荔枝nrDNA ITS序列多态性分析
2022-10-02黄小凤余金昌招胜坚韦阳连
黄小凤,袁 叶,余金昌,招胜坚,韦阳连
(东莞植物园,广东 东莞 523086)
高等植物的核糖体DNA内转录间隔区(nrDNA ITS,包括ITS1、5.8S和ITS2),在核基因组内是高度重复的,这些拷贝位于同一染色体或不同染色体上[1]。nrDNA ITS通常是以致同进化模式进行,在这种模式下,ITS通过不等交换、基因转换和基因扩增等方式使得不同ITS拷贝的序列基本趋于一致或完全一致[2-4],因此,同一物种基因组内ITS序列多样性会倾于降低或消失,即ITS序列在物种内极少出现多态性,其多态性常存在于物种间。ITS序列进化速率快,片段长度适中,能够提供较多的变异信息,因此常用于植物系统发育研究。
随着研究的深入,越来越多的研究表明,ITS序列多样性不仅存在于植物基因组间,在植物基因组内也存在较高的多态性,某些物种内甚至个体内的ITS序列存在致同进化不完全现象,拷贝之间存在明显的差异,一些ITS拷贝可能由于功能退化而变成假基因[5-8]。据报道,裸子植物中,松科(Pinaceae)、买麻藤属(GnetumL.)、苏铁属(CycasL.)等多种植物基因组内均存在ITS序列多态性[8-13];被子植物中,梨属(PyrusL.)和栎属(QuercusL.)、四照花亚属[Cornussubg.Syncar-pea(Nakai)Q.Y.Xiang]等植物基因组内也存在ITS序列多态性,同一物种不同个体间、甚至同一个体的不同克隆间均存在ITS序列多态性[14-16]。这些发现成为应用ITS序列进行进化分析的一个新的挑战,相应的系统进化分析会因同一物种中多样化的序列类型而无法构建出可靠的进化树[17]。
荔枝(Litchi chinensisSonn.)隶属被子植物门无患子科(Sapindaceae)荔枝属(LitchiSonn.),著名岭南佳果,起源于中国和越南,我国海南、广东、广西和云南等地现今仍保存有完好的野生荔枝林。