菜心和芥蓝种间杂交创制异源四倍体蔬菜种质
2022-08-27吕凤仙和江明李崇娟胡靖锋杨红丽徐学忠张丽琴
吕凤仙,和江明,李崇娟,杨 鼎,胡靖锋,*,杨红丽,兰 梅,徐学忠,张丽琴
(1.云南省农业科学院 园艺作物研究所,云南 昆明 650205; 2.云南大学 农学院,云南 昆明 650504; 3.国家蔬菜改良中心云南分中心,云南 昆明 650205)
菜心(L. sspvar.Tsen. et Lee, AA, 2=20)和芥蓝(var., CC, 2=18)均为十字花科芸薹属中以花薹为产品的蔬菜,由于营养丰富、风味独特,广受消费者的喜爱。由于菜心和芥蓝在生产上推广应用的多为地方常规品种,存在产量低、品质差、病虫害严重等问题,需要选育商品性和品质更好的品种。但菜心和芥蓝现有遗传资源背景狭窄,种质资源不够丰富,仅通过常规的种内杂交很难在现有基础上取得突破性进展,选育的推广品种抗逆性和品质都不能很好地满足市场和生产的需求。通过菜心和芥蓝的种间杂交,可以创造综合亲本优势的新物种、新材料,是解决基因资源匮乏的重要手段。但远缘杂交胚乳的败育导致胚的最终死亡,以及远缘杂交受精后的不亲和性,杂交后无法结种,自然生长无法得到后代,需要通过人工胚挽救等措施才能得到种间杂种。梁红等、乔海云通过芸薹属间的远缘杂交,创造了新的作物种类,丰富了芸薹属的遗传背景。满红、魏云晓等通过种间杂交结合胚挽救的方法,创建了菜薹和芥蓝的种间异源四倍体和异源三倍体种质,但作为重要的薹用蔬菜,有关菜心和芥蓝的种间杂种创制新型薹用蔬菜的系统研究还不多。本研究以菜心作为母本,芥蓝作为父本,在种间杂交后进行人工胚挽救,并筛选适合的胚挽救方式、增殖扩繁方式、染色体加倍方式,以克服种间杂交不亲和性,提高种间杂种获得率,创制异源四倍体蔬菜种质,以期获得综合两亲本优良性状、具有良好商品性和生产价值的新薹用蔬菜,为后续菜心和芥蓝的育种过程提供材料。
1 材料与方法
1.1 材料
供试的菜心编号CX001,芥蓝编号JL001,均为云南省农业科学院园艺作物研究所十字花科课题组多年选育材料。
1.2 试验方法
1.2.1 种间杂交
于2020年6月16日播种,2020年7月8日定植于云南省农业科学院嵩明实验基地塑料大棚内,进行常规栽培管理。菜心于2020年7月20日开始抽薹,芥蓝于2020年7月27日开始抽薹。在初花期,杂交前2 d摘除父本花序上已开放花朵套袋隔离。于母本开花当天,去除母本植株上已开放和已露柱头花蕾,对未开放花蕾进行剥蕾去雄,授以父本花粉,授粉后套袋隔离,严防父本以外的花粉污染。
1.2.2 培养基
