白菜bZIP转录因子基因家族应答春化反应关键基因表达分析
2022-07-16马李广张贺龙庞小可王浩李广汪承刚袁凌云侯金锋唐小燕陈国户
马李广 张贺龙 庞小可 王浩 李广 汪承刚 袁凌云 侯金锋 唐小燕 陈国户





摘要: 为研究白菜( Brassica rapa )碱性亮氨酸拉链(Basic leucine zipper,bZIP)转录因子家族基因( BrbZIP )的相关功能,通过生物信息学分析技术,鉴定全基因组的 BrbZIP 基因家族成员,并对其染色体分布、进化关系、表达模式及其应答春化反应等进行了分析。白菜bZIP转录因子基因家族共有118个成员,在染色体上不均等分布。白菜组织转录组分析结果显示,大部分 BrbZIP 基因在根、茎、叶、花及荚中均有较高的表达丰度,且具有组织表达特异性;春化反应转录组、荧光定量PCR及相关基因互作网络分析结果表明,白菜 bZIP 基因家族中应答春化反应相关基因上调与下调表达基因数量差异不大,相互之间存在复杂的互作网络,其中4个成员(Bra039631、Bra020620、Bra004550与Bra020471)是应答春化反应的中心节点。
关键词: 白菜; bZIP转录因子; 基因家族; 春化反应; 生物信息学分析
中图分类号: S634.3 文献标识码: A 文章编号: 1000-4440(2022)03-0765-10
Genome-wide identification of bZIP transcription factor gene family in Brassica rapa and its association with vernalization
MA Li-guang 1 , ZHANG He-long 1 , PANG Xiao-ke 1 , WANG Hao 1 , LI Guang 1,2 , WANG Cheng-gang 1,2,3 ,YUAN Ling-yun 1,2 , HOU Jin-feng 1,2 , TANG Xiao-yan 1,2 , CHEN Guo-hu 1,2,3
(1.Anhui Provincial Engineering Laboratory of Horticultural Crop Breeding, School of Horticulture, Anhui Agricultural University, Hefei 230036, China; 2.Wanjiang Vegetable Industrial Technology Institute, Maanshan 238200, China; 3.Anhui Southwest Comprehensive Experimental Station of Anhui Agricultural University, Huaining 246100, China)
Abstract: In order to explore the function of basic leucine zipper (bZIP) transcription factor in Brassica rapa and its association with vernalization, bioinformation methods were used to identify the B. rapa bZIP gene family members. The chromosome location, evolutionary relationship, expression pattern and vernalization response were analyzed through a variety of biological information softwares. The results revealed that 118 BrbZIP transcription factors were unevenly distributed on the chromosomes of B. rapa . Transcriptome analysis showed that most of the BrbZIP genes had high expression levels in roots, stems, leaves, flowers and pods, and had tissue expression specificity. The results of transcriptome, fluorescence quantitative PCR and interaction network analysis suggested that there was n o significant difference in the number of up-regulated an d down-regulated genes responding to vernalization in bZIP gene family of B. rapa , and the interaction network was complex. Four BrbZIP genes were the central nodes in response to vernalization.