荔枝品种繁多,其中绝大多数是从实生荔枝中选育出来的,从芽变选出的品种极少,通过杂交育种、诱变育种、多倍体育种等现代育种方法获得的品种少有报道[18]。姜帆等[19]对无患子科不同物种的ITS序列进行分析发现,ITS序列在龙眼、车厘子、Beguea apetala等物种的基因组内均存在多态性。笔者在NCBI网站的GenBank数据库中发现,ITS序列在无患子科的欧洲七叶树、Serjania pyramidata、Paullinia stipitata等物种基因组内也存在多态性。荔枝属无患子科,品种繁多,其ITS序列在基因组内是否存在多态性目前尚未知。本文以4个荔枝品种为研究对象,对其ITS片段进行克隆测序,以期检测这些荔枝品种的ITS序列多态性。
1 材料与方法
1.1 试验材料
荔枝依据成熟果实中部果皮上的龟裂片和裂片峰的主要特征可分为3种类型:尖突型、隆起型和平坦型。本研究在3种果皮类型的荔枝种质中各选1个品种作为供试材料。‘妃子笑’是栽培范围最广、适应性最强的荔枝品种,是荔枝种质遗传研究的理想材料,因此也被选为供试材料。4份材料(表1)均采自东莞植物园荔枝种质资源圃。
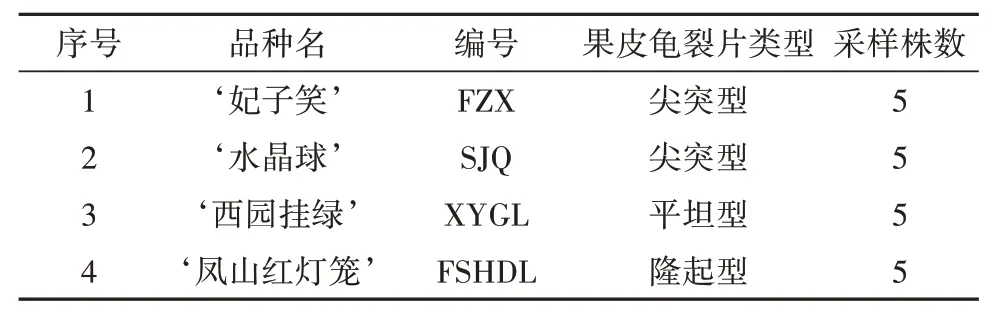
表1 供试材料清单Tab.1 Litchi varieties used in this study
1.2 试验方法
将各品种的叶片样品混合后进行试验。DNA提取、PCR扩增、产物纯化、克隆、测序均委托广州艾基生物技术有限公司完成。所用克隆载体为pGSI,所用测序引物委托测序公司设计(F:5'-CGCCAGGGTTTTCCCAGTCACGAC-3';R:5'-AGCGGATAACAATTTCACA-CAGGA-3')。每个品种选择13个阳性克隆进行测序。
1.3 序列分析及系统发育分析
用DNAMAN9对ITS序列进行比对,根据Gen-Bank中已有的荔枝ITS序列确定ITS1、5.8S、ITS2的边界和序列信息。利用DNAMAN9计算各个ITS序列的长度及GC含量。用DNASP v6分析单倍型核苷酸多态性(π)和核苷酸差异平均值(K)。采用MEGA5.2对遗传距离进行分析,使用ML法构建系统发育树,置信度采用自引导法(Bootstrap analysis)重复检测1 000次。用Arlequin3.5.2.2进行分子方差分析(AMOVA),计算品种间、品种内的变异方差分布。
