苦荞蔗糖转运体家族FtSUCs的鉴定与生物信息学分析
2022-08-27梁成刚关志秀韦春玉孟子烨石桃雄
梁成刚,汪 燕,关志秀,韦春玉,邓 娇,黄 娟,孟子烨,石桃雄
(贵州师范大学 生命科学学院 荞麦产业技术研究中心, 贵州 贵阳 550001)
蔗糖不仅是重要的信号因子,还是植物体内长距离运输的主要碳水化合物,因此,蔗糖对于植物的生长和发育至关重要。蔗糖在植物体内的转运主要是通过质外体运输与共质体运输两个途径来实现的。植物体内共质体与质外体运输的主要不同点是蔗糖由维管薄壁细胞向伴胞/筛管复合体的运输。共质体运输途径中蔗糖通过胞间连丝进入伴胞,而质外体运输途径中蔗糖则需要由蔗糖转运体蛋白(sucrose transporter,SUC)参与跨膜运输至伴胞/筛管复合体。植物体内SUCs蛋白是由5个基因家族编码,又根据SUC序列的同源性与生理功能可归为3类。类型Ⅰ由双子叶植物SUCs组成(如AtSUC1、AtSUC2、AtSUC5),类型Ⅱ(如AtSUT3)和类型Ⅲ(如AtSUC4等)由单子叶与双子叶植物SUTs共同组成。
苦荞起源于中国,富含蛋白质与类黄酮物质,营养保健价值高。由于栽培驯化时间较短,苦荞还存在无限花序、种子异步灌浆等习性,而光合同化产物不足与分配不当是导致苦荞结实率低、充实度差的主要原因。弄清苦荞蔗糖运输的生理与分子调控机制,对于苦荞生长发育与产量形成具有重要意义。近年来在拟南芥、水稻等模式植物中对基因的研究较为透彻。例如,拟南芥中2是调控蔗糖运输的关键基因,2功能缺失会导致蔗糖和淀粉超量积累,植株生长发育受阻。在水稻中,5个基因功能缺失均未出现类似2突变的表型特征,因此,普遍认为OsSUTs蛋白对水稻蔗糖的韧皮部装载贡献较小。Liang等报道了水稻蔗糖和淀粉超量积累的突变体2和3,其表型与拟南芥2突变体类似,其目标基因可能在调控水稻蔗糖运输中起关键作用。