亚胺还原酶的改造及其催化合成1-苯基-1,2,3,4-四氢异喹啉的性能
2022-08-29曹文斌陈飞飞郑高伟许建和
曹文斌, 李 昊, 陈飞飞, 潘 江, 郑高伟, 许建和
(华东理工大学生物工程学院,省部共建上海生物制造产业协同创新中心,上海 200237)
四氢异喹啉(THIQs)作为具有生物活性的分子骨架在天然产物与药物中存在非常普遍[1-2],常作为受体拮抗剂[3]、受体激动剂[4]、酶抑制剂[5-6]等。其中1-苯基-1,2,3,4-四氢异喹啉(1-Ph-THIQ)由于对神经系统具有独特的抑制作用引起制药领域的广泛关注[7],而(S)-1-苯基-1,2,3,4-四氢异喹啉((S)-1-Ph-THIQ)是膀胱过度活动症治疗药物索利那新(Solifenacin)的关键结构砌块[8]。近年来,亚胺还原酶(IRED)催化环形亚胺不对称氢化的反应路线具有反应条件温和、立体选择性高的优点,被越来越多地用于不同结构手性胺的不对称合成[9-10]。然而,由于亚胺底物1-苯基-四氢异喹啉(1-Ph-DHIQ)中苯基的巨大位阻和苯基对邻近官能团的共轭效应,使其酶法合成具有较大挑战性。
迄今为止,对1-Ph-DHIQ 具有S对映体选择性的亚胺还原酶报道仅有2 例[11-12],一是亚胺还原酶SnIR[11],比活力只有13 mU/mg,对映体选择性(ee值)为51%;而改造所得的突变体T123I/G228T的比活力增加到56 mU/mg,ee值达91% ;二是武汉大学瞿旭东课题组报道的亚胺还原酶IR45[12],其对映体选择性具有较高ee值(99%),且对底物1-Ph-DHIQ 具有较好的催化性能,但存在明显的底物抑制。
为获得高效催化合成(S)-1-Ph-THIQ 的亚胺还原酶,本文首先对前期筛选获得的亚胺还原酶(IRED)进行反应性能比较,然后对性能较好的候选酶AdIR1 进行分子改造,并对所得的酶突变体进行动力学与热稳定性表征,最后尝试利用优质突变酶进行(S)-1-Ph-THIQ 克级制备实验。
1 实验部分
1.1 原料和试剂
琼脂糖凝胶DNA 回收试剂盒,质粒小量提取试剂盒和PCR 产物回收试剂盒购自上海艾德莱生物有限公司;DNA marker、蛋白质marker、2×Taq PCR Mix购自上海康为有限公司;引物由上海生工生物工程技术有限公司代合成;Ni 柱吸附亲和层析柱购自GE Healthcare 公司;蛋白胨和酵母膏购自Oxoid 公司;限制性内切酶购自New England Biolabs有限公司;PrimeSTAR 高保真酶和T4 DNA 连接酶购自Takara有限公司;其余化学试剂均为国产分析纯。
PCR 仪、电泳仪、凝胶成像仪、紫外切胶仪、蛋白电泳仪、酶标仪,美国Bio-Rad 公司;超微分光光度计,Nanodrop 2000c 型,美国Thermo Fisher 公司;全自动高压灭菌锅,日本TOMY 公司;高速离心机,日本HITACHI 公司;紫外-可见分光光度计,日本SHIMADZU 公司;超净工作台,苏州净化设备有限公司;恒温培养箱,上海一恒科技有限公司;恒温震荡摇床、深孔板摇床,上海智诚科技有限公司;超声破碎仪,宁波新芝生物科技有限公司。