杂交后第13天,饱满胚珠数量较多,观察种荚生长情况。选取表面有明显凸起的种荚,于超净工作台内用75%乙醇浸泡50 s,将乙醇倒出后用10%次氯酸钠浸泡15 min,倒出次氯酸钠后用无菌水冲洗3次,最后用无菌水浸泡3 min。用解剖刀沿子房腹缝线和背缝线处轻轻切开,用镊子挑取饱满且呈绿色的胚珠,接种到不同成分的培养基(表1)上,在(25±2) ℃、光周期12 h/12 h(L/D)、光照强度1 500 lx的条件下培养,培养30 d,统计不同培养基的出胚率,筛选最适宜的培养基。
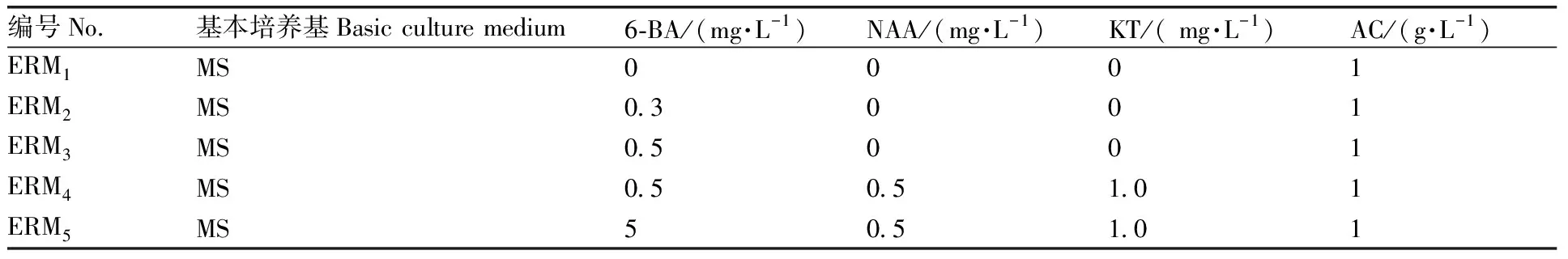
表1 菜心和芥蓝种间杂种胚挽救培养基成分
1.2.3 取材时间
分别于大田授粉后10、11、12、13、14、15 d,选取表面有明显凸起的种荚,用1.2.2节方法消毒后,挑取饱满且绿的胚珠,接种到ERM培养基上,在1.2.2节条件下培养,接种一周后观察是否发生污染,30 d后统计出胚率,观察不同取材时间对出胚率的影响。
1.2.4 生根培养基
将成活并成功分化的胚珠转移至增殖培养基DM(MS+0.9 mg·L6-BA+0.1 mg·LNAA)上进行增殖分化,具有完整植株形态后转接至1/2MS培养基上继续生长,至植株具3对真叶后,转移至MS或1/2MS添加不同生长素的RM、RM、RM、RM培养基,具体成分如表2所示,统计各培养基生根率和生根时间,筛选最适宜的生根培养基。
1.2.5 人工诱导染色体加倍
通过增殖分化培养基扩繁后的植株,使用秋水仙碱处理进行染色体加倍,处理方法分为3种。A处理:3对真叶幼苗转接至含50~90 mg·L秋水仙碱的1/2MS培养基,在培养箱内培养8 d,转接至不含秋水仙碱的生根培养基,生根后定植。B处理:3对真叶幼苗转接至含50~90 mg·L秋水仙碱的生根培养基,生根后定植。C处理:3对真叶幼苗转接至不含秋水仙碱的生根培养基,生根后炼苗,在成活植株抽薹前,将0.2%秋水仙碱溶液滴于植株顶芽和叶腋之间(每日上午1次,连续5 d)。
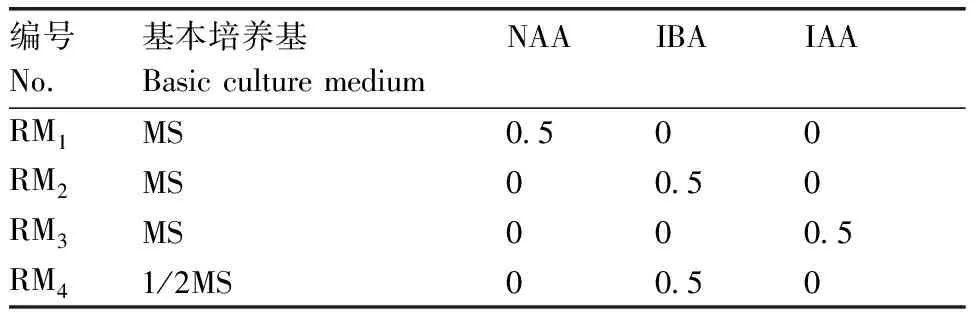
表2 菜心和芥蓝种间杂种生根培养基成分
处理后的植株,在田间常规管理,开花后取新鲜花朵,去除花瓣和雌蕊,将花粉置于干净载玻片上,滴加2~3滴亚历山大染色液(Alexander Stain),充分混合后盖上干净盖玻片,染色5~10 h后在显微镜下观察。
1.2.6 种间杂种F代鉴定
形态学鉴定:在F代杂种植株繁茂时观察,记录其叶形、叶色、叶表面特征、花器官的颜色大小、植株高度和开展度,以及生长习性,对经秋水仙素处理植株和未处理植株进行比较,并分别与菜心和芥蓝两亲本进行比较。