1.4 二级结构分析
5.8S区域相对于ITS1区域和ITS2区域而言更为保守,因此利用网络在线版Mfold对所有样本序列中5.8S二级结构最小自由能(△Gat 37℃)进行测算,并分析5.8S中保守基序的碱基变化情况;同时利用在线版Mfold对ITS序列进行二级结构分析。
2 结果与分析
2.1 序列特征及遗传距离分析
将所获得的所有序列信息在NCBI中进行BLAST搜索,其中克隆FXZ-5、FZX-9、SJQ-1、SJQ-5、SJQ-8、XYGL-3、XYGL-10、FSHDL-3、FSHDL-4、FSHDL-8、FSHDL-9、FSHDL-10的序列搜索结果为真菌ITS序列,这是因为本研究所用ITS扩增引物为通用引物(ITS4:5'-TCCTCCGCTTATTGATATGC-3';ITS5:5'-GGAAGTAA-AAGTCGTAACAAGG-3'),在实验中将叶片内生真菌的ITS序列也扩增出来,故而克隆测序结果中会出现真菌的ITS序列;其余克隆序列均与荔枝的ITS序列(登录号MW070204.1)高度同源,因此选择登录号MW070204.1的ITS序列作为荔枝ITS对照序列。
各供试材料除去真菌的克隆序列后,‘妃子笑’、‘水晶球’、‘西园挂绿’和‘凤山红灯笼’共获得40条ITS序列(尚未上传到GenBank上)。序列比对结果显示:40个克隆所获序列均与ITS对照序列不同;同一荔枝品种的不同克隆之间,‘西园挂绿’的2个克隆XYGL-8和XYGL-9的序列完全一致,‘凤山红灯笼’的4个克隆(FSHDL-1、5、6、13)的序列完全一致;不同品种的克隆之间,除了‘西园挂绿’的克隆XYGL-11和‘凤山红灯笼’的4个克隆(FSHDL-1、5、6、13)的序列完全一致外,其余克隆的序列各不相同;FZX-6在ITS1区有一段27 bp的插入片段,使其序列全长达642 bp。