目前,尚未见有关苦荞蔗糖运输及其调控基因的相关报道,笔者将结合转录组测序数据库对苦荞进行生物信息学分析,初步明确基因家族的分类、蛋白理化特性、系统发育以及表达模式等,为苦荞蔗糖运输的基因表达调控等相关研究提供依据。
1 材料与方法
1.1 材料
登录拟南芥基因数据库(https://www.arabidopsis.org/about/datasources.jsp),查询拟南芥蔗糖转运体()基因序列。利用苦荞转录组数据库筛选苦荞基因(https://www.ncbi.nlm.nih.gov/bioproject/PRJNA007201)。
1.2 方法
1.2.1 苦荞基因序列分析
苦荞基因序列的鉴定和序列分析以拟南芥基因序列为参考模板,以苦荞转录组测序基因序列为数据库,利用本地BLASTN检索同源序列。利用在线网站(https://www.ncbi.nlm.nih.gov/orffinder/)进行开放阅读框及编码氨基酸序列查询,利用在线网站(http://smart.embl-heidelberg.de/)进行蛋白跨膜结构域查询。利用在线网站(https://web.expasy.org/protparam/)进行FtSUCs蛋白分子量、等电点和疏水性查询。利用在线网站(https://www.genscript.com/psort.html和http://localizer.csiro.au/)进行蛋白亚细胞定位。利用在线网站(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)进行蛋白二级结构预测。利用在线网站(http://swissmodel.expasy.org/interactive)进行蛋白三级结构预测。利用MEGA5.0软件Cluster W进行多重序列比对分析,Neighbor-Joining进行进化树构建,重复1 000次。