花药染色体观察:经人工染色体加倍处理后的植株现蕾后,取植株未开放花序,选取直径≤2 mm的花蕾,用新鲜卡诺固定液(乙醇∶醋酸体积比3∶1)固定过夜,后视固定液颜色每6 h更换一次固定液,直至花蕾颜色泛白且固定液无色,剥取花蕾取其中1个花药,参照杨汉等的方法进行染色体制片,卡宝品红染色液染色后,用电子显微镜观察并统计花药染色体数量。
雌蕊染色体观察:选取植株未开放花序上直径2~3 mm花蕾,用解剖针取出完整雌蕊,置于0.002 mol·L8-羟基喹啉中,4 ℃冰箱中暗处理3 h,转入卡诺固定液中固定过夜,取1个雌蕊置于1 mol·L盐酸中,60 ℃水浴7.5 min后,蒸馏水浸泡材料1 min,在载玻片上捣碎材料后用卡宝品红染色30 s,压片后在电子显微镜下观察并统计雌蕊染色体数量。分别将花药和雌蕊染色体数量与亲本染色体数量进行比较。
倍性鉴定:参照Yang等的解离液、染液配方和实验步骤,采用流式细胞仪对种间杂种F进行倍性分析。以菜心、芥蓝和甘蓝型油菜为参照样本,取F植株、亲本、甘蓝型油菜新鲜幼嫩叶片25 mg左右,加入400 μL预冷的“Aru” 缓冲液,用锋利刀片快速切碎至没有明显大块,经400目过滤器过滤至5 mL管中,加入2.5 μL RNase A(20 mg·mL)和5 μL PI染液(10 mg·mL),充分混匀后于冰上避光孵化30 min,移至上样管上机检测,收集5 000~10 000个颗粒,观察样本主峰位置,分析样本倍性。
1.3 数据分析
利用SPSS23.0软件对试验数据进行差异性显著分析。
2 结果与分析
2.1 培养基成分对出胚的影响
如表3所示,接种在ERM培养基上的胚珠均未成功分化,出胚率为0,其余4种培养基的出胚率均不同,ERM培养基出胚率为10.47%,显著高于其他培养基。因此,ERM(MS+0.5 mg·L6-BA+0.5 mg·LNA+1.0 mg·LKT+1 g·LAC)为菜心和芥蓝种间杂种的最佳胚挽救培养基。
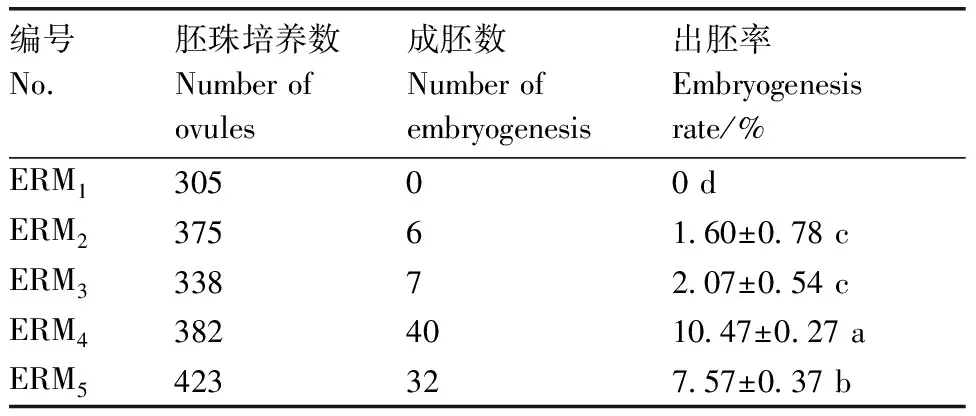
表3 菜心×芥蓝杂种胚珠在不同培养基上出胚率
2.2 不同取材时间对出胚的影响
如表4所示,不同取材时间的胚珠,接种后出胚率均存在差异,授粉后10~12 d取材,出胚率逐渐增大,授粉后第12天取材出胚率达到最高,为26.32%,且显著高于其他取材时间。因此,菜心和芥蓝种间杂种胚挽救的最佳取材时间为授粉后第12天。
2.3 不同生根培养基对生根效果的影响