序列分析结果显示,40个ITS拷贝的序列全长为601-642 bp,GC含量为57.17%~67.74%。其中,ITS1长度为226-265 bp,GC含量为57.98%~70.19%;5.8S长度均为164 bp,GC含量均为50.61%~55.49%,最小自由能为-53~-42.9 kcal/mol;ITS2长度为207-222 bp,GC含量为61.32%~71.23%。
ITS序列核苷酸多态性分析结果(表2)显示,‘妃子笑’的ITS序列核苷酸多态性π为0.028 12,平均核苷酸差异为16.564,11条ITS克隆序列中含有变异位点39个。‘水晶 球’的ITS序 列 核苷 酸多 态 性π为0.065 37,平均核苷酸差异为39.289,10条ITS克隆序列中含有变异位点155个。‘西园挂绿’的ITS序列核苷酸多态性π为0.013 32,平均核苷酸差异为8.073,11条ITS克隆序列中含有变异位点23个。‘凤山红灯笼’的ITS序列核苷酸多态性π为0.033 04,平均核苷酸差异为19.893,8条ITS克隆序列中含有变异位点71个。ITS序列在荔枝品种内的核苷酸多态性排序为‘水晶球’>‘凤山红灯笼’>‘妃子笑’>‘西园挂绿’。
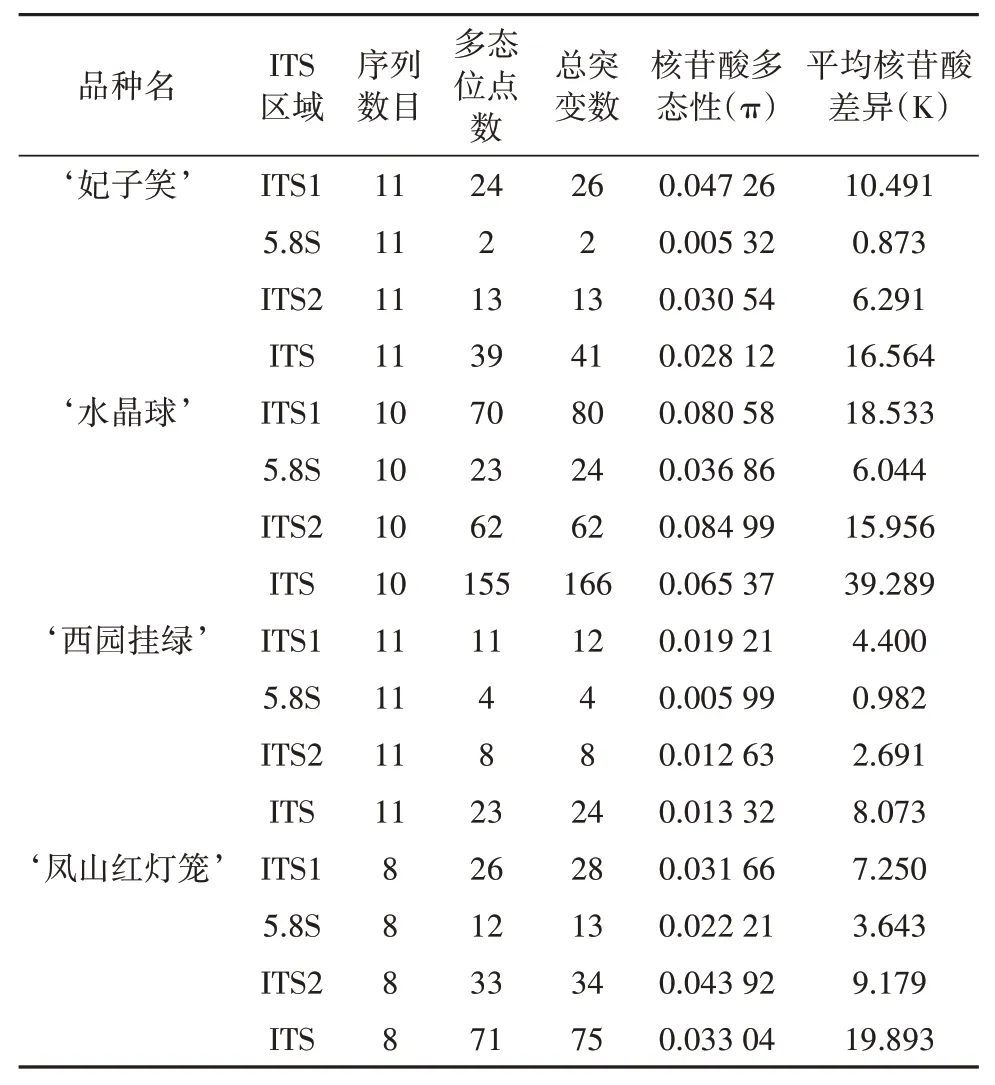
表2 不同荔枝品种的ITS序列核苷酸多态性Tab.2 Nucleotide diversity of the ITS region in different litchi varieties
基于ITS克隆序列对荔枝进行品种内和品种间遗传距离分析,结果(表3)表明荔枝ITS序列品种内平均遗离为0.013 5~0.072 7,其排序与其品种内核苷酸多态性排序一致,均为‘水晶球’>‘凤山红灯笼’>‘妃子笑’>‘西园挂绿’,其中‘水晶球’品种内平均遗传距离(0.072 7)大于品种间平均遗传距离(0.043 4)。
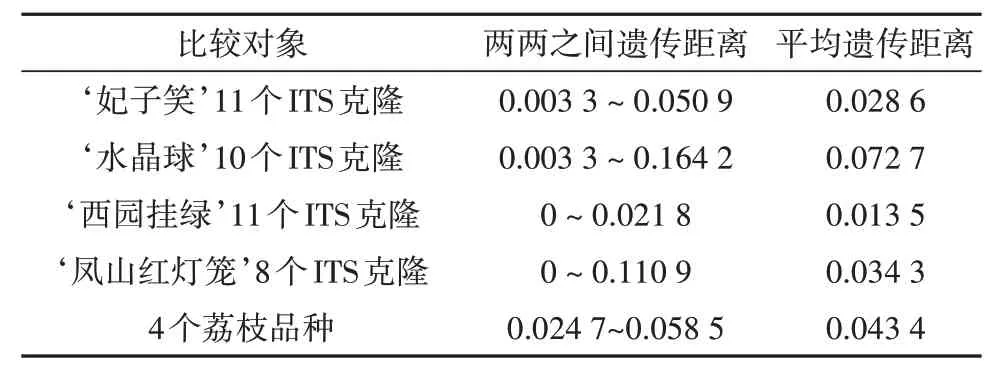