1.2.2 苦荞基因的表达与相关性分析
利用苦荞幼苗发育过程(出苗后5 、10 、15 d)中茎的转录组数据,通过功能注释筛选差异表达的基因。利用MeV软件(https://mev.tm4.org/)进行差异表达基因的热图与相关分析。
2 结果与分析
2.1 苦荞FtSUCs基因及其编码蛋白序列
基于苦荞转录组测序数据库,进行基因筛选,结果鉴定到10个基因(表1)。苦荞基因序列长度为1 436~7 109 bp,其中,最长转录本序列长度为447~1 818 bp,编码氨基酸序列长度为148~605个。蛋白分子量为16 503.56~65 140.75 u,理论等电点为4.32~11.62,脂肪族氨基酸指数为60.54~111.72,蛋白质疏水性为-0.018~0.751。蛋白亚细胞定位预测发现,除FtPinG0001943900.01和FtPinG0001943500.01可能定位于细胞核外,其余8个FtSUCs蛋白均可能定位于内质网。蛋白序列比对分析发现,8个可能定位于内质网的FtSUCs蛋白序列一致性达到34.30%,保守位点达到20个。
2.2 苦荞FtSUCs基因序列系统发育树
利用苦荞与拟南芥序列构建进化树,结果发现,苦荞10个与拟南芥5个可聚为三类。其中,0001943500.01、0009584000.01、0005577300.01、0002608200.01与拟南芥2、1、5聚为一类,0000342600.01、0008441300.01与拟南芥4聚为一类,0000812400.01、0000184200.01、0001943900.01、000194410-0.01与拟南芥3聚为一类(图1)。
2.3 苦荞FtSUCs蛋白结构