1.2 目的基因克隆
本文使用表1 中所示引物通过PCR 分别从菌株Saccharothrix espanaensisDSM 44229 和Amycolatopsis decaplaninaDSM 44594 的基因组中扩增获得亚胺还原酶目的基因AdIR1和SeIR2。
PCR 体系(20 μL):基因组模板1 μL (0.2 μg),上下游引物分别加入1 μL(0.5 μmol/L),2 × Taq Mix 10 μL,和ddH2O 7 μL。PCR 扩增步骤:(1) 95 ℃,预变性3 min;(2) 95 ℃,变性10 s;(3) 55 ℃,退火5 s;(4) 72 ℃,延伸1 min,然后步骤(2)~(4)循环30 次;(5) 72 ℃延伸,延伸10 min,冷却至4 ℃保存。
使用φ=1%的琼脂糖凝胶对PCR 产物进行电泳分析,采用琼脂糖凝胶DNA 回收试剂盒回收目的基因片段。目的基因和pET-28a(+)质粒经限制性内切酶NdeI 和EcoR I 双酶切、T4 DNA 连接酶连接构建重组质粒,并转化至Escherichia. coli(E.coli) BL21 (DE3)感受态细胞。
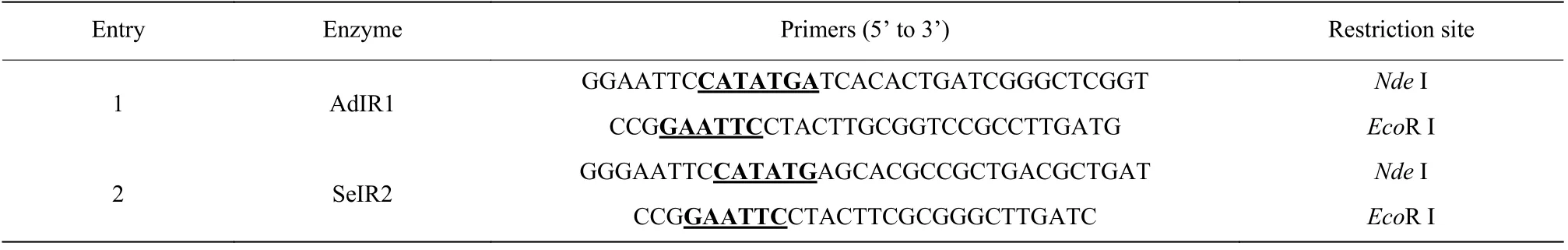
表1 目的基因克隆所用的引物Table 1 Primers used in target gene cloning
1.3 重组菌诱导与表达
从重组菌E. coliBL21 (DE3)甘油保藏管中取样10 μL 接种至4 mL 的Lysogeny Broth (LB)培养基中(含50 μg/mL卡那霉素),于37 ℃、200 r/min 条件下培养过夜。从菌液试管中转接1 mL 菌液至含100 mL LB 培养基(含50 μg/mL 卡那霉素)的摇瓶中放大培养,当摇床(37 ℃,220 r/min)培养至吸光度OD 值(OD600)为0.6~0.8 时,加入异丙基-β-D-硫代半乳糖苷(IPTG,终浓度为0.2 mmol/L)于16 ℃、200 r/min条件下诱导表达24 h。最后在4 ℃、7 000g下离心收集菌体,对收集的菌体进行超声破碎获得细胞破碎液。
1.4 1-苯基-3,4-二氢异喹啉的不对称还原反应
1 mL 反应体系包含1 mmol/L 1-苯基-3,4-二氢异喹啉、0.1 mmol/L 还原型辅酶Ⅱ(NADPH)、5 mmol/L葡萄糖、1 mg/mL葡萄糖脱氢酶BmGDH 冻干酶粉[13]、100 mmol/L KPB 缓冲液(pH 7.0)和200 μL 新鲜细胞破碎上清(上清液按1 g 细胞配比10 mL 缓冲液制备)。不对称还原反应如图1 所示,其中烟酰胺腺嘌呤二核苷酸(NADP+)是还原型烟酰胺腺嘌呤二核苷酸磷酸(NADPH)的氧化形式,GDH 为葡萄糖脱氢酶。