Key words: Brassica rapa ; bZIP transcription factor; gene family; vernalization; bioinformatics analysis
轉录因子(Transcription factors,TFs),是一类通过与下游基因启动子顺式元件特异性结合或与其他蛋白质互作来调控基因表达的一类蛋白质。在只存在于真核生物中的转录因子家族中,碱性亮氨酸拉链(Basic leucine zipper,bZIP)家族是转录因子中成员数量最多、最保守的家族之一,在植物生长发育和应答胁迫反应等生物过程中起着重要的调控作用 [1] 。
bZIP蛋白最显著的特征是含有一个保守的60~ 80个氨基酸组成的bZIP结构域( N -×7-R/K-×9- L -×6- L ),包括1个碱性氨基酸区域和1个相邻的亮氨酸拉链区;碱性氨基酸区域位于C端,由16~20个氨基酸残基组成 N -×7-R/K基序,负责与DNA启动子区域内特定序列结合;亮氨酸(Leu)拉链区位于 N 端,由亮氨酸或异亮氨酸(Ile)、缬氨酸(Val)组成的七肽重复序列(×9- L -×6- L -×6- L ),主要作用为bZIP蛋白特定识别和形成同源二聚体 [2] 。植物bZIP蛋白优先与含有ACGT核心元件的DNA序列结合,特别是G-box(CACGTG)、C-box(GACGTC)以及A-box(TACGTA)等元件 [3] 。
目前bZIP转录因子基因家族已在多个植物基因组中被鉴定出来,如拟南芥( Arabidopsis thaliana ) [4] 、油菜( B.napus ) [5] 、麻疯树( Jatropha curcas ) [6] 、藜麦( Chenopodium quinoa ) [7] 、杨树( Populus tremula ) [8] 、猕猴桃( Actinidia chinensis ) [9] 等。大量研究结果表明,bZIP转录因子在植物光信号、花的诱导与发育、种子成熟与萌发、非生物和生物胁迫等多个关键生物学过程中起着重要的作用。例如,在拟南芥中,bZIP11基因可影响根系发育 [10] ,HY5基因参与调控根和下胚轴的发育 [11] ;而bZIP1基因与花药发育过程中的水分运动密切相关,其过量表达不利于花粉发育 [12] 。小麦( Triticum aestivum )bZIP转录因子 TaGBF 基因调控 CO 及 FT 等基因的表达,影响植物开花 [13] 。大豆( Glycine max )植株中FDL19蛋白与FT2a、FT5a相互作用,可影响大豆的开花时间 [14-15] 。风疹卷尾草( Capsella rubella )TGA4基因的过表达,可延迟拟南芥开花时间 [16] 。编码bZIP转录因子的HY5基因,在植物幼苗光形态建成期间,对激活光反应基因起着重要的作用 [17] 。水稻( Oryza sativa )应激激活蛋白激酶SAPK100磷酸化OsbZIP77,导致开花时间提前 [18] ;OsFD4基因通过作用于茎尖分生组织中成花整合因子复合物的形成,促进水稻开花 [19] 。因此,bZIP转录因子在植物开花过程中具有重要的作用。