表3 荔枝品种内、品种间遗传距离Tab.3 Genetic distance within or between litchi varieties
为进一步分析荔枝品种间、品种内ITS序列的差异,运用Arlequin软件对其进行AMOVA分析,结果(表4)显示荔枝品种间的遗传变异百分比为11.74%,而品种内的遗传变异百分比达88.26%,表明其品种内遗传多样性水平高于品种间。
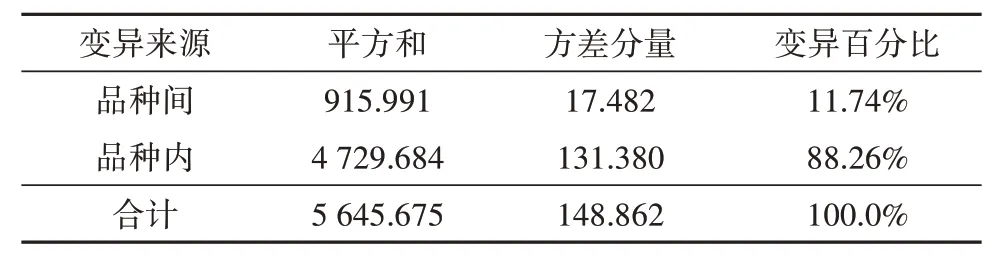
表4 4个荔枝品种遗传变异的分子方差分析(AMOVA)Tab.4 AMOVA analysis of genetic variation of 4 litchi varieties
2.2 ITS假基因
nrDNA ITS基因家族有成千上万的拷贝,在进化过程中一些拷贝可能会功能退化而变成假基因,假基因通常具有明显的长度变异、较低的GC含量、较高的最小自由能、5.8S区变异增加等特征[15,17,20]。
荔枝为种子植物,其ITS序列中的5.8S区域中应包含3个种子植物特有的保守基序(motif 1∶CGATGAAGAACGyAGC、motif 2∶GAATTGCAGAATCC、motif 3∶TTTGAAyGCA),其中保守基序2最为保守,5.8S基序的保守性分析是假基因判定的主要标准[20-23],通过对荔枝的40个克隆序列进行保守基序分析,发现所有序列的保守基序2均无变化,部分序列的保守基序1和保守基序3有不同程度的变异(表5):保守基序1中,SJQ-3在第50个碱基位点存在核苷酸的替换,SJQ-4在49、52、54这3个位点中存在核苷酸的替换,SJQ-6在第54个碱基位点有替换,FSHDL-7在第49个碱基位点存在核苷酸的替换;保守基序3中,FSHDL-11的第112位碱基发生替换;保守基序1和3中的替换均为C→T、G→A的转变。SJQ-3、SJQ-4、SJQ-6、FSHDL-7、FSHDL-11等5条序列在5.8S的3个保守基序中至少有一个基序存在碱基变异,可判定为假基因。
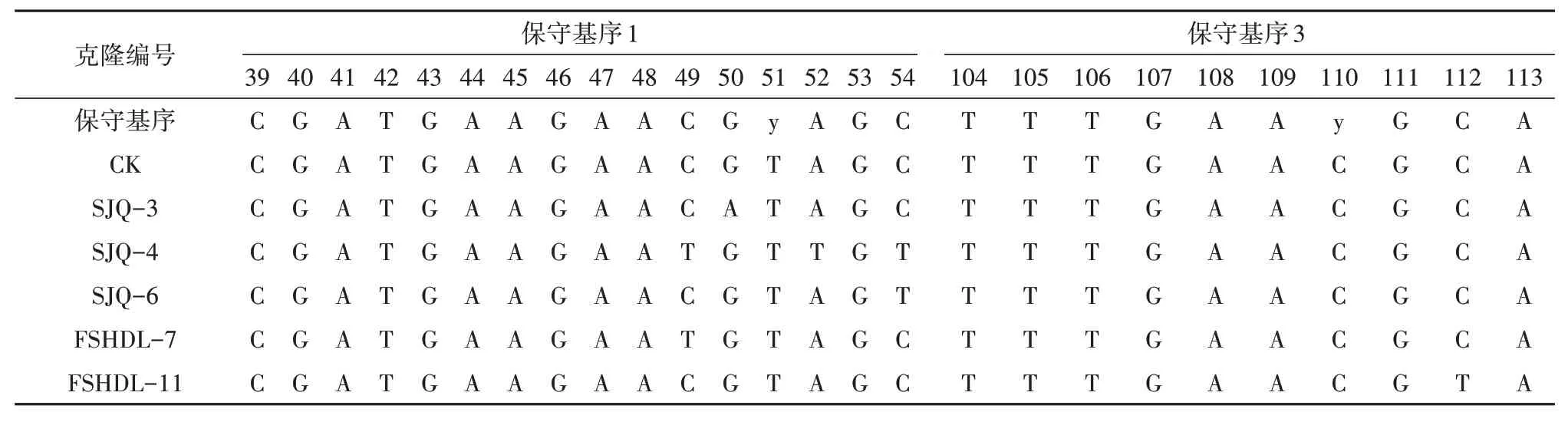