反应条件为30 ℃、200 r/min,每组反应至少重复2 次。采用2 mol/L NaOH 淬灭反应并调节pH 至13,随后立即用乙酸乙酯进行萃取,并用高效液相色谱分析反应转化率和产物ee值。1-苯基-1,2,3,4-四氢异喹啉经柱前乙酰化后的高效液相色谱法(HPLC)分析,分析条件为:OD-H 柱(250 mm × 4.6 mm, 5 μm),正己烷和异丙醇体积比为95∶5,流速0.8 mL/min,波长220 nm,温度30 ℃。(R)-产物出峰时间为20~21 min;(S)-产物出峰时间为22~23 min。
1.5 亚胺还原酶的序列与结构比对
通过CLUSTAL X 程序进行序列比对。通过Swiss-model 在线同源建模[14](以PDB: 4OQY 的蛋白结构作为模板)分别构建得到SnIR 与AdIR1 (与4OQY 的序列一致性分别为43%和46%)的结构模型,再利用Pymol 软件对SnIR 与AdIR1 进行空间结构比对。

图1 亚胺还原酶催化的不对称还原反应Fig. 1 Asymmetric reduction reaction catalyzed by IRED
1.6 AdIR1 酶突变库的构建与初筛
以母本AdIR1 重组pET-28a(+)质粒为模板,使用PrimeSTAR 高保真聚合酶和表2 所示的目的基因定点突变所用PCR 引物进行PCR 构建单点饱和突变库。PCR 产物经DpnI 酶消化(37 ℃、2 h)后转化至E. coliBL21 感受态细胞,并涂布LB 平板(含50 μg/mL 卡那霉素),于37 ℃培养箱过夜培养。
突变体库筛选步骤如下:
(1) 用牙签将平板上的突变体转化子转移至一级板(单孔有300 μL LB 培养基,含50 μg/mL 卡那霉素)中,于37 ℃、220 r/min 下振荡过夜培养,每块板中含有3 个母本和34 个转化子。
(2) 取50 μL 一级种子液转接至二级板(单孔有600 μL LB 培养基,含50 μg/mL 卡那霉素)中培养,37 ℃下培养3 h,加入异丙基-β-D-硫代半乳糖苷(IPTG,终浓度0.2 mmol/L),并于16 ℃下继续培养24 h 诱导突变体表达。
(3) 在3 500 r/min 条件下离心10 min 以收集菌体。
(4) 每孔加入200 μL 裂解液(100 mmol/L KPB 缓冲液,pH 7.0,含750 mg/L 溶菌酶和10 mg/L DNase酶),振荡使细胞悬浮均匀,然后于37 ℃、200 r/min条件下保温2 h 破碎细胞。
(5) 在4 ℃条件下,3 500 r/min 离心30 min 后取上清用酶标仪在酶标板中测定活力。
筛选反应条件(200 μL):筛选反应液150 μL (含NADPH 0.27 mmol/L,底物1-Ph-DHIQ 0.13 mmol/L,100 mmol/L KPB 缓 冲 液,pH 7.0),离 心 破 碎 液 上清50 μL。
1.7 AdIR1 突变体的复筛
采用1.3 节所述方法对目标突变体进行发酵培养,使用镍离子亲和吸附柱(His Trap Ni-NTA FF column, 5 mL)对发酵表达的含有His-tag 的重组酶蛋白进行纯化[13],并对纯酶的比活力进行测定。