对蛋白结构进行在线预测,发现FtSUCs蛋白二级结构主要由α-螺旋、无规则卷曲、延长链和β-折叠组成。10个FtSUCs蛋白中α-螺旋占12.86%~54.05%、β折叠占2.60%~6.16%、延长链占12.72%~20.27%、无规则卷曲占28.70%~59.46%。其中9个FtSUCs蛋白序列具有多个跨膜结构功能域,而FtPinG0001943900.01存在差异,其最长转录本编码蛋白不存在跨膜结构功能域,其他转录本开放阅读框(ORF)编码蛋白中存在跨膜结构功能域(表2)。
利用在线软件对FtSUCs蛋白进行三级结构预测,选取类型Ⅰ中FtPinG0005577300.01、类型Ⅱ中FtPinG0008441300.01和类型Ⅲ中FtPinG0000184200.01进行制图。如图2所示,3个FtSUCs蛋白结构主要以α-螺旋与不规则卷曲为主构成特殊空间结构。FtSUCs蛋白的α-螺旋数量与其跨膜结构域数量密切相关。
2.4 苦荞FtSUCs基因家族表达差异性
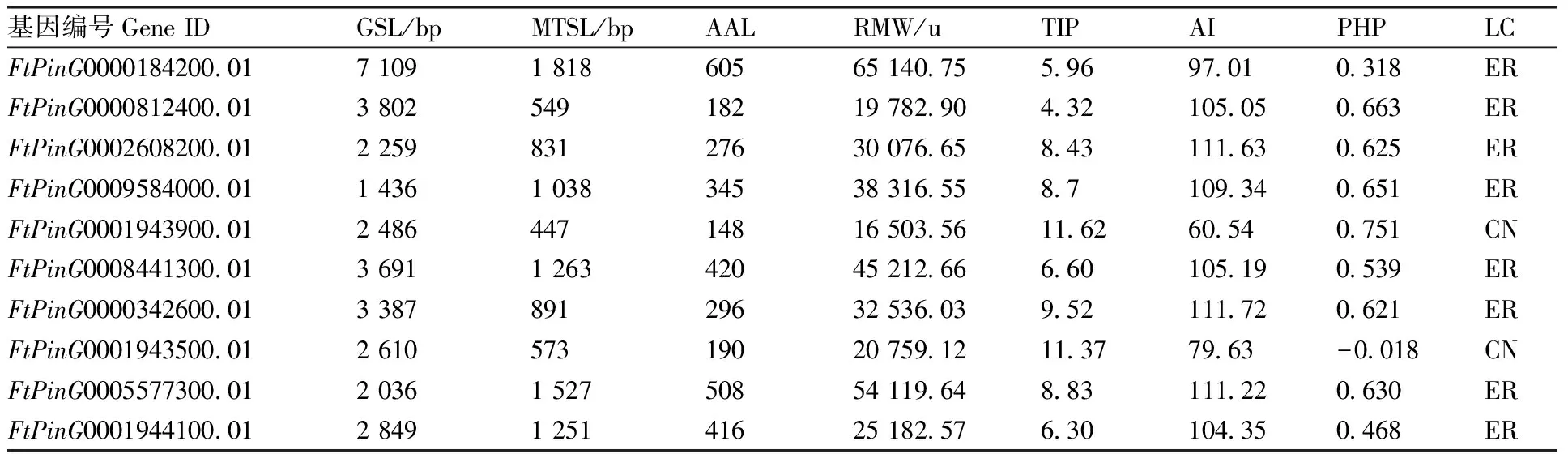
表1 苦荞FtSUCs基因序列分析
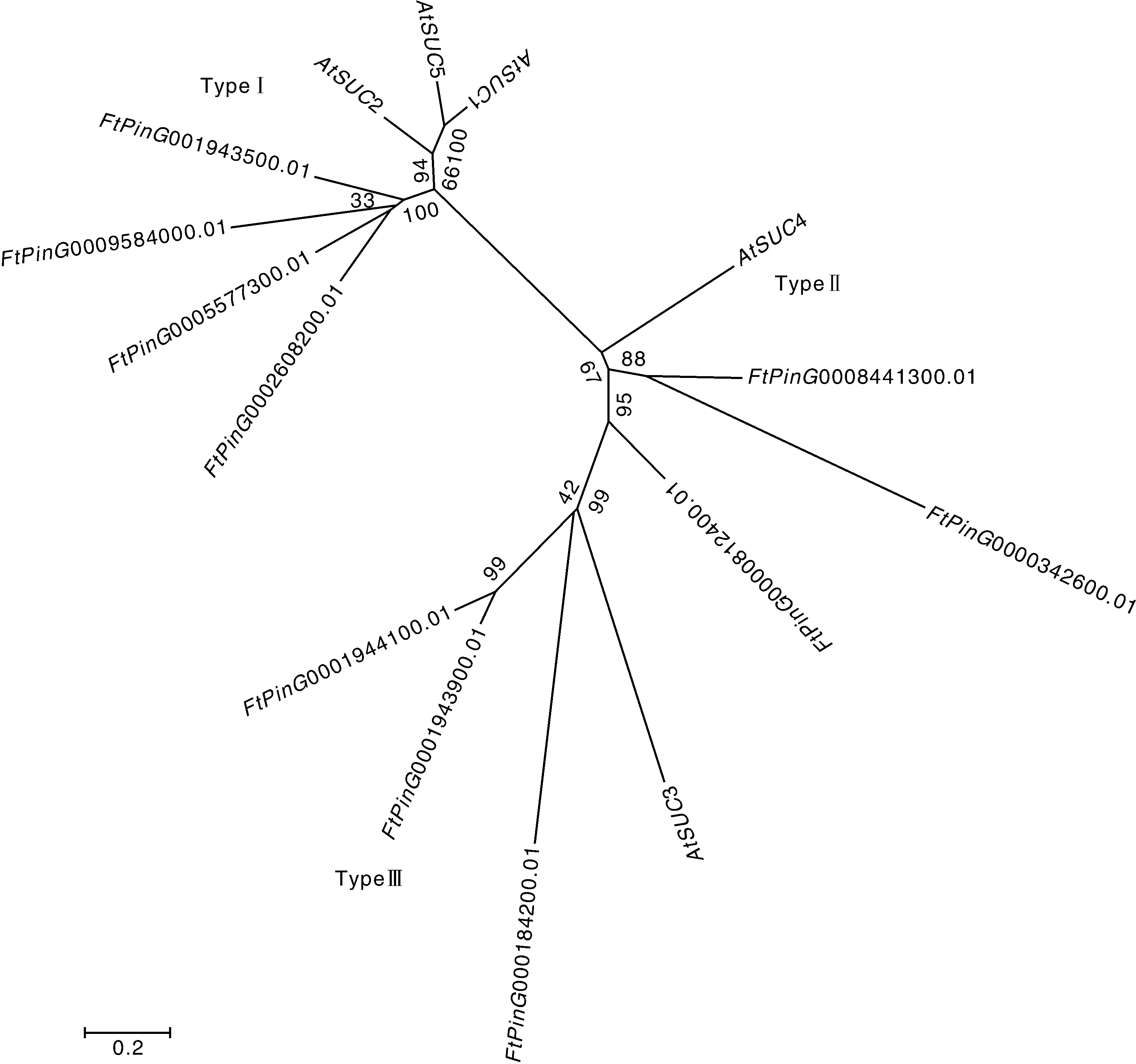
图1 苦荞FtSUCs序列的系统进化图Fig.1 Phylogenetic diagram of FtSUCs sequence in Tartary buckwheat
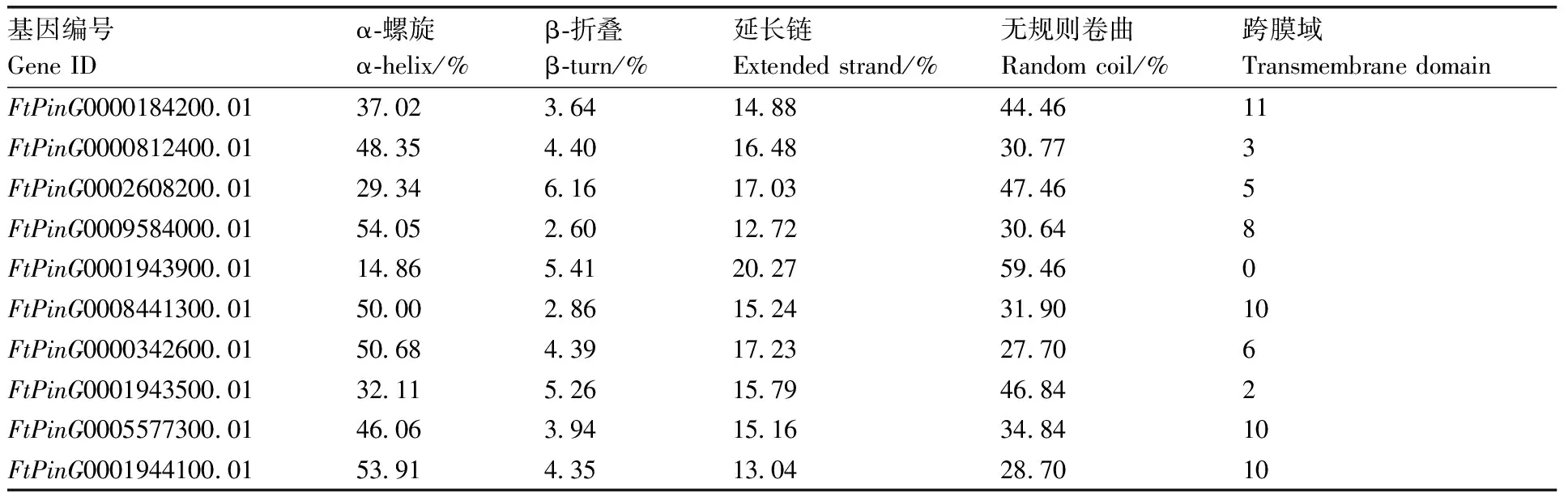
表2 苦荞FtSUCs蛋白结构