表5 不同ITS序列拷贝5.8S中2个保守基序的核苷酸变化Tab.5 Nucleotide changes in two motifs of 5.8S of different ITS copies
对所获各序列的长度、GC含量、5.8二级结构最小自由进行排序分析,发现5.8S没有变异的序列中:FZX-6的序列长度明显长于对照,也长于4个假基因(SJQ-3、SJQ-4、SJQ-6和FSHDL-7);SJQ-9的GC含量低于5个假基因;SJQ-2、XYGL-6的二级结构最小自由能大于等于3个假基因(SJQ-4、FSHDL-7和FSHDL-11)。假基因的判断存在多种标准,从序列具有明显的长度变异、GC含量较低、最小自由能较高等标准来分析,FZX-6、SJQ-2、SJQ-9、XYGL-6也可判定为假基因。
2.3 系统发育分析
利用ML法构建系统发育树,分析同一品种的不同克隆以及不同品种克隆之间亲缘关系的远近。由图1可知,同一荔枝品种的不同克隆没有按照品种聚在一起。40个ITS克隆大致可分为2组:A组含32个ITS克隆,其中除了XYGL-6是假基因外,其余全部为功能基因;B组含8个ITS克隆,全部为假基因。从ML树整体可以看出,4个荔枝品种的31个ITS功能基因首先聚在一起,显示其亲缘关系较近;4个荔枝品种的9个ITS假基因中,除了XYGL-6穿插到ITS功能基因聚类的A组外,其余8个假基因因序列变异较大,分别聚在B组的4个分支,表明其亲缘关系较远。
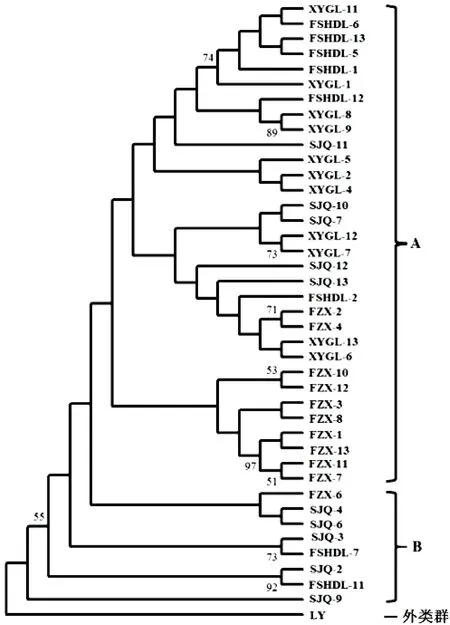
图1 基于ITS序列构建的ML系统发育树Fig.1 ML phylogenetic tree based on ITS sequence
2.4 RNA二级结构分析