1.8 纯酶测活与蛋白浓度测定
亚胺还原酶的标准酶活测定体系(1 mL):底物为1-苯基-3,4-二氢异喹啉的二甲基亚砜(DMSO)溶液(10 μL,底物终浓度为1 mmol/L)、NADPH (10 μL,终浓度为0.1 mmol/L),一定浓度的酶液(10 μL)及970 μL KPB 缓冲液(100 mmol/L,pH 7.0)。在30 ℃条件下检测340 nm 处吸光度的变化。
酶活力(U)单位的定义为:在上述条件下,每分钟氧化1 μmol 辅酶所需的酶量,酶活计算公式如下所示:
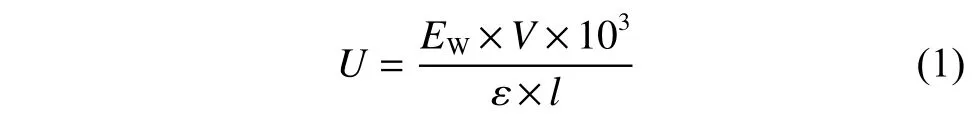
其中:EW代表340 nm 处吸光度在1 min 内的变化值;V代表反应液体系的体积量(mL);ε代表消光系数(L/(mol·cm));l代表光程距离(cm)。
蛋白质含量测定方法:使用Nanodrop 2000c 超微量分光光度计对纯酶蛋白含量进行测定,所得测量值除以蛋白的消光系数即得实际蛋白浓度。
1.9 突变体的酶学性质表征
最适催化pH 研究条件:测定亚胺还原酶在pH 3.0~11.0 范围内不同缓冲液的酶活性,其中pH 3.0~6.0选用柠檬酸-柠檬酸钠缓冲液(100 mmol/L),pH 6.0~8.0 选用KPB 缓冲液(100 mmol/L),pH 8.0~9.0 选用Tris-HCl 缓冲液(100 mmol/L),pH 9.0~11.0 选用甘氨酸-氢氧化钠缓冲液(100 mmol/L)。
热稳定性研究条件:对酶催化活力的半衰期(t1/2)考察,预先用KPB 缓冲液(100 mmol/L, pH 7.0)将AdIR1 突变体酶溶液稀释至1 mg/mL 后,置于不同的温度(30、40、50 ℃)下保温,间隔取样,按标准测活条件测定酶的残余活力。以初始活力为基准,将测定得到酶的残余活力换算为相对活力,相对活力数据通过Excel 线性处理并拟合得到t1/2。

表2 目的基因定点突变所用PCR 引物Table 2 PCR primers used for site-directed mutations of target gene
催化动力学研究条件:测定不同浓度底物(1-甲基-3,4-二氢异喹啉)、不同辅酶(NADPH)浓度下酶的比活力数据,并采用Origin 9.0 软件根据米氏方程模型拟合获得酶的动力学参数,分为以下3 种:
(1)无底物抑制条件下的催化模型
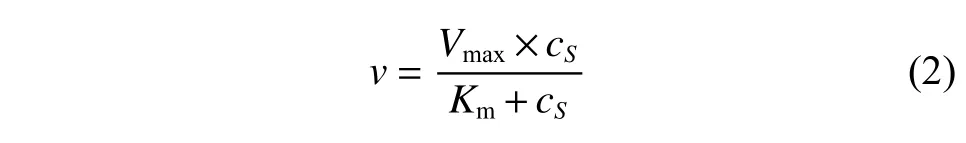
其中:v为酶的比活(μmol/(min·mg));cS为底物或者辅酶浓度(mmol/L);Vmax为催化最大反应速度(μmol/(min·mg)),是底物饱和时的反应速度;Km为米氏常数(mmol/L),是酶达到1/2Vmax时的底物浓度。
(2)底物抑制条件下的催化模型
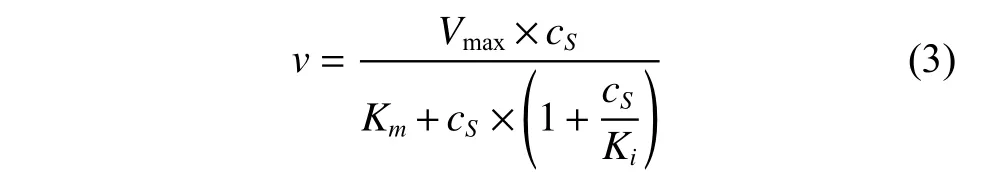
其中:Ki为抑制常数(mmol/L),反映出抑制剂与蛋白质的结合紧密程度。
