降香黄檀不同种源/家系种质差异研究
2022-08-27罗水兴郭俊誉裴男才林明平李小飞
罗水兴, 郭俊誉, 孙 冰, 裴男才, 林明平, 李小飞
(中国林业科学研究院热带林业研究所, 广东 广州 510520)
降香黄檀(DalbergiaodoriferaT. Chen) 是豆科(Leguminosae) 蝶形花亚科黄檀属(Dalbergia)大乔木,又名海南黄花梨、降香、花梨木等[1],因其心材的珍贵稀少而备受关注,是制作高级红木家具、工艺制品、乐器和进行雕刻、镶嵌,美工装饰等的上等材料[2]。其根又名降香,可入药,几百年来被广泛用于止血、调气、散血瘀、止痛[3-5],木材精油可作为香料的定香剂。降香黄檀为海南特有乡土珍贵树种,多分布在山脊、陡坡、岩石裸露的贫瘠干旱地带等低山丘陵地区,天然分布于半落叶季雨林中,多以零星或团聚状散生,少数可成小片林[6]。由于其生存环境恶劣和长期人为砍伐,现存野生资源极其稀少,目前多为人工引种种植,海南、广东、福建、广西等省均有分布。
降香黄檀心材形成极为缓慢,树龄超过 6 年时开始形成心材,心材占总茎材比例非常小[1]。关于降香黄檀的研究大多集中在引种栽培[7]、播种育苗[8]、心材形成促进[9]、成林抚育[10]、抗逆性[2,11]等方面。优良种源(家系)选育方面,洪舟等[6]对广东省阳江市 11 年生降香黄檀种源试验林进行调查分析,初步挑选出白沙种源可作为适宜广东地区造林的降香黄檀优良种源。麻永红等[1]对 49 个 6 年生试验林的降香黄檀生长指标进行综合分析,初步筛选出 5 个生长优良的家系。陈英强[12]对 6 年生试验林的降香黄檀生长指标进行综合分析,初步筛选出适合闽南沿海地区种植的优良家系 10 个。面对降香黄檀人工林庞大的市场需求,优良种源(家系)研究依旧显得薄弱。本研究收集华南 10 个地区 66 个家系,对比不同产地降香黄檀种质资源质量以及苗期生长表现,以期为华南地区优质降香黄檀种质资源筛选提供科学依据。
1 材料与方法
1.1 材 料
降香黄檀种源采于广东广州、肇庆、茂名、英德,广西凭祥,海南海口、临高、白沙、东方、乐东共 3 省 10 个市(县)66 个家系,以地名和种号命名(地名1-n),产地及产地自然条件详见表1。种子于 2020 年 11 月至 2021 年 1 月采集,每家系500 g,选择15年以上树龄优树采种带回,自然晾干。育苗试验在中国林业科学研究院热带林业研究所苗圃试验地(113°23′4.64″E、23°11′27.22″N)进行,采用随机区组试验, 3 次重复。
1.2 方 法
1.2.1种子形态及发芽率测定
每家系随机取 30 个荚果,4次重复,测量荚果长度和宽度,称重后人工搓揉去掉边缘果翅即为生产用果种,计量出种率。每家系随机选取 30 粒降香黄檀种子,游标卡尺测定种子的长、宽、厚,重复 3 次,每家系随机选取 100 粒降香黄檀种子计算千粒重,3 次重复。
各家系种子在平底塑料沙盆(15 cm×25 cm)中发芽,30粒一盆,3次重复播种前苗床、沙盆、种子先用蒸馏水洗净,用0.5% 的高锰酸钾进行表面杀菌,河沙用蒸馏水洗净后0.5%高锰酸钾消毒,间隔 2 d 后播种,播种后定期浇水保持基质一定湿度,间隔 24 h 观察一次,连续 3 d 没有新增种子发芽数时计算发芽率。