从表5可知,基本培养基添加不同种类和浓度的生长素,生根效果均存在差异。统计数据显示,RM与RM生根率最高,分别为36.00%、37.21%,二者无显著差异,但显著高于其他培养基。生根时间最短的为RM,平均生根时间为8.33 d,与RM平均生根时间8.67 d无显著差异,显著短于其他培养基。
将幼苗转接至以MS培养基为基本培养基的RM、RM、RM时,部分幼苗出现盐害现象,叶片部分黄化且逐渐脱落(图1-A、B、C),转接至以1/2MS培养基为基本培养基的RM,幼苗盐害现象得到缓解(图1-D)。综上所述,RM(1/2MS+0.5 mg·LIBA)生根率最高、生根时间最短,幼苗生长状况良好,为菜心和芥蓝种间杂种的最佳生根培养基。
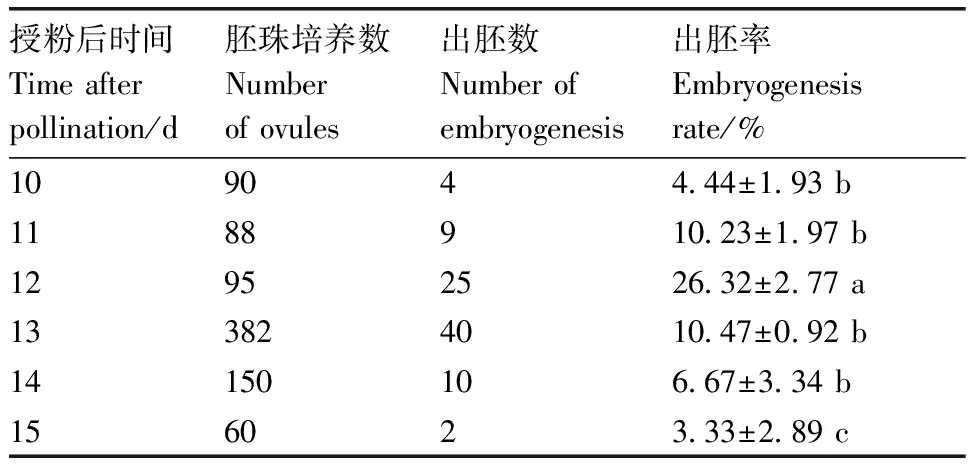
表4 菜心×芥蓝不同取材时间出胚率
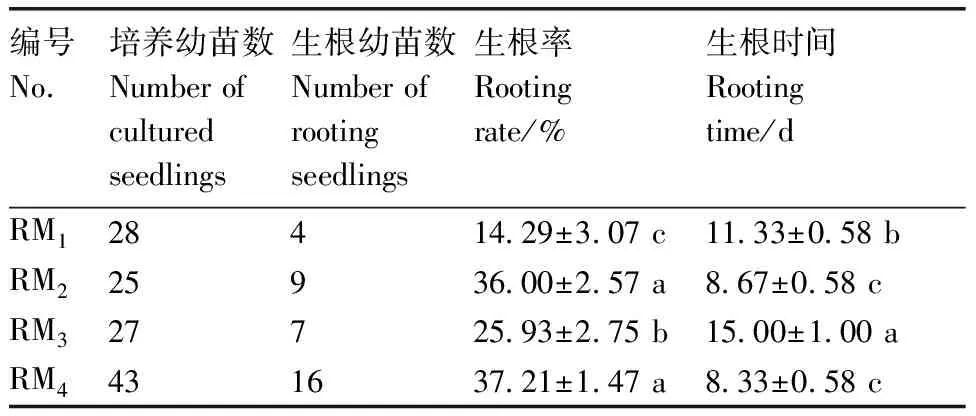
表5 菜心×芥蓝种间杂种不同培养基生根情况

A,RM1上发生盐害现象的幼苗;B,RM2上发生盐害现象的幼苗;C,RM3上发生盐害现象的幼苗;D,RM4上正常生长幼苗。标尺=1 cm。A, Seedlings with salt damage on RM1; B, Seedlings with salt damage on RM2; C, Seedlings with salt damage on RM3; D, Normal growth of seedlings on RM4; Bar scale=1 cm.图1 菜心×芥蓝种间杂种幼苗生根过程生长情况Fig.1 Growth during rooting of Brassica rapa L. ssp chinensis var. utilis Tsen. et Lee×Brassica oleracea var. Alboglabra interspecific hybrid seedlings
2.4 秋水仙素处理诱导染色体加倍效果
不同染色体加倍处理效果如表6所示,A、B处理幼苗受秋水仙碱影响,生根率较低,大部分苗会出现叶片枯黄脱落、生长点死亡至整株幼苗死亡(图2);即使正常生根,在炼苗阶段会由于根系发育不良,死苗率较高,炼苗后部分幼苗会出现根系褐化,逐渐腐烂,叶片枯萎脱落,幼苗死亡现象(图3)。C处理因处理时间相对偏后,秋水仙碱对幼苗抑制效果较小,成活率较高,但诱导所得可育植株均为嵌合体,即出现同一植株部分侧枝、同一侧枝的部分花序可育,部分侧枝、部分花序不可育的现象;经花粉育性观察结果显示,可育的花粉粒经染色后花粉壁呈绿色,原生质呈红色(图4-A),花粉具有活性,不可育花粉粒呈绿色(图4-B),不具有活性,还需要扩繁至F代才能用于蔬菜育种试验。但统计结果显示,C处理的诱导加倍率可达62.26%,为最佳的染色体诱导加倍处理方式。