在4个荔枝品种的ITS功能基因中,分别选择遗传距离最远的两个克隆(‘妃子笑’中的FZX-2、FZX-12;‘水晶球’中的SJQ-7、SJQ-13;‘西园挂绿’中的XYGL-5、XYGL-13;‘凤山红灯笼’中的FSHDL-2、FSHDL-6),采用Mfold在线版绘制其rRNA二级结构,如图2所示,各品种中遗传距离最远的两个克隆因ITS片段一级结构的长度变异以及核苷酸的转换、颠换等变化,导致二级结构构象变异较大,茎环结构以及内部环的结构均不相同,二级结构不稳定且不保守,说明ITS序列在样本内出现了较为明显的变异,发生了明显的遗传分化。
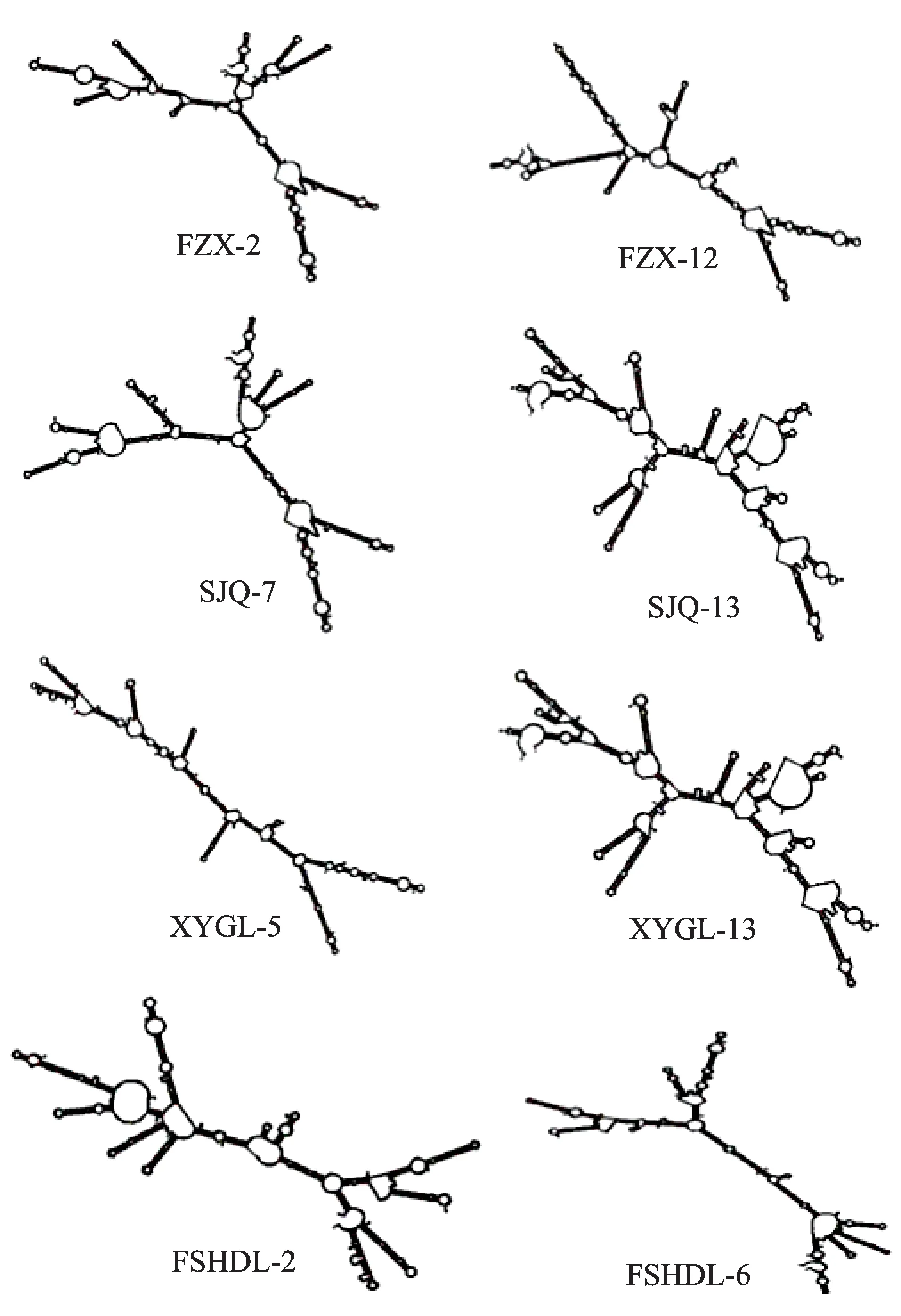
图2 基于ITS序列构建的rRNA二级结构Fig.2 rRNA secondary structure based on ITS sequence
3 讨论与结论
3.1 荔枝nrDNA ITS多态性
近年来,nrDNA ITS多态性在越来越多的植物中被发现,如苏铁属[8]、四照花亚属[16]、龙眼属[19]、云南山茶[24]、枸杞属[25]等。多倍化、孤雌生殖、杂交以及基因家族在染色体上的结构方面因素等都可能导致致同进化不完全,产生基因组内ITS多态性[8]。本研究发现,荔枝nrDNA ITS在基因组内存在多态性:4个荔枝品种的40个ITS克隆获得了35条不同的序列,ITS序列全长变异范围为601-642 bp,ITS1长度变异范围为226-265 bp,ITS2长度变异范围为207-222 bp,其中5个克隆在5.8S区发生了变异。荔枝nrDNA ITS序列多态性不仅出现在品种间,还出现在品种内,且品种内遗传变异高于品种间遗传变异,因此,ITS序列不宜用于进行荔枝的系统进化研究。
荔枝nrDNA ITS在品种间和品种内的多态性表明,其ITS逃离了致同进化,其原因可能与荔枝品种的选育有关:迄今为止,我国大多数荔枝主栽品种都是前人有意或无意从实生资源中选择出来的。荔枝实生选种是指从荔枝实生繁殖所产生的自然变异中,选择出无性系品种的方法,供选群体主要是由播种自然杂交(即自然授粉)种子所产生的实生树。我国近些年选育的‘凤山红灯笼’、‘御金球’、‘燎原’、‘冰荔’、‘井岗红糯’等30个新品种均来自实生选种,父、母本都不清楚[26]。荔枝为异花授粉的果树,本身的遗传基础十分复杂,基因型为杂合型。