表1 降香黄檀种子产地基本情况Table 1 Basic situation of Dalbergia odorifera seed producing areas
HH为海南海口;HL为海南临高;HB为海南白沙;HD为海南东方;HJ为海南尖峰永涛;GP为广西凭祥市。
1.2.2幼苗生长性状测定
降香黄檀种子发芽长出 4 片真叶时,移栽到1.5 kg黄心土育苗袋中继续生长 6 个月,每家系10 株,3次重复,采取完全随机区组设计,按照苗圃常规育苗方法进行日常管护并及时浇水和除草,每月施肥 1次(5%复合肥溶液)、消毒2次(1 000 倍多菌灵)。之后测定苗高、地径,并将整株取出冲洗干净,105 ℃ 杀青 20 min,75 ℃烘干至恒重,测定根、径、叶生物量。
1.3 数据处理和统计分析
数据统计用 Excel 2017软件进行,运用 SPSS 24.0软件进行方差分析、Duncan 多重比较和主成分分析,运用Orgin 2021软件进行相关性分析和作图,对不同产地的种子和幼苗性状及其与各产地地理气候因子进行相关性分析,对各性状进行主成分分析。
2 结果与分析
2.1 不同家系降香黄檀种子表型性状及发芽率差异
从表2可以看出,10个产地66个家系降香黄檀种子表型性状和发芽率在家系间均表现出显著差异,表型性状和发芽率的变异系数较大,变幅为8.99%~25.28%。表型性状中变异系数超过10%的有5项,包括荚果千粒重(22.34%)、去翅千粒重(23.80%)、出种率(20.17%)、种子千粒重(25.28%)、种厚(14.20%),发芽率的变异系数为16.11%。
66个家系降香黄檀种子的荚果长3.79~6.54 cm,荚果宽1.42~2.12 cm,荚果千粒重96.42~322.56 g,去翅荚果千粒重74.23~260.64 g,荚果出种率25.00%~98.33%,种子千粒重16.46~92.34 g,种子长8.29~11.31 mm,种子宽4.52~7.71 mm,种子厚1.04~2.44 mm,种子发芽率16.67%~97.78%。
不同家系降香黄檀种子中荚果长排名前五的依次为HH 3、GM 4、HB 2、HB 6、HH 2,荚果宽排名前五的依次为GM 2、HD 16、HB 2、HD 8、HH 2;荚果千粒重排名前五的依次为HD 8、GM 2、CAF 2、GZ 3、HD 11;去翅荚果千粒重排名前五的依次为HD 8、CAF 2、GM 2、GZ 3、HL 5;荚果出种率排名前五的依次为HD 15、CAF 2、HB 4、HD 2、HD 8;种子千粒重排名前五的依次为GM 2、HL 2、GP 1、HL 1、HD 8;种长排名前五的依次为GM 2、HD 8、GM 1、HH 2、GP 2;种宽排名前五的依次为HD 16、GM 2、HL 4、HD 8、HB 2;种厚排名前五的依次为HL 1、HH 3、GP 1、HL 2、CAF 3;发芽率排名前五的依次为HD 15、HD 6、HB 9、HB 6、HD 13。
2.2 降香黄檀不同家系幼苗性状差异