表6 菜心×芥蓝种间杂种染色体加倍方法处理效果
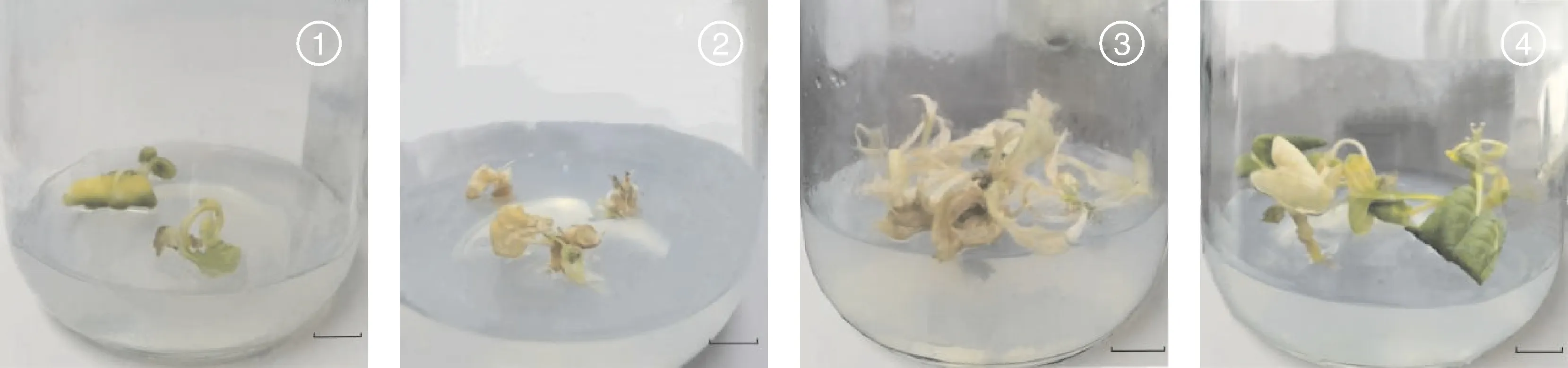
1和2,A处理;3和4,B处理。标尺=1 cm。1 and 2, Treatment A; 3 and 4, Treatment B. Bar scale=1 cm.图2 菜心×芥蓝种间杂种染色体加倍过程死亡现象Fig.2 Death of seeldings during the chromosome doubling process of Brassica rapa L. ssp chinensis var. utilis Tsen. et Lee×Brassica oleracea var. Alboglabra interspecific hybrids
2.5 杂种后代表现
2.5.1 形态学表现
菜心-芥蓝种间杂种F代植株各性状皆介于亲本之间(图5),基生叶形状为长卵圆,叶色为深绿色,主薹形状均与菜心相似,但主薹颜色为浅绿色,叶片和主薹表面都有微量蜡粉,更接近芥蓝。此外,母本菜心所开的花为黄色,杂种植株所开花朵颜色与父本芥蓝相同,为白色。综合各性状判断,杂种植株为菜心×芥蓝种间杂交真杂种。经染色体加倍后可育的植株与不育植株相比,花器官和花瓣均较大,可育株的雄蕊饱满且有花粉,不育株雄蕊干瘪无花粉(图6)。可育株的角果较长,表面有明显膨大,内部有饱满胚珠;不育株的角果较短,表面无膨大,内部无胚珠。
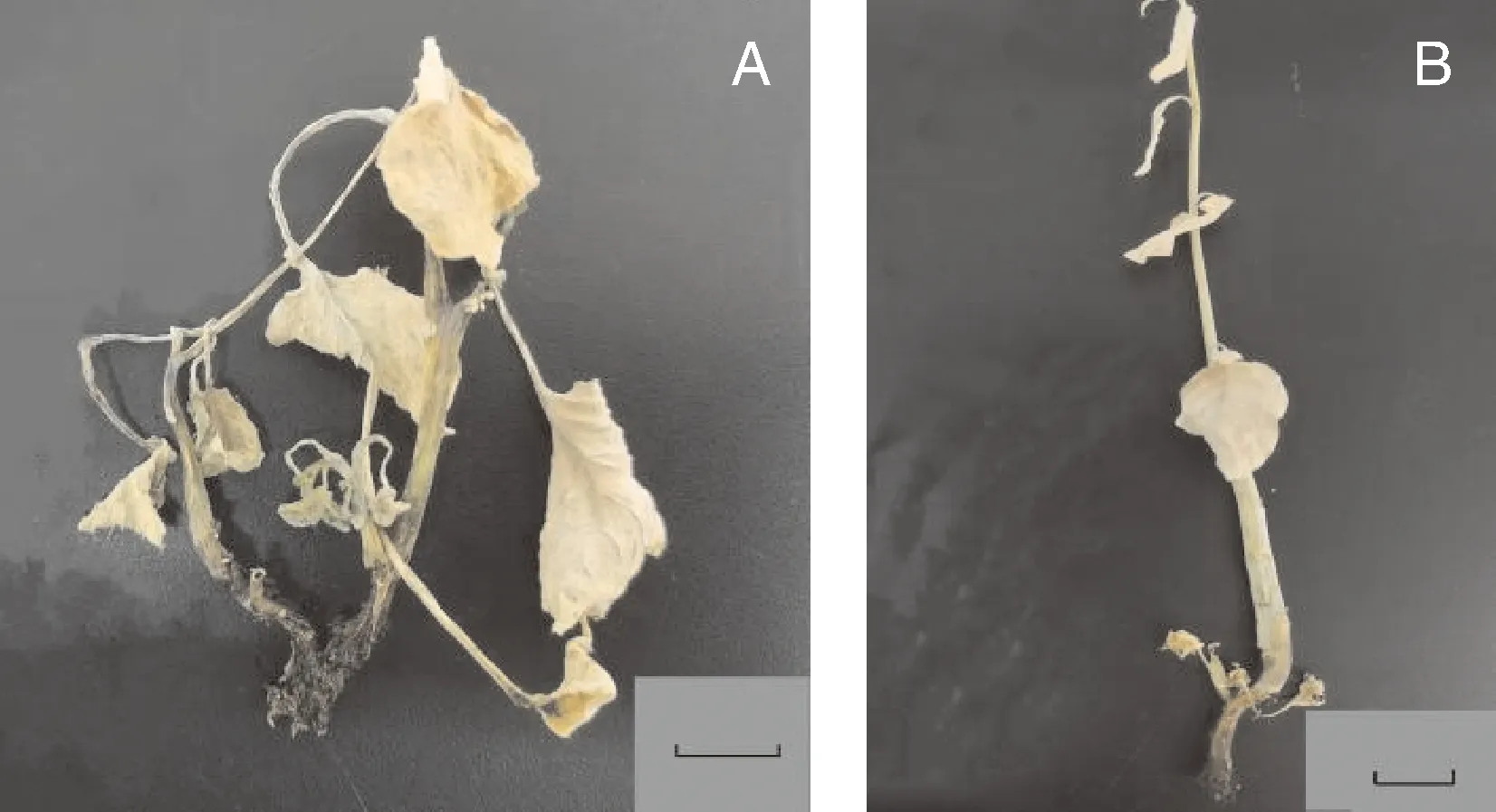
A,A处理;B,B处理。标尺=1 cm。A, Treatment A; B, Treatment B. Bar scale=1 cm.图3 菜心×芥蓝种间杂种经染色体加倍处理后幼苗炼苗后死亡现象Fig.3 Death of seeldings with chromosome doubling treatment after the seedling hardening of Brassica rapa L. ssp chinensis var. utilis Tsen. et Lee× Brassica oleracea var. Alboglabra interspecific hybrids
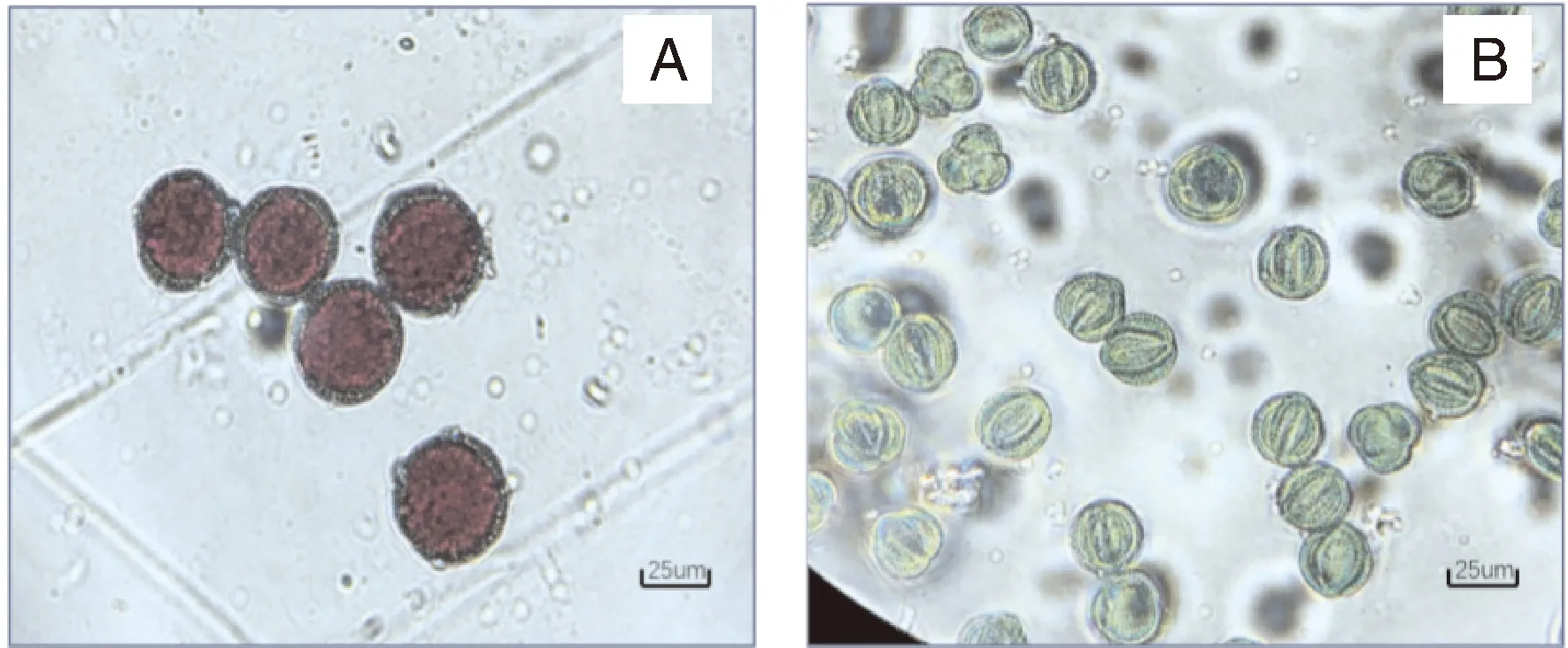