(3)催化常数kcat计算方程
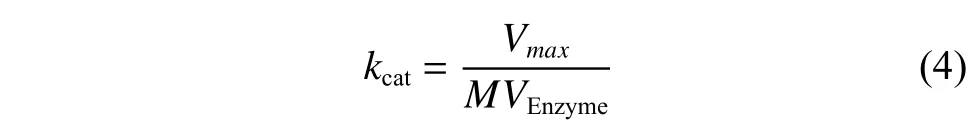
其中:kcat为催化速率(s-1),是一个酶(或者一个活性部位)在底物饱和状态下反应快慢的参数;MVEnzyme为单位质量酶的分子数量(mmol/mg)。
1.10 1-苯基-1,2,3,4-四氢异喹啉的克级制备
1.10.1 最适反应温度考察 1 mL 反应体系包含5 mmol/L 1-苯基-3,4-二氢异喹啉、0.1 mmol/L NADPH、25 mmol/L 葡萄糖、1 mg/mL BmGDH 冻干酶粉、5 mg/mL 亚胺还原酶冻干酶粉、100 mmol/L KPB 缓冲液(pH 7.0)和 φ =1%的DMSO。反应条件为转速200 r/min,温度分别为30、40 ℃。
1.10.2 最适助溶剂体积分数考察 10 mL 反应体系包含5 mmol/L 1-苯基-3,4-二氢异喹啉、0.1 mmol/L NADPH、25 mmol/L葡萄糖、1 mg/mL BmGDH 冻干酶粉、5 mg/mL 亚胺还原酶冻干酶粉、100 mmol/L KPB 缓冲液(pH 7.0),反应条件为转速200 r/min,40 ℃,助溶剂体积分数分别为1%、2%、5%及10% 。
1.10.3 克级制备反应 在以上筛选得到的最适温度以及最适助溶剂浓度条件下,以8 g/L 的突变体冻干酶粉为催化剂,其余反应条件参照1.4 节按比例扩大至2 L 反应体系下进行1-苯基-1,2,3,4-四氢异喹啉的克级制备反应。反应过程定期取样检测反应进程。反应结束后进行淬灭、萃取通过硅胶柱层析法(流动相:乙酸乙酯和石油醚体积比为2∶1)对目标产物进行纯化。
2 结果与讨论
2.1 1-苯基-3,4-二氢异喹啉的不对称酶促还原反应
通过以上反应条件,本文探索了AdIR1 和SeIR2分别对1-苯基-3,4-二氢异喹啉的不对称还原反应效果,结果如表3 所示。可以看出,AdIR1 是一个具有较优催化效率的亚胺还原酶,经过24 h 的反应,实现了98%的转化率(C),合成S构型产物,ee值达98%,结果明显优于SeIR2。因此,后续以反应效果更优质的AdIR1 作为分子改造的起始酶。

表3 亚胺还原酶不对称还原1-苯基-3,4-二氢异喹啉的反应Table 3 Asymmetric reduction of 1-phenyl-3,4-dihydroisoquinoline by IRED
2.2 AdIR1 与SnIR 的序列与结构比对
在前期研究中发现,T123I、F178L、G228A 3 个位点的突变会显著影响SnIR 酶催化1-芳基二氢异喹啉不对称催化产物的ee值,同时也导致了活力的下降。因此,文中将SnIR 与AdIR1 进行序列及结构比对,以期找到与SnIR 的T123、F178 和G228 位点相对应的AdIR1 关键残基,结果如表4 以及图2 所示。

表4 亚胺还原酶的关键位点比对结果Table 4 Alignment results of key residues of the imine reductases