降香黄檀幼苗性状66个家系间均表现出显著差异性(表3),且幼苗表型性状存在较大的变异性,性状变异系数的变幅为8.49%~33.03%,变异系数超过10%的指标有茎干重(33.03%)、叶干重(27.87%)、根干重(22.62%)和苗高(15.71%),茎干重变异系数最大,地径变异系数最小(8.49%)。
10个产地66个家系降香黄檀幼苗苗高16.67~35.80 cm,地径 2.74~4.41 mm,根干重0.54~1.24 g,茎干重0.52~1.89 g,叶干重0.79~2.88 g。不同家系间幼苗平均高度排名前五的依次为GM 4、HL 1、HH 1、HL 2、HD 2;地径排名前五的依次为GM 4、HJ 12、HD 6、HH 1、HJ 7;根干重排名前五的依次为HB 5、HJ 13、HJ 7、HJ 14、GZ 3;茎干重排名前五的依次为CAF 3、GZ 1、GM 4、GZ 2、HB 5;叶干重排名前五的依次为GM 4、HB 5、GZ 3、GZ 1、GZ 2。

表2 不同家系降香黄檀种子表型性状Table 2 The phenotypic traits and germination status of Dalbergia odorifera seeds from different families

表2(续)
2.3 降香黄檀种子及幼苗表型性状和环境因子的相关性分析
对10个产地66个家系降香黄檀种子和幼苗的15个性状及种源地环境因子的相关性分析,结果见图1。由图1可知,海拔(A)与种子千粒重(TGW)、种子发芽率(GT)呈显著正相关;年均温(AT)与荚果宽(PW)、荚果千粒重(TGWP)、去翅荚果千粒重(TGWW)、种子千粒重、种长(SL)、种宽(SW)、种子发芽率呈显著正相关;年降雨量(AP)与出种率(SP)呈显著负相关,与叶干重(LW)呈显著正相关;荚果长(PL)与荚果宽、荚果千粒重、去翅荚果千粒重、种子千粒重、种长、种宽、苗高呈显著正相关,与出种率呈显著负相关;荚果宽与荚果千粒重、去翅荚果千粒重、种子千粒重、种长、种宽、种厚(ST)、苗高(PH)、地径(GD)、茎干重(SWG)呈显著正相关;荚果千粒重与去翅荚果千粒重、种子千粒重、种长、种宽、种厚、苗高、地径、茎干重、叶干重呈显著正相关;去翅荚果千粒重与荚果千粒重趋势一致;出种率与发芽率显著正相关,与茎干重、叶干重呈显著负相关;种子千粒重与种长、种宽、种厚、苗高、地径、茎干重、叶干重呈显著正相关;种长与种宽、种厚、苗高、地径、茎干重、叶干重呈显著正相关;种宽与地径呈显著正相关;种厚与发芽率、苗高、茎干重呈显著正相关;苗高与地径、根干重(RW)、茎干重、叶干重呈显著正相关;地径与根干重、茎干重、叶干重呈显著正相关;根干重与茎干重、叶干重呈显著正相关;茎干重与叶干重呈显著正相关。
2.4 不同家系种子和幼苗表型性状的主成分分析及综合评价
对降香黄檀种子性状和子代幼苗的 15 个指标进行主成分分析,结果(表4)表明,前 5 个主成分的累积贡献率达到 80.29%,说明前 5 个主成分可以有效概括家系种子性状和子代幼苗性状的 15 个指标。5 个主成分的贡献率依次为22.53%、21.11%、13.33%、12.27%、11.05%。5个主成分中,如果特征向量值为正,说明此成分对该主成分的影响为正向,相反,如果特征向量值为负值,说明此成分对该主成分的影响为负向。