基于苦荞出苗后5 、10 、15 d茎的转录组测序分析,发现苗期不同发育阶段苦荞茎中基因表达模式不一致(图3-A)。其中,0001943500.01和0009584000.01在3个时期均稳定表达,另外8个基因表达存在显著差异,差异倍数(Fold Change)≥2,且错误发现率(FDR)<0.01。由图3得知,其中0009584000.01和000342600.01表达量呈先降低后升高趋势,0008441300.01和005577300.01表达量呈先升高后降低趋势,0000184200.01表达量呈逐渐升高趋势;0002608200.01、0001944100.01和0001943900.01在5 d时为高表达,10 d和15 d时为低表达。
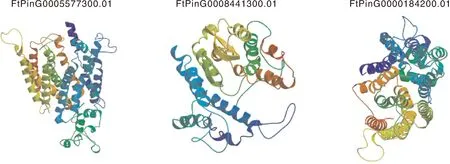
图2 苦荞FtSUCs蛋白的结构分析Fig.2 Protein structure of FtSUCs in Tartary buckwheat
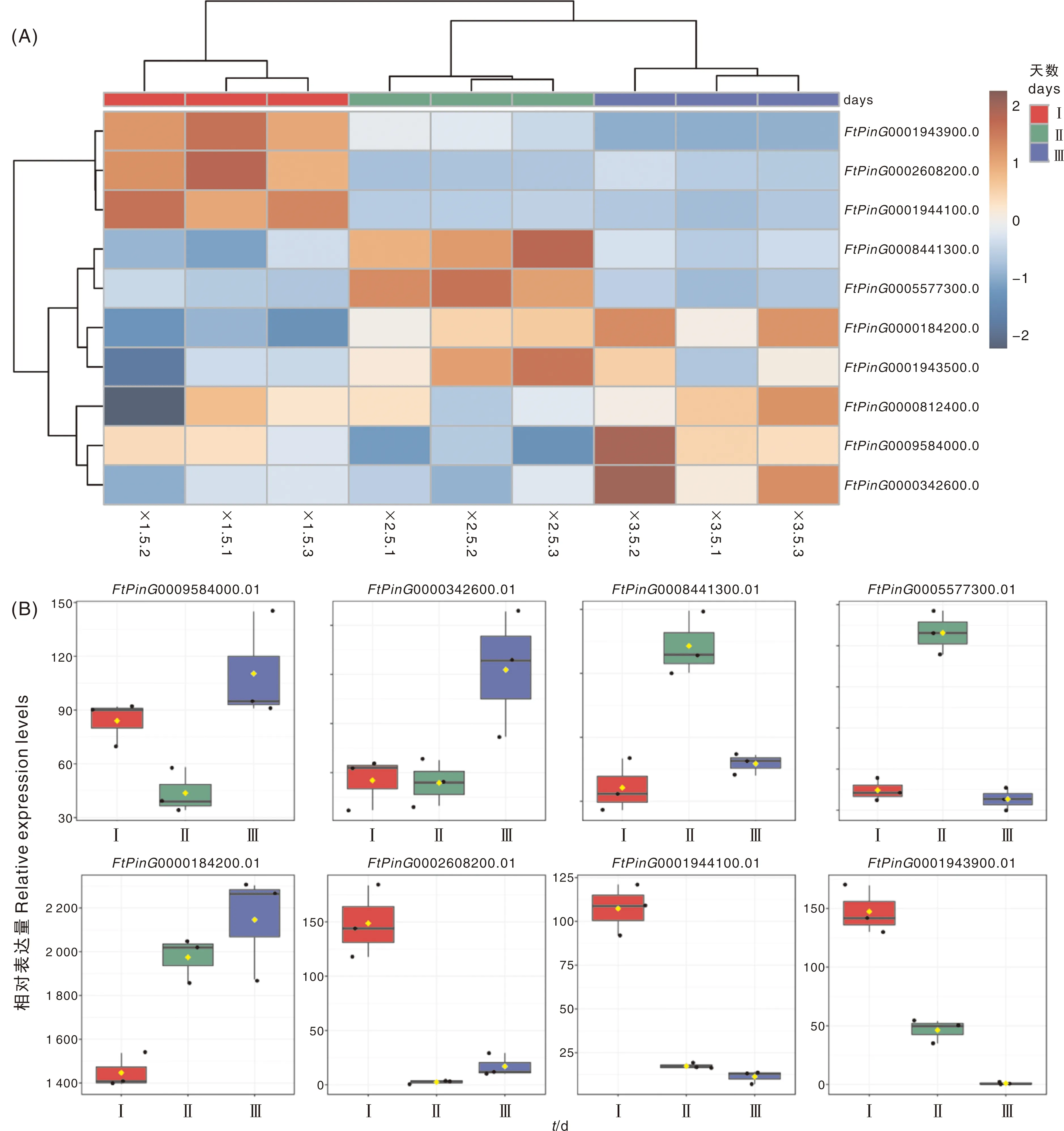
Ⅰ,出苗后5 d;Ⅱ,出苗后10 d;Ⅲ,出苗后15 d。Ⅰ, 5 d after emergence; Ⅱ, 10 d after emergence; Ⅲ, 15 d after emergence.图3 幼苗期苦荞FtSUCs的表达热图与DEGs基因表达分析Fig.3 Gene expression hotmap and DEGs expressing analysis of FtSUCs in Tartary buckwheat seedling
2.5 苦荞FtSUCs基因表达量相关性
对10个苦荞基因的表达量进行相关性分析,结果发现,0000184200.01与0000342600.01(=0.69,<0.05)、0001943500.01(=0.68,<0.05)均成显著正相关,与0002608200.01(=-0.80,<0.01)、0001943900.01(=-0.89,<0.01)、0001944100.01(=