目前,bZIP转录因子在植物开花过程中的功能研究,已有一些进展。然而迄今为止,未有白菜bZIP转录因子参与春化反应的研究报道。本研究擬对白菜( B. rapa ) bZIP 基因家族进行系统研究,包括基因家族成员鉴定、蛋白质氨基酸序列分析和系统发育关系、基因的染色体分布、基因组串联复制和片段复制、基因共线性分析等,此外,基于转录组数据及荧光定量PCR分析,探讨白菜 bZIP 基因在不同组织中的表达模式及应答春化反应相关基因的表达,为进一步解析白菜 bZIP 基因在春化过程中的功能奠定基础。
1 材料与方法
1.1 数据来源
从白菜基因组数据库(BRAD;http://brassicadb.cn/)下载白菜全基因组数据(V1.5)及其注释文件,从NCBI Sequence Read Archive(SRA)数据库下载白菜不同组织(根、茎、叶、花、荚等) [20] 及春化反应(JWW材料春化0 d、25 d、35 d;XBJ材料春化0 d、10 d、25 d) [21] RNA-Seq原始数据。RNA-Seq原始数据经过质量控制过滤后,比对至白菜基因组(V1.5)序列,再利用FeatureCounts进行有参转录组定量。基因表达丰度采用FPKM值进行计算,差异表达基因采用DESeq软件包进行计算 [22-23] 。
1.2 白菜 bZIP 基因家族成员的鉴定
从白菜基因组数据库(BRAD)下载 bZIP 基因家族候选成员ID,利用TBtools软件 [24] 提取白菜 bZIP 基因家族候选基因序列、蛋白质氨基酸序列等信息。使用Pfam(http://pfam.xfam.org/)、SwissProt(https://www.expasy.org/resources/uniprotkb-swiss-prot)、SMART(http://smart.embl-heidelberg.de/)和InterProScan(https://www.ebi.ac.uk/interpro/about/interproscan/)等数据库鉴定所有候选蛋白质氨基酸序列。对确定BrbZIP成员的蛋白质氨基酸序列,利用MEGA 7.0软件采用邻接法以及泊松校正与成对删除法,构建系统进化树 [25] 。
1.3 白菜bZIP家族理化性质及 bZIP 基因组信息分析
采用ExPasy工具(https://web.expasy.org/compute_pi/),分析白菜bZIP家族的相对分子质量、等电点和长度等理化性质。提取白菜 bZIP 基因家族注释信息,并定位到染色体上,利用TBtools工具进行可视化,获得 BrbZIP 基因家族成员染色体定位图谱。将白菜 bZIP 基因家族编码的蛋白质氨基酸序列进行all-BLASTP-all分析( top <5, E <1e -5 )。采用TBtools软件中MCScanX工具提取共线性基因对,再利用Circos工具进行可视化。