表3 降香黄檀不同家系幼苗性状差异Table 3 Differences in seedling traits of Dalbergia odorifera from different families
主成分分析可以有效比较不同家系间种子性状和子代幼苗生长状况的综合质量,对良种和优良子代植株的选择及开发利用有较大的应用价值。用X1、X2、X3、X4、X5分别代表这 5 个主成分,用A1、A2、A3……A13、A14、A15分别代表 15 个指标,得到以下线性方程:
X1=0.07A1+0.12A2+0.43A3+0.42A4+0.16A5+0.03A6+0.14A7-0.06A8-0.13A9-0.11A10-0.07A11-0.22A12-0.10A13+0.03A14+0.08A15
X2=-0.06A1-0.12A2-0.02A3+3.44×10-4A4-0.01A5-0.04A6-0.03A7-0.08A8-0.04A9+0.03A10+0.22A11+0.25A12+0.33A13+0.24A14+0.28A15
X3=0.20A1+0.22A2-0.17A3-0.17A4-0.05A5+0.05A6+0.06A7+0.50A8-0.15A9+0.07A10+0.06A11+0.50A12+0.05A13-0.21A14-0.20A15
X4=-0.08A1+0.04A2-0.17A3-0.17A4-0.18A5+0.32A6+0.11A7-0.07A8+0.66A9+0.25A10+0.04A11-0.21A12-0.12A13+0.14A14-0.05A15
X5=-0.26A1-0.11A2+0.06A3+0.12A4+0.59A5-0.03A6+0.02A7+0.02A8-0.06A9+0.37A10-0.05A11+0.11A12+0.15A13-0.08A14-0.06A15
以1、2、3、4、5主成分对应的贡献率b1、b2、b3、b4、
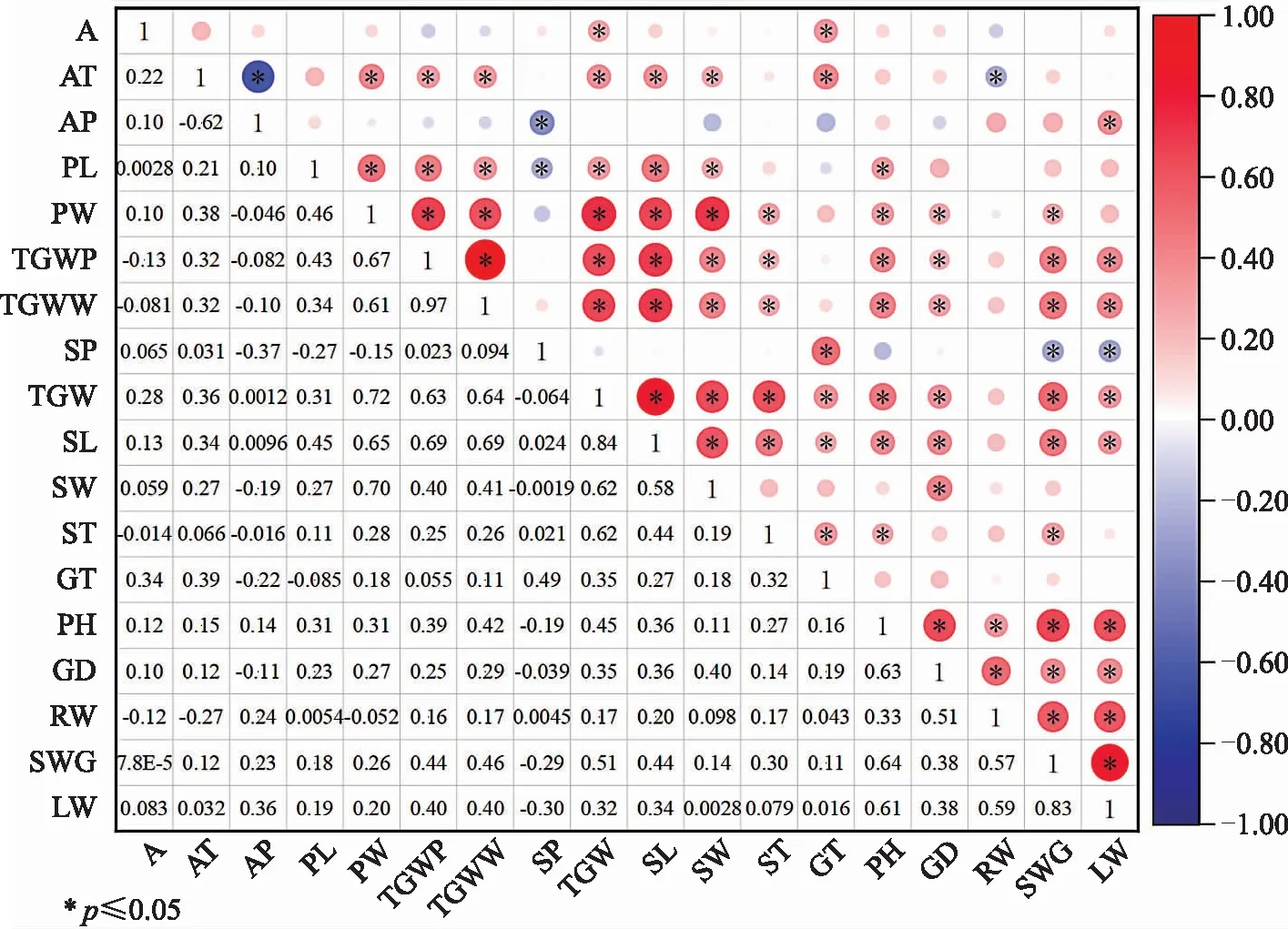
图1 降香黄檀种子及幼苗表型性状和环境因子的相关性分析Fig.1 Correlation analysis of phenotypic traits of Dalbergia odorifera seeds and seedlings and environmental factors
b5作为权重系数,建立综合评价模型:
C=b1X1+b2X2+b3X3+b4X4+b5X5,式中:C为综合评价得分。

表4 降香黄檀种子和幼苗15个表形性状主成分分析Table 4 Principal component analysis of 15 phenomorphic characters of Dalbergia odorifera seeds and seedlings

表5 不同产地/家系降香黄檀种子综合得分情况Table 5 Comprehensive scores of Dalbergia odorifera seeds from different producing areas/families