-0.89,<0.01)均成极显著负相关;0002608200.01与0001943900.01(=0.92,<0.01)、0001944100.01(=0.92,<0.01)均成极显著正相关,与0008441300.01(=-0.68,<0.05)成显著负相关;0009584000.01与0000342600.01(=0.69,<0.05)成显著正相关,与0008441300.01(=-0.72,<0.05)和0005577300.01(=-0.73,<0.05)均成显著负相关;0008441300.01与0001943500.01(=0.82,<0.01)和0005577300.01(=0.90,<0.01)成极显著正相关;0005577300.01与0009584000.01(=-0.73,<0.05)成显著负相关(图4)。
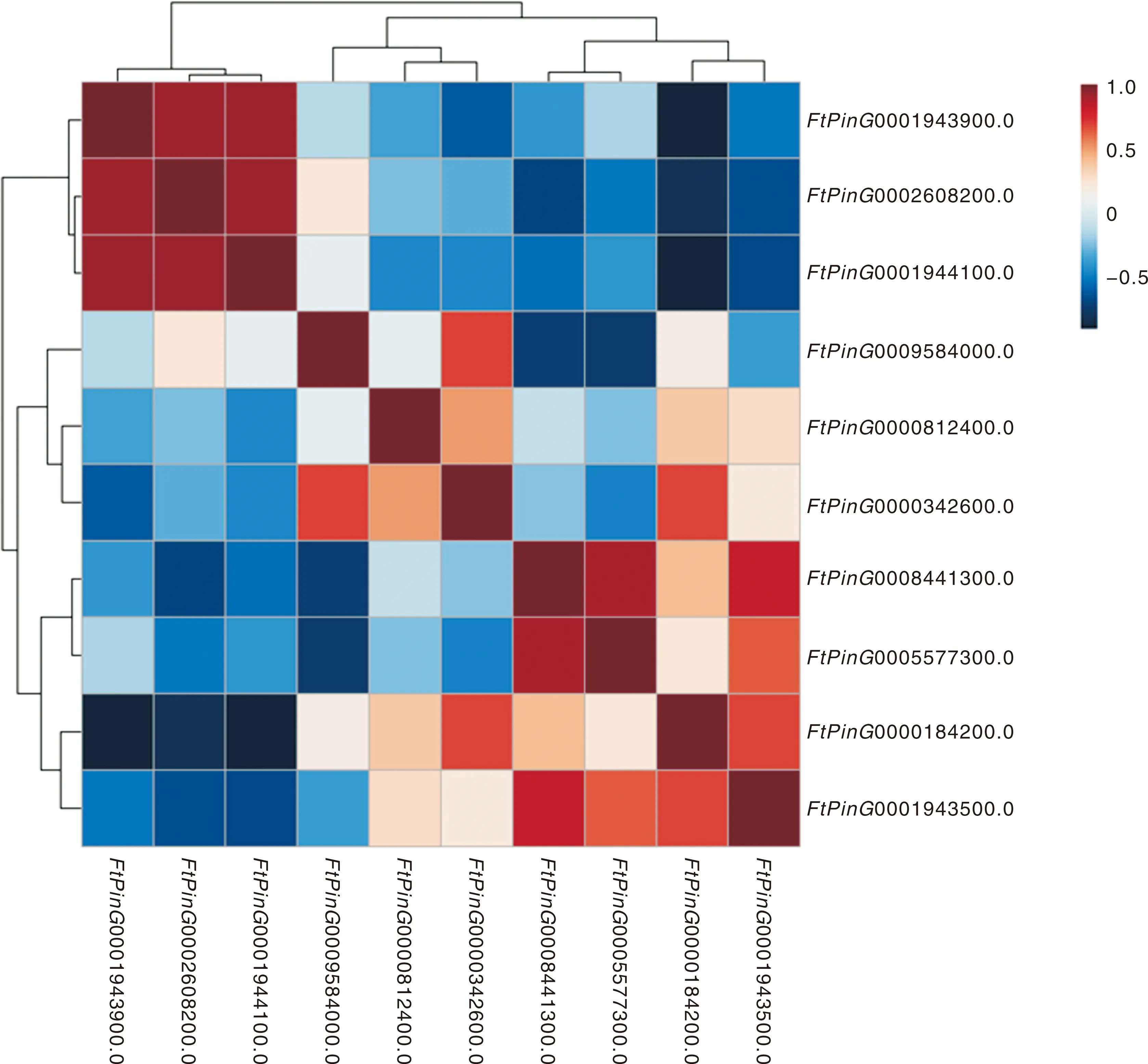
图4 苦荞FtSUCs基因表达的相关性分析Fig.4 Correlation analysis of FtSUCs expressing in Tartary buckwheat
3 讨论
基于前期苦荞茎转录组测序获得的数据库,本研究鉴定到10个蔗糖转运体编码基因。跨膜结构功能域是蔗糖转运体的典型结构域。通过氨基酸序列分析,发现9个苦荞最长转录本编码蛋白含有多个跨膜结构功能域,其中,FtPinG0000184200.01达到11个,FtPinG0005577300.01和FtPinG0001944100.01具有10个,FtPinG0001943900.01的最长转录本不存在跨膜结构功能域,但它其余的ORF中存在跨膜结构功能域,说明鉴定获得的10个均能参与跨膜运输。