1.4 白菜 bZIP 基因家族组织表达分析及其应答春化反应分析
利用白菜不同组织转录组数据 [20] ,分析 BrbZIP 基因家族的组织表达特征。利用白菜春化反应材料转录组数据 [21] ,分析白菜 bZIP 基因家族应答春化反应。利用STRING数据库(https://string-db.org/)分析白菜应答春化相关 bZIP 基因互作网络关系。对 BrbZIP 基因家族中应答春化反应关键基因,利用荧光定量PCR技术分析其在JWW不同春化时间材料中的相对表达量 [21] 。
2 结果与分析
2.1 白菜 bZIP 基因家族鉴定及其编码的蛋白质理化特性分析
经筛选鉴定,在白菜基因组中共获得118个 bZIP 基因家族成员。理化性质分析发现,白菜 bZIP 基因家族开放阅读框(ORF)长度为273~ 1 893 bp,编码90~ 630个氨基酸(AA),预测相对分子质量大小在1.062× 10 4 至6.874× 10 4 之間,等电点(pI)范围为4.80~ 10.11。对白菜bZIP蛋白的等电点与相对分子质量之间进行比较分析,发现酸性与碱性BrbZIP蛋白数量相差不大,且相对分子质量分布较规律。对白菜bZIP蛋白进行亚细胞定位预测分析,结果显示绝大部分白菜bZIP蛋白家族成员定位于细胞核内,只有6个成员定位于细胞膜、线粒体或叶绿体中,表明白菜bZIP蛋白家族的功能主要发生在细胞核中。
2.2 白菜 bZIP 基因家族染色体定位分析
利用白菜基因组注释信息,将白菜 bZIP 基因家族定位于染色体上。结果如图1所示,118个白菜 bZIP 家族基因不均等分布在10条染色体上,各染色体上的基因数量与染色体大小无关。其中,染色体A09上的 BrbZIP 基因家族数量最多(23个),染色体A10上的 BrbZIP 基因家族数量最少(7个)。此外,白菜 bZIP 家族基因在A04(Bra016959与Bra016953)、A09(Bra007274与Bra007276)染色体上各有1对基因串联重复,表明白菜 bZIP 家族基因 在进化过程中发生了少量的串联重复现象。
2.3 白菜与拟南芥 bZIP 基因家族系统发育分析
为了确定白菜 bZIP 基因家族成员的进化关系,通过MEGA-X软件中MUSCLE工具将118个白菜 bZIP 基因与73个拟南芥 bZIP 基因进行序列分析比对,运用邻接法构建了系统发育树(图2)。白菜与拟南芥 bZIP 基因家族分成了5个大簇(GroupA~ GroupE)、11个小簇。其中,Group E的 bZIP 基因数量最多,包括21个 AtbZIP 基因和41个 BrbZIP 基因;Group C的数量最少,仅有16个,包括7个 AtbZIP 基因和9个 BrbZIP 基因。系统发育树分析结果表明,各簇中白菜和拟南芥 bZIP 基因具有较高的同源性。
2.4 白菜 bZIP 家族基因共线性分析
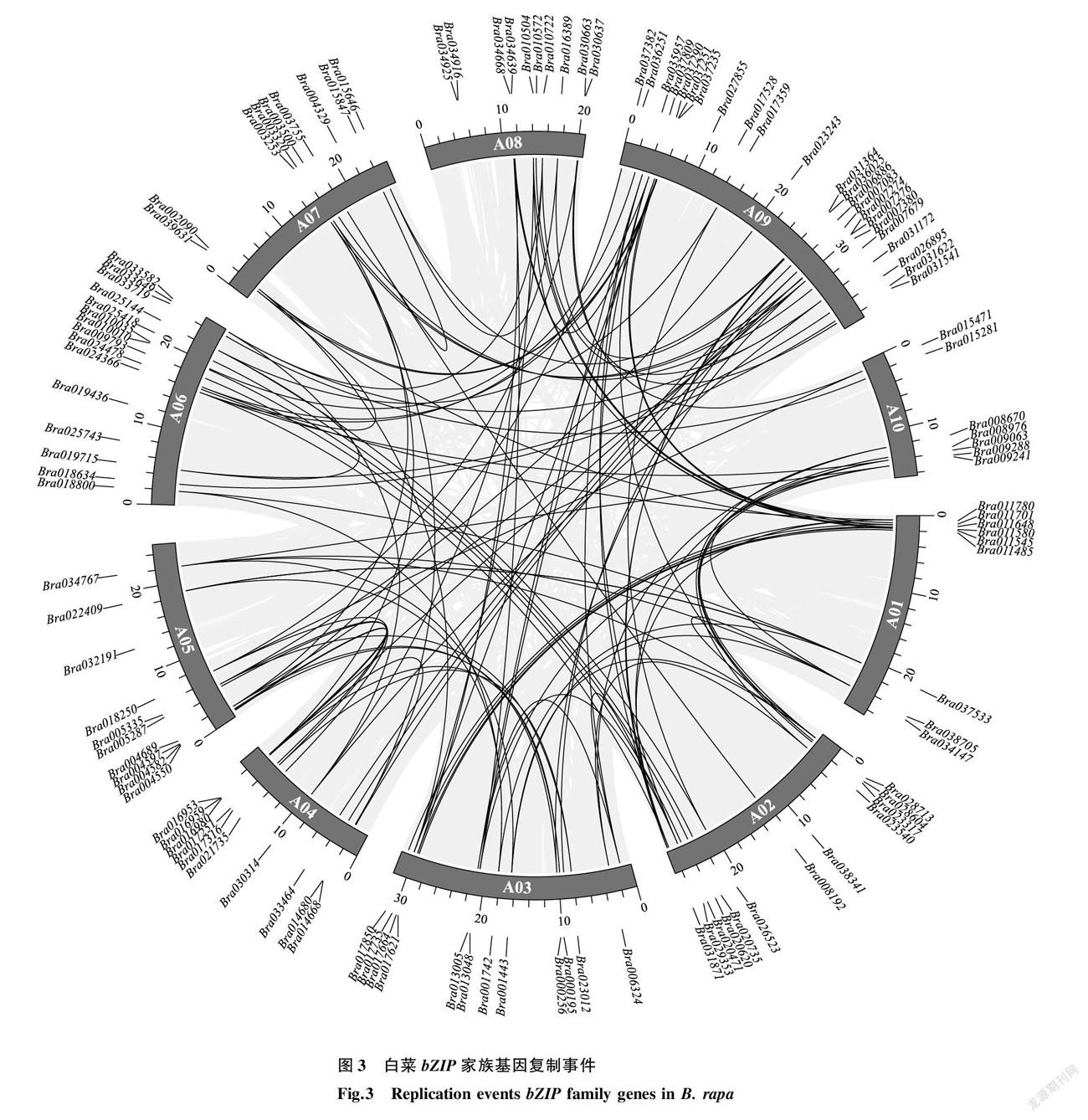