由表5可知,10 个产地 66 个家系降香黄檀种子综合得分排名前十依次为HD 8(54.62)、CAF 2(51.15)、GM 2(49.25)、GM 1(44.17)、HD 11(43.72)、GZ 3(43.20)、HL 5(43.00)、HD 15(42.71)、HD 4(42.62)、HL 2(41.27),66 个家系降香黄檀种子中HD 8最为优良;排名后十依次为 HJ 9(30.94)、HD 10(30.63)、HB 11(30.57)、HJ 5(30.44)、HH 3(30.42)、HB 1(28.00)、HJ 4(27.90)、HB 12(27.46)、HJ 2(25.07)、HJ 1(19.86),最差的是HJ 1。
3 讨 论
植物稳定遗传性状的表型多样性揭示了群体的遗传规律和变异程度,而且有助于评价群体的遗传多样性水平,也是遗传育种工作的基础,这些表型是环境与遗传因素双重作用的结果,而表型变异是植物适应环境变化的表现,同时促进了物种进化[13-15]。形态性状变异是遗传变异的一部分,形态变异越大,遗传变异的可能性就越大。通过对10个产地66个家系降香黄檀种子及幼苗性状测定发现,降香黄檀种子和幼苗表型性状具有丰富变异,不同产地及家系间均出现显著差异,说明降香黄檀具有明显的地理遗传分化。降香黄檀66个家系种子与幼苗性状的变异系数范围在8.49%~33.03%之间,说明降香黄檀具有优良种源选择潜力,与葛玉珍等[16]的研究结果相似。降香黄檀荚果长、荚果宽、种长、种宽、地径变异系数较小,说明这些性状是较为稳定的表型性状。形态不是判定种子优劣的唯一指标,种子的发芽率也可以反映种子的劣变程度,是种子活力的重要指标[17],本研究不同家系中,海南东方15号发芽率最高,为97.78%。
幼苗生长表型性状是可以直观地反映苗木长势的指标[18],本研究发现,降香黄檀种子的千粒重、种长、种宽、种厚与幼苗苗高、地径、地上干重存在显著正相关关系,说明根据种子性状可对降香黄檀优树幼苗生长进行早期选择,与李峰卿等[19]研究结果一致。同时,降香黄檀幼苗根干重与种子性状并不存在显著相关性,与苗高、地径、地上部分干重呈显著正相关,相较于苗高、地径,根干重与地上干重(茎、叶)相关性更强,这与植物光合作用资源分配有关,植物在幼苗阶段,其生长的最佳状态是将更多的资源分配给叶、茎和根的功能组织,通过叶面积、茎高度和根生物量的增加来获取更多的资源,以达到更大的生存适合度[20]。
植物表型性状是遗传与环境共同作用的结果,环境因素与植物的生长发育之间有着紧密的联系,不仅影响种子的发育,也影响林木生长[21]。本研究发现,海拔和年均温显著影响降香黄檀种子大小和发芽率,这与林玮等[22]研究任豆(Zeniainsignis)种子、刘桂丰等[23]研究白桦种子得出的结果一致。此外,年均温和降雨量也显著影响降香黄檀幼苗生长性状。这说明在降香黄檀引种栽培时海拔、年均温和年降雨量也应作为首先考虑的重要因素。
为筛选出综合表现优良的降香黄檀家系种苗,本研究采用主成分分析法对10个产地66个家系降香黄檀种子和幼苗性状进行综合比较,发现66个家系中HD 8、CAF 2、GM 2、GM 1、HD 11为表现较优的5个家系。实验集中于降香黄檀种子、苗期的评价筛选,缺乏后期生长评价,但研究结果对华南地区降香黄檀种质资源选优仍具有一定的参考意义。