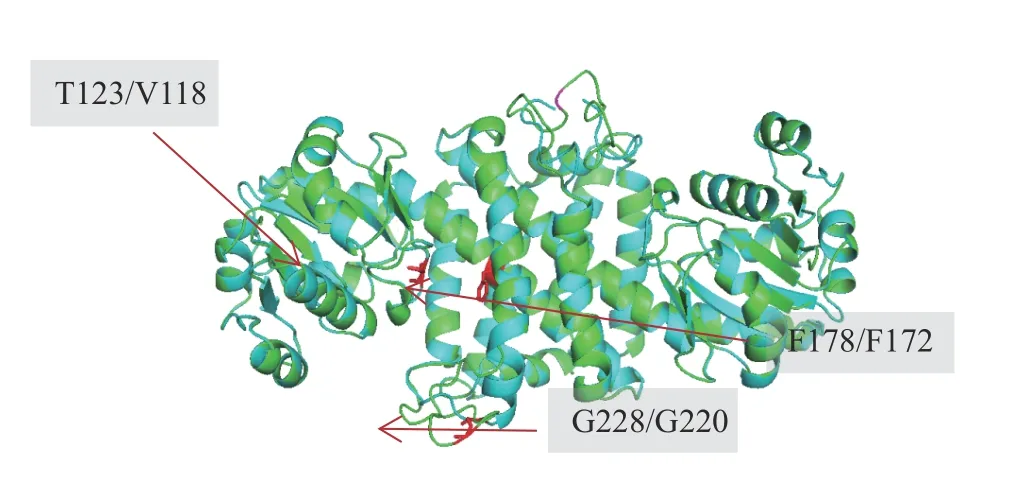
图2 SnIR 与AdIR1 的结构比对结果Fig. 2 Results for structural alignment of AdIR1 with SnIR
以上比对结果表明,对于V118 与F172 位点,AdIR1 与SnIR 通过序列比对的定位结果与空间结构比对的结果一致。由于这两个位点都位于序列保守区域,故可信度比较高。而另一方面对于G228 位点,一级序列的比对结果呈现出与空间比对不一致的结果。由于酶分子的半理性改造在原理分析上通常偏好基于空间比对的结果,故最终选择了G220 作为改造位点。
2.3 AdIR1 的分子改造结果
通过分子改造,虽然在G220 残基的突变中并未发现酶活有效提升的突变体,但在V118 和F172 位点的单突变尝试中筛选到几个活力可能提升的突变体。通过对其宿主细胞进行培养、诱导表达以及亲和纯化,获得纯酶。随后,在30 ℃条件下测定纯化后的突变酶(M1~M4)对1-苯基-3,4-二氢异喹啉(底物浓度为0.1 mmol/L)活力,结果如表5 所示。

表5 AdIR1 突变体的测活结果Table 5 Activity assay results of AdIR1 mutants
通过测活发现,突变酶M1 与M3 并没有有效地增加AdIR1 对底物的催化活力,甚至在M2 中,分子改造使得其活力下降。而M4 突变有效增加了酶活,提升了1.9 倍。因此,通过对F172 位点的改造,本文得到了使AdIR1 活力显著提升的突变体。随后通过基因测序发现该突变型为F172Y。
2.4 亚胺还原酶突变体的酶学性质表征
2.4.1 pH 对酶活力的影响 通过使用不同的缓冲液,得到了在不同pH 条件下母本(WT)与突变体F172Y的催化活力特征曲线,结果如图3 所示。与母本相似,突变体F172Y 对pH 呈现出典型的钟形曲线,且最适pH 同样为 7.0。
2.4.2 热稳定性表征结果 对母本和突变体F172Y稀释酶液在30、40 ℃和50 ℃条件下保温一定时间后取样,用底物1-苯基-3,4-二氢异喹啉测定酶活,得到其在不同温度下残余活性的变化曲线,结果如图4所示。半衰期结果由Excel 进行曲线模拟所得,结果如表6 所示。
结果表明,突变体F172Y 虽然仅突变了一个位点,但是热稳定性有显著提升,其在30 ℃和40 ℃条件下96 h 保温后酶活力损失低于20%,并且拟合得到的t1/2在30、40 ℃以及50 ℃条件下分别是母本的4.6 倍、6.3 倍和3.9 倍。其中原母本在40 ℃条件下保温12 h 残留活性不足70%,而突变体F172Y 却保持了91%的残留活性,与30 ℃条件92%的残余活性基本保持一致。因此后期的反应进程表征以及制备反应将选用40 ℃作为反应条件。

图3 pH 对母本(a)与F172Y(b)酶活的影响Fig. 3 Effect of pH on the enzyme activity of WT(a) and F172Y(b)