为了分析白菜 bZIP 家族基因的扩张模式,对其基因复制事件进行了分析。如图3所示,118个白菜 bZIP 家族基因成员之间,有110对存在共线性基因对,其中存在着单个基因与多个基因对应的情况,表明白菜 bZIP 家族在发育进化过程中发生了大量的基因复制现象,是白菜 bZIP 基因多样性的主要驱动力。
2.5 白菜 bZIP 家族基因表达模式及响应春化反应分析
为了研究白菜 bZIP 基因在不同组织中的表达模式,利用白菜组织转录组数据分析了 BrbZIP 基因在根、茎、叶、花、荚等组织中的表达情况。结果如图4A所示,大部分白菜 bZIP 基因在各组织中均有表达,表明这些 bZIP 基因在白菜整个生长发育阶段均有重要的作用。对 FPKM 值大于500的 BrbZIP 基因进行了维恩分析,发现各组织共有 BrbZIP 基因仅13个(图4B)。此外,少数 BrbZIP 基因具有较强的组织表达特异性,如Bra037809、Bra018634、Bra031871、Bra023540、Bra031622等主要在根系中表达,Bra010504、Bra004329等主要在茎中表达,Bra025144主要在花中表达,而Bra009288、Bra035957等在荚中表现出较高的转录丰度,表明 BrbZIP 基因的表达具有组织特异性(图4)。
我们还利用白菜春化反应转录组数据分析了应答春化的相关 BrbZIP 基因,结果如图5所示,在白菜JWW材料与XBJ材料春化转录组数据中,共筛选出42个差异表达 BrbZIP 基因(DEGs)(图5A),这些DEGs中,JWW材料与XBJ材料共有18个(图5B),其中8个DEGs上调表达,10个DEGs下调表达(图5C)。对白菜JWW材料与XBJ材料中春化相关 BrbZIP 基因分别进行表达趋势分析发现,共有5个显著富集趋势( P < 0.05)。在JWW材料中,18个 BrbZIP 基因随春化时间延长,转录丰度逐渐增大,其中12个 BrbZIP 基因在春化25 d时表达量最大(Profile 6)、6个 BrbZIP 基因在春化35 d时表达量最大(Profile 7);10个 BrbZIP 基因随春化时间延长转录丰度逐渐降低(Profile 1)。在XBJ材料中,16个 BrbZIP 基因随春化时间延长,转录丰度逐渐增大,在春化10 d时表达量最大(Profile 6);7个 BrbZIP 基因随春化时间延长转录丰度逐渐降低(Profile 0)。在JWW材料与XBJ材料共有模块6(Profile 6)中,共有9个 BrbZIP 基因(图6),表明这些 BrbZIP 基因在白菜春化反应中具有重要的作用。
2.6 白菜 bZIP 家族应答春化相关基因互作网络分析
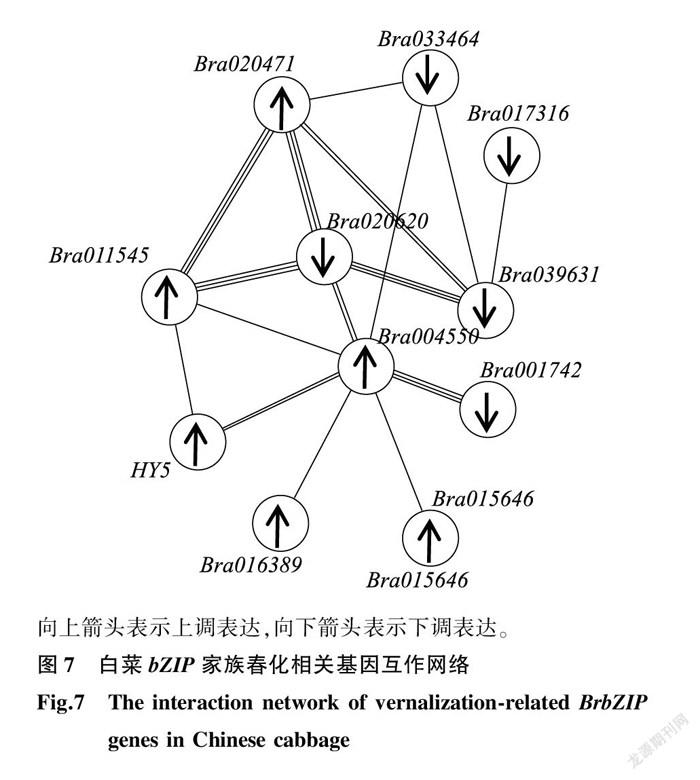
为了进一步研究白菜春化应答 BrbZIP 基因的相互作用,利用STRING数据库对JWW材料与XBJ材料中共有18个应答春化 BrbZIP 基因进行互作网络分析(图7)。结果显示18个 BrbZIP 基因中有11个与其他基因有相互作用关系,且互作网络表现出复杂的相关性,其中Bra039