荔枝品种间的自然杂交使遗传物质从一个品种进入另一个品种,易引起遗传多样性的增加,产生多样复杂的自然变异[8]。‘凤山红灯笼’是通过实生选种选育出来的荔枝品种,在本研究构建的ML系统发育树中,它的8个ITS克隆没有聚在一起,而是分散在不同的分支:5个克隆与‘水晶球’、‘西园挂绿’的部分克隆聚在A组上部的一个小分支,1个克隆与‘妃子笑’、‘水晶球’、‘西园挂绿’的部分克隆聚在A组下部的1个小分支,另有2个克隆聚在B组的2个小分支上,表明不同ITS序列拷贝在‘凤山红灯笼’基因组内发生了不同程度的变异,导致序列拷贝之间的遗传距离各不相同,分别聚在系统树的不同小分支上。‘凤山红灯笼’功能基因中遗传距离最远的2个克隆(FSHDL-2、FSHDL-6)的rRNA二级结构的发夹环、内环、凸环的数量和位置及单链结构的位置等均有差异,这些变化均与其ITS序列的长度变异以及核苷酸的转换、颠换等变化有关。同样地,类似的ITS序列拷贝变异情况也发生在‘妃子笑’、‘水晶球’和‘西园挂绿’的基因组内,导致同一品种的不同克隆分散在进化树的不同小分支上,不同克隆的rRNA二级结构产生变异。荔枝基因组内的ITS序列多态性是其品种间自然杂交引起种质遗传多样性增加的体现。
3.2 荔枝ITS假基因
本文在所研究的4个荔枝品种中共发现9个ITS假基因,其中‘水晶球’中的假基因数量最多(5个),‘妃子笑’和‘西园挂绿’中的假基因数量最少(1个),表明假基因在荔枝品种间的产生频率不同;‘水晶球’的5个ITS假基因拷贝分散在不同的分支上,‘凤山红灯笼’的2个ITS假基因拷贝也分散在2个分支上,表明ITS假基因在荔枝品种内也存在多态性。
ML系统发育树显示,绝大部分假基因拷贝在基因树上能与功能基因(A组)明显分开,独自形成分支聚在B组,这与梨属[14]、四照花亚属[16]、乳突球属[27]等的报道相似,说明这些假基因起源较早,且难以与功能序列之间进行遗传信息的交换。而假基因拷贝XYGL-6与功能基因聚在A组,显示出较近的亲缘关系,其原因可能是XYGL-6正从功能基因向假基因转化,因此它与功能基因的亲缘关系较近。