A,可育株花粉; B,不育株花粉;标尺=25 μm。A, Pollen of fertile plant; B, Pollen of sterile plant. Bar scale=25 μm.图4 菜心×芥蓝种间杂种经染色体加倍处理后花粉染色观察结果Fig.4 Observation of pollen staining of interspecific hybrids of Brassica rapa L. ssp chinensis var. utilis Tsen. et Lee×Brassica oleracea var. Alboglabra after chromosome doubling treatment
2.5.2 染色体分析
对菜心×芥蓝种间杂交胚挽救获得的F代植株进行花药和柱头染色体数量鉴定,结果表明,不育的植株柱头染色体数量为19条(图7-A),与预期染色体数量相符,且A组染色体和C组染色体分界明显,为异源二倍体。经秋水仙碱处理花粉可育的植株,柱头染色体数量为38条(图7-B),花药染色体数量为19条(图7-C),为亲本菜心(2=20)和芥蓝(2=18)的染色体数量之和,进一步证明所获得杂种为种间杂交真杂种,且为异源四倍体。
2.5.3 倍性鉴定
以亲本菜心和芥蓝,以及甘蓝型油菜作为参照样本,对可育和不育的种间杂种F代植株进行倍性检测,结果显示,不育植株(图8-D)荧光强度最高峰与亲本菜心(图8-A)的荧光强度最高峰处于同一位置,结合染色体分析结果判断不育植株为异源二倍体;可育植株(图8-E、8-F)荧光强度最高峰位置与甘蓝型油菜(图8-C)的荧光强度最高峰位置相同,为异源四倍体,倍性检测结果与染色体数量鉴定结果一致。
3 结论与讨论

A和D,菜心;B和E,种间杂种;C和F,芥蓝;A、B、C的标尺=5 cm;D、E、F标尺=0.5 cm。A and D, Brassica rapa L. ssp chinensis var. utilis Tsen. et Lee; B and E, Einterspecific hybrid; C and F, Brassica oleracea var. Alboglabra; Bar scales of figure A, B and C were 5 cm; Bar scales of figure D, E and F were 0.5 cm.图5 菜心×芥蓝种间杂种植株形态Fig.5 Plant morphology of Brassica rapa L. ssp chinensis var. utilis Tsen. et Lee×Brassica oleracea var. Alboglabra interspecific hybrids

