时),在强耦合的作用下神经元的活动近似为规则的周期1峰放电[图7(a)],这不同于弱和中耦合强度,所有神经元的ISI集中分布在40附近[图7(b)].当D增大至0.001时,神经元的放电规律性减弱[图7(c)],ISI的分布变宽(在40~240之间)且ISI统计直方图的高度降低[图7(d)].当D增大至0.1时,神经元的放电规律性增强[图7(e)],ISI的分布变窄(在30~90之间)且ISI统计直方图的高度增加[图7(f)].当D进一步增大至1时,神经元的放电规律性减弱[图7(g)],ISI的分布变宽(在30~90之间)且ISI统计直方图的高度降低[图7(h)].当D= 0.001时(反相干共振),放电达到最不规则的状态;当D= 0.1时(相干共振),放电达到最规则的状态.

(a)D=0.0000001时的放电图(a)Spike trains for D = 0.0000001
2.5 不同耦合强度下的反相干共振向相干共振的转迁
2.2~2.4节中的结果表明:对于弱、中和强三组不同的耦合强度,神经元网络都会出现反相干共振向相干共振的转迁.图8刻画了六组不同耦合强度g下变差系数CV随着噪声强度D的变化规律.首先,对于六组不同的耦合强度g,变差系数CV都先增大后减小再增大.这表明对于六组不同的耦合强度g,神经元网络都会出现反相干共振向相干共振的转迁.此外,随着耦合强度g的增大,反相干共振和相干共振对应的噪声强度D右移.这说明随着耦合强度的增大,出现反相干共振和相干共振的噪声强度都增大.
图9描绘了变差系数CV随着噪声强度D和耦合强度g的变化规律.其中,深色区域对应于小的CV值,浅色区域对应于大的CV值,如颜色条所示.首先,对于固定的耦合强度g,随着噪声强度D的增大,CV先增大后减小再增大.这表明对于不同的耦合强度g,神经元网络都会出现反相干共振向相干共振的转迁.此外,随着耦合强度g的增大,反相干共振对应的噪声强度(黑色虚线标注)和相干共振对应的噪声强度(白色虚线标注)整体上都有右移的趋势.这说明随着耦合强度的增大,出现反相干共振和相干共振的噪声强度整体上都有增大的趋势.
(a)g = 0.0001

图9 变差系数CV随耦合强度g和噪声强度D的变化规律Fig.9 Dependence of the CV on both the coupling strength g and the noise intensity D
3 结论
混合振荡和相干共振都是神经系统的有代表性和重要意义的动力学行为.本文深入研究了神经元网络混合振荡的随机动力学.结果发现,高斯白噪声能诱导反相干共振向相干共振的转迁,比一般网络只出现相干共振或者只出现反相干共振更为复杂,而且该现象不依赖于神经元间的耦合强度.本结果将最近研究发现的单神经元混合振荡会表现出反相干共振向相干共振的转迁[16]现象扩展到了网络,丰富了神经元网络的复杂随机动力学,揭示了具有混合振荡的神经元网络在信息处理中的潜在功能.
此外,本文还揭示耦合强度对混合振荡有重要的影响,增大耦合强度不仅能增强神经元网络的同步,还能使得混合振荡过渡为周期1峰放电.因此,强耦合强度下的反相干共振向相干共振的转迁是从周期1开始的,不像弱和中等耦合强度下及单神经元[16]中是从混合振荡开始的.因此,进一步研究不同的网络行为在噪声作用下是否会出现相干共振的转迁,是需要研究的随机动力学问题.此外,最近研究表明抑制性自突触能增强神经元及网络的相干共振[24, 34].因此,研究不同的耦合或者调控对混合振荡神经元网络中共振转迁的影响也十分重要.