图4 母本(a)与F172Y(b)的热稳定性Fig. 4 Thermostability of WT (a) and F172Y (b)
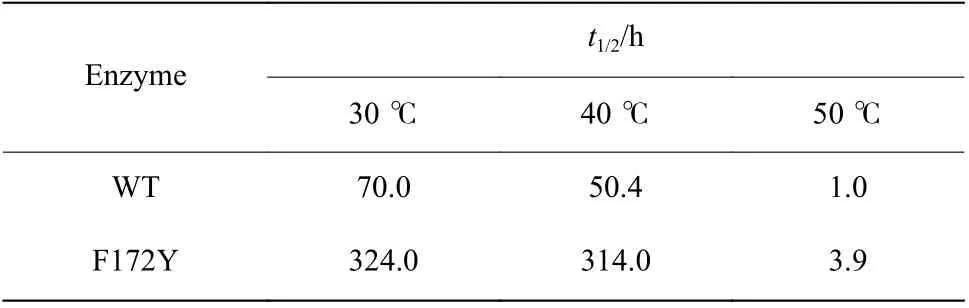
表6 F172Y 与母本的半衰期Table 6 Half-lives of WT and F172Y at different temperatures
2.4.3 动力学表征结果 利用不同浓度的1-苯基-3,4-二氢异喹啉为底物,本文分别测定了母本与突变体F172Y 的底物动力学特征曲线,结果如图5 所示,并依据动力学特征曲线拟合计算出动力学参数,结果如表7 所示。
通过以上实验结果表明,WT 与F172Y 对底物1-苯基-3,4-二氢异喹啉皆不存在底物抑制,意味着其在未来的工业生产上具有非常优质的应用前景。此前,通过F172Y 单点的突变,使该酶在Km保持不变的情况下,kcat有了3.4 倍的提升,即催化效率常数(kcat/Km)提升了3.4 倍。而温度提高时催化效率常数提升不明显,主要因为kcat有显著提升的同时Km亦有提升。活力测定表明,在40 ℃且饱和的底物浓度条件下,比活可达46 mU/mg,这比30 ℃条件下的活力(15.3 mU/mg)提高了约3 倍。
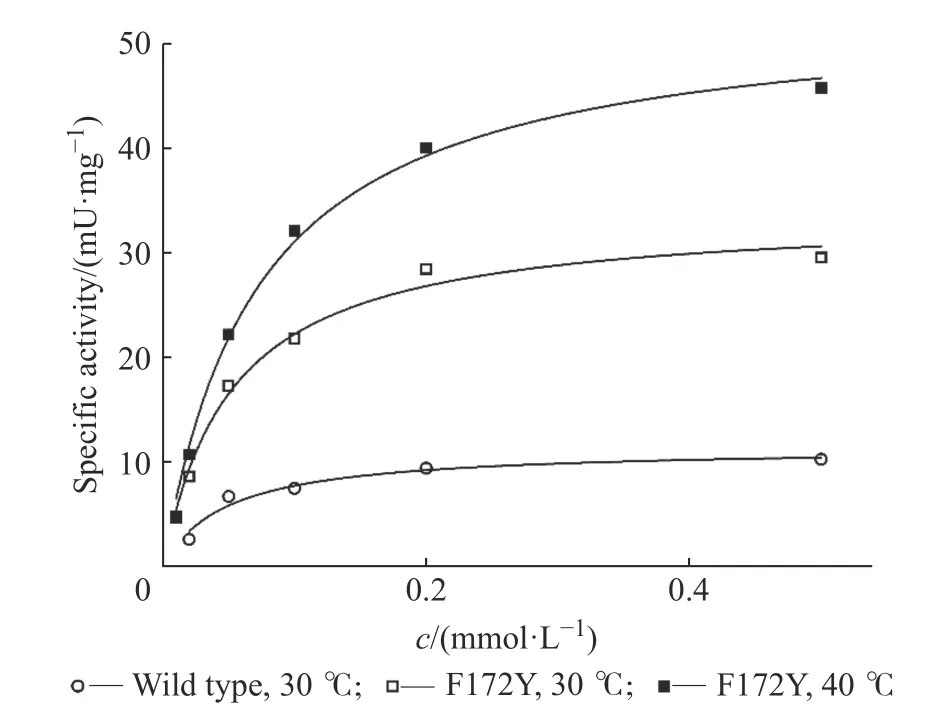
图5 母本与F172Y 的动力学特征曲线Fig. 5 Kinetic characteristic curves of WT and F172Y
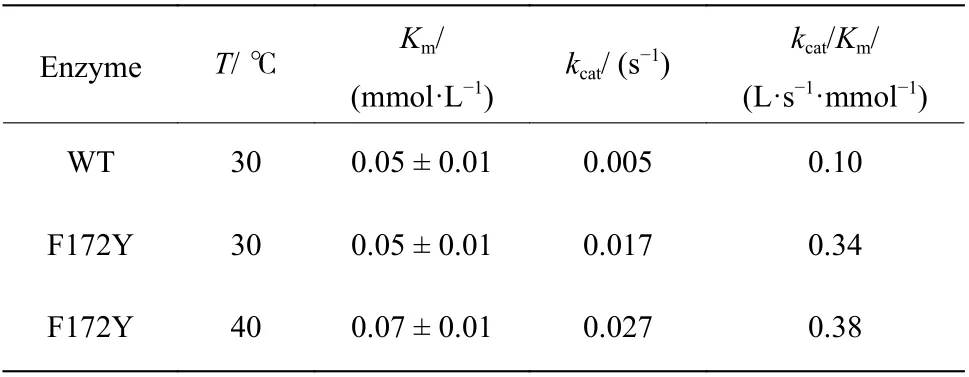
表7 母本与F172Y 的催化动力学参数Table 7 Kinetic parameters of WT and F172Y
2.5 亚胺还原酶母本与突变体的反应进程对比
通过对母本与突变体F172Y 在30 ℃、1 mmol/L底物浓度条件下的反应进程表征,本文得到了两者的反应进程曲线,结果如图6 所示。
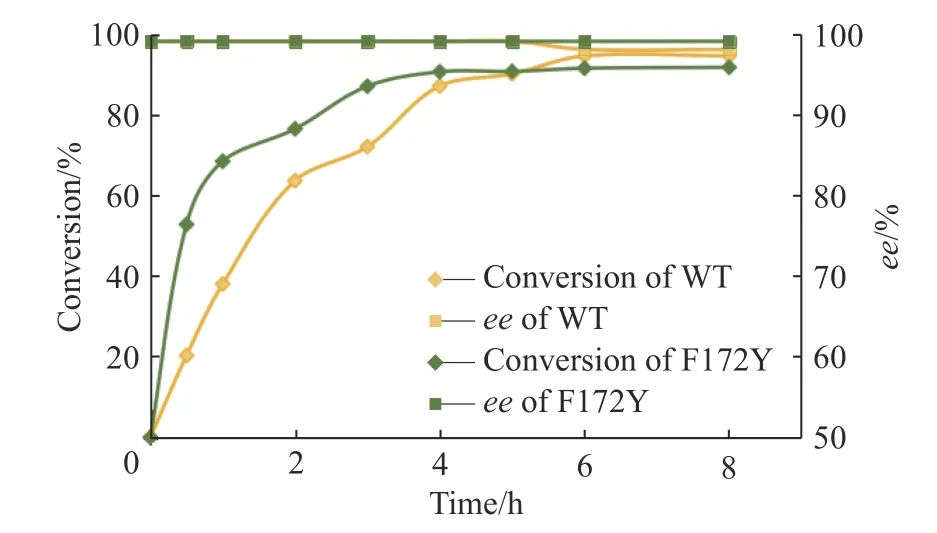
图6 母本(黄色)与突变体F172Y(绿色)的反应进程曲线Fig. 6 Reaction process curves of WT (yellow) and F172Y(green) of AdIR1
以上结果表明,母本可以在6 h 实现94%的转化率,产物ee值为98%。而突变体F172Y 并没有因为点位突变而影响其对映体选择性,其在前3 h 基本完成转化,实现90%的转化率和99%的ee值,后续过程中转化率慢慢提升至92%,ee值始终保持在99%。
2.6 1-苯基-1,2,3,4-四氢异喹啉的克级制备反应
2.6.1 制备反应的最适温度筛选 前期的酶学性质表征结果表明突变体F172Y 催化合成1-苯基-1,2,3,4-四氢异喹啉的最适催化温度为40 ℃。由于制备级反应的底物浓度提升到了5 mmol/L,在此浓度下本文考察了突变体F172Y 不同温度下催化合成1-苯基-1,2,3,4-四氢异喹啉的反应效果,结果如表8 所示,在1 mL 反应体系中,酶在40 ℃条件下的确表现出了比30 ℃更高的转化率,其在40 ℃反应6 h 的结果与30 ℃反应12 h 的结果一致。因此,确定40 ℃为制备反应的温度条件。