植物SUCs包含5个家族,被归为3大类。系统发育分析表明,苦荞与5个拟南芥基因亲缘关系较近,可归为3类,这与前人利用其他植物进行研究的结果一致。另外,苦荞8个可能定位于内质网的FtSUCs蛋白序列一致性达到34.30%,保守位点达到20个,表明植物SUCs蛋白序列在进化过程中相对保守。不同类的SUCs蛋白在蔗糖亲和力及功能方面存在差异,其中,Type Ⅰ即SUC1类的蔗糖亲和力较强,Type Ⅱ即SUC4类的蔗糖亲和力普遍较弱,而Type Ⅲ即SUC2类的蔗糖的亲和力普遍非常低,可能不行使蔗糖转运的功能。本研究共鉴定到4个属于Type I的苦荞FtSUCs,推测它们可能在蔗糖运输过程中发挥重要作用。
植物SUCs介导的蔗糖运输在植物生长发育以及逆境胁迫调控中发挥重要作用。例如,拟南芥AtSUC2在韧皮部装载中发挥重要作用,拟南芥3受创伤诱导上调表达,而4则在干旱胁迫响应中发挥重要作用。基于转录组测序发现苦荞10个在幼苗发育过程中表达模式不一致,推测它们可能在不同发育时期发挥其功能。苦荞种子出苗5 d时,植株处于子叶期,糖类物质主要由种子向上运输,Type Ⅰ中0002608200.01和Type Ⅲ中0001943900.01、0001944100.01在苦荞种子出苗5 d高表达,但在后期表达量急剧降低,说明它们主要在子叶期发挥功能。苦荞种子出苗10 d,2片真叶形成,植株由子叶期向真叶期过渡;出苗后15 d,植株则完全进入真叶期。Type Ⅱ中0000342600.01表达量在出苗后5~15 d间呈逐渐增加的趋势,说明其可能在出苗后逐步发挥功能。另外,Type Ⅰ中0001943500.01和0009584000.01在不同发育时期稳定表达,说明它们在整个发育过程中持续发挥功能。相关性分析证实,苗期苦荞中多个基因间存在显著正相关与显著负相关关系,暗示其可能在幼苗发育过程中存在协同效应或功能互补,进而实现对蔗糖运输的调控。