A、D、E、G、I为可育株;B、C、F、H、J为不育株;标尺=1 cm。A, D, E, G and I were fertile plants; B, C, F, H and J were sterile plant. Bar scale=1 cm.图6 菜心×芥蓝种间杂种花朵和角果形态Fig.6 Flower and pod forms of Brassica rapa L. ssp chinensis var. utilis Tsen. et Lee×Brassica oleracea var. Alboglabra interspecific hybrids
邱德勃等研究芸薹属植物远缘杂交发现,胚乳的败育导致胚的败育,最终无法获得种子,远缘杂交的发展受到制约。戴林建研究发现,远缘杂交的不亲和性主要受杂交组合方式和亲本基因型的影响,且杂交亲本亲缘关系越近,越容易取得成功。本试验选择了芸薹属中A和C组染色体进行种间杂交,相较于B和C、A和B组染色体,A和C组染色体亲缘关系较近,更容易克服不亲和性。李明山等研究表明,适宜的种间杂种幼胚离体培养方式,可有效克服远缘杂交的不亲和性。Harberd研究表明,杂交胚珠的培养受到胚龄大小和培养基成分等多种因素的影响。满红对四倍体菜心和四倍体芥蓝种间杂种进行子房培养,得出的最佳取材时间为授粉后7 d;谭亚飞对白菜型油菜黄籽沙逊与芥蓝种间杂种胚珠进行培养,得出的最佳取材时间为授粉后11 d,最佳培养基为MS+2.0 mg·L6-BA+0.2 mg·LNAA+1.0 mg·LAC。受幼胚培养方式和母本基因型的影响,本试验最佳胚挽救培养基为MS+0.5 mg·L6-BA+0.5 mg·LNAA+1.0 mg·LKT+1g·LAC,出胚率为10.47%;最佳取材时间为授粉后第12天,出胚率为26.32%。

A,不育株柱头染色体;B,可育株柱头染色体;C,可育株花药染色体。A, Stigma chromosome of sterile plant; B, Stigma chromosome of fertile plant; C, Anther chromosomes of fertile plants.图7 菜心×芥蓝种间杂种染色体数量Fig.7 Chromosome number of Brassica rapa L. ssp chinensis var. utilis Tsen. et Lee×Brassica oleracea var. Alboglabra interspecific hybrids