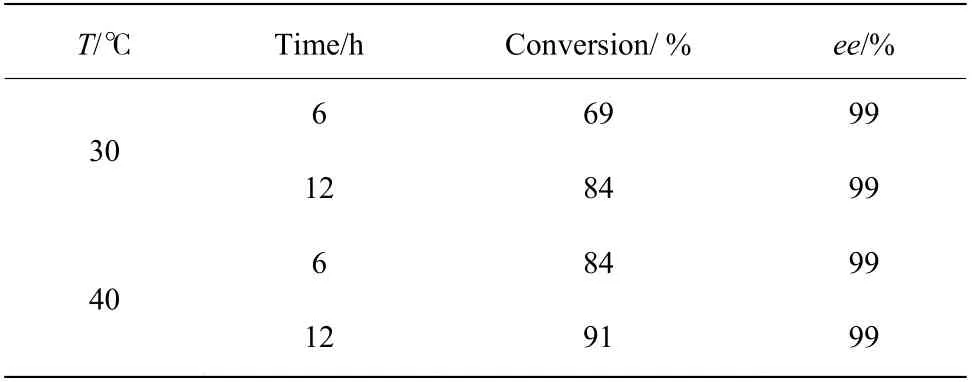
表8 F172Y 在30 ℃与40 ℃下的反应转化结果Table 8 Reaction conversion results of F172Y at 30 ℃ and 40 ℃
2.6.2 最适助溶剂体积分数筛选结果 除了最适温度,本文还对最适助溶剂体积分数进行了考察。为了降低DMSO 加入量的误差,反应体系扩大到了10 mL级别,结果如图7 所示。结果表明, φ =2%的DMSO加入量的转化效果略优于1%与5%,且远优于10%。因此将 φ =2%的DMSO 确定为最适助溶剂。另外,该反应条件下4 h 便完成了92%的转化,该结果高于最适温度筛选中6 h 的转化率,原因可能是10 mL 反应器相对于振荡反应器具有更佳的传质、传热效果。

图7 不同助溶剂浓度下的反应曲线Fig. 7 Reaction curves with different cosolvent volume fractions
2.6.3 克级制备反应 经过以上的反应条件优化,本文将反应体系扩大到2 L,在5 L 发酵罐中开展反应,底物1-苯基-3,4 二氢异喹啉的上载量为10 mmol(2.05 g),反应进程曲线如图8 所示。由图8 可以看出,12 h 后反应转化率仅维持在85%,且基本不再变化;反应24 h 后对反应进行淬灭,通过柱层析纯化得到0.34 g 的(S)-1-苯基-1,2,3,4-四氢异喹啉产物(得率为16.4%)。
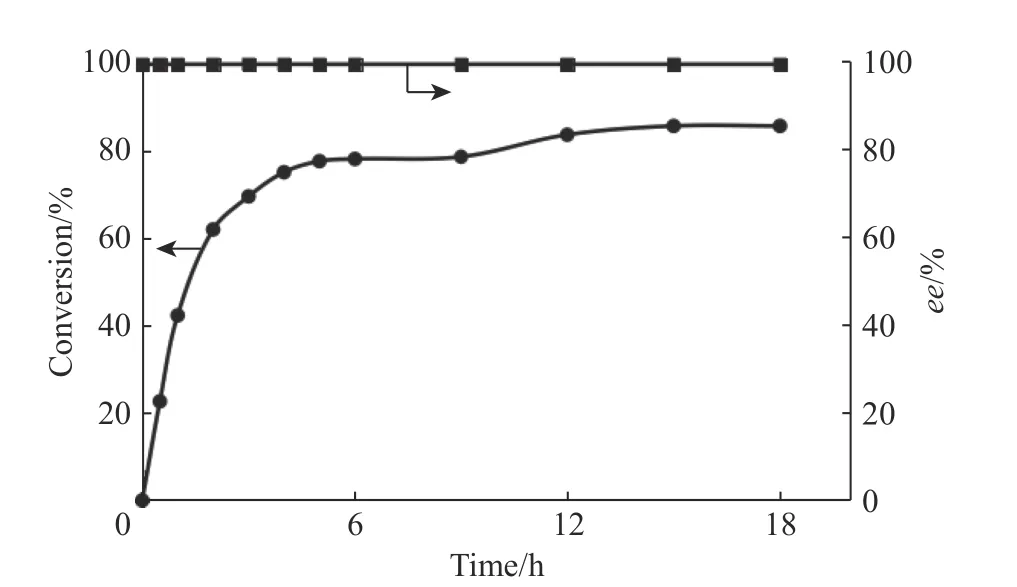
图8 克级制备反应的反应进程曲线Fig. 8 Reaction process curves of gram-scale preparative reaction
3 结 论
以反应转化率和对映体选择性结果较优的AdIR1 作为分子改造的起始酶,得到了突变体F172Y,其在30 ℃条件下酶活为15.3 mU/mg,相较于母本提升了1.9 倍,催化效率(kcat/Km)相较于母本有3.4 倍的提升。同时突变体F172Y 具有更优的热稳定性,在30 ℃和40 ℃条件下的t1/2分别是母本4.6 倍和6.3倍。另外,母本与突变体都未发现底物抑制情况。在催化合成1-苯基-1,2,3,4-四氢异喹啉的反应进程中,突变体F172Y 具有更快的转化效果,相较于母本需要6 h 基本完成反应转化,其仅需要3 h,且ee值维持在99%。在最适反应条件(40 ℃, φ =2% DMSO)下,利用突变体F172Y 作为催化剂开展了S构型1-苯基-1,2,3,4-四氢异喹啉的克级制备反应,反应12 h后可达85%的转化率,通过柱层析分离得到了0.34 g产物,分离得率为16.4%,展示出所开发的突变酶F172Y 不对称合成手性1-苯基-1,2,3,4-四氢异喹啉的实用性。