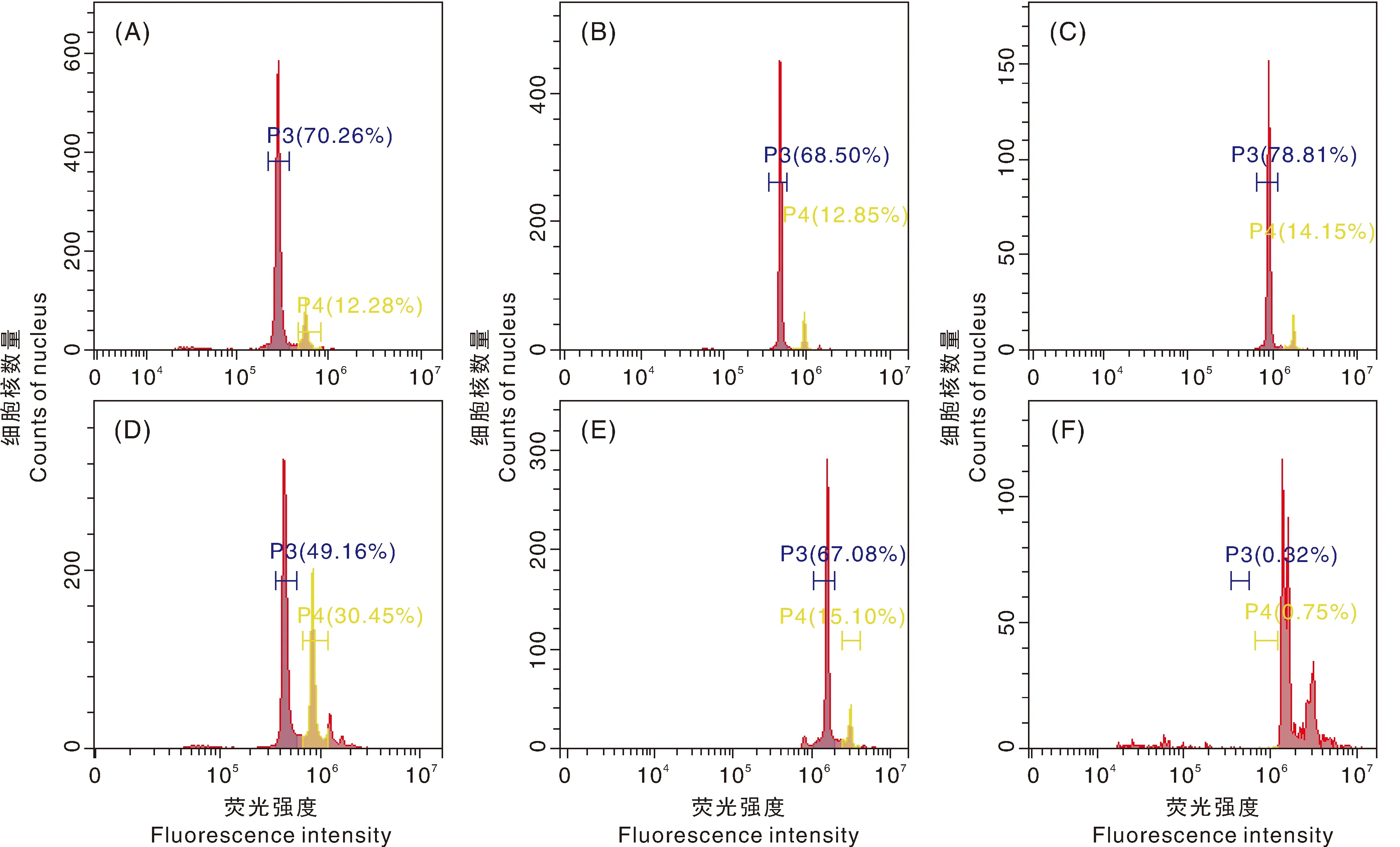
A,菜心;B,芥蓝C,甘蓝型油菜;D,不育株;E和F,可育株。A, Brassica rapa L. ssp chinensis var. utilis Tsen. et Lee; B, Brassica oleracea var. Alboglabra; C, Brassica napus L.; D, Sterile plant; E and F, Fertile plant.图8 菜心×芥蓝种间杂种倍性鉴定直方图Fig.8 Histogram of ploidy identification of Brassica rapa L. ssp chinensis var. utilis Tsen. et Lee×Brassica oleracea var. Alboglabra interspecific hybrids
对胚珠培养后获得的幼苗进行快速增殖扩繁,是获得菜心和芥蓝种间杂种的重要阶段,胚培苗的生根过程是扩繁的重要环节。不同植物诱导生根的适宜生长调节剂种类不同,本试验中添加不同生长素的培养基生根率和生根时间均有差异,1/2MS+0.5 mg·LIBA培养基最适宜生根。石淑敏等在诱导甘蓝型油菜小孢子再生苗染色体加倍的研究中,利用培养基内添加秋水仙碱溶液的方法,小苗成活率低且操作复杂,利用秋水仙碱溶液涂抹生长点2次的方法,加倍率极低。本试验中将单倍体苗接种于含秋水仙碱溶液的培养基中,也存在成活率极低的问题;但将秋水仙碱涂抹次数增加至5次时,可将染色体加倍率提高至62.26%,加倍效果佳。
江莹芬等对埃塞俄比亚芥和白菜型油菜远缘杂种进行鉴定,认为形态学鉴定操作简单易行,但还需结合细胞学等方法确保结果准确。本研究结果显示,黄花菜心和白花芥蓝杂交所得后代均开白花。卢长明等统计了人工合成甘蓝型油菜,结果也显示,白花对黄花基因具有上位性。黄花菜心和白花芥蓝杂交所得后代叶形、叶色、主薹形状等接近母本,主薹颜色接近父本,初步确定为真杂种;观察花器官大小、花药,以及是否正常结籽,区分出可育株和不育株,判断染色体加倍是否成功。同时利用染色体观察和流式细胞仪倍性鉴定对形态学鉴定结果进行检验,鉴定结果均显示可育株为异源四倍体,不育株为异源二倍体。
本试验创制了菜心和芥蓝种间杂交的异源四倍体种质,但由于人工染色体加倍处理,秋水仙碱溶液对植株毒性大,影响植株正常生长。后续研究将扩繁至F代,进一步观察田间表现,并对可溶性糖和维生素等营养物质含量进行研究,分析后代作为薹用蔬菜的应用价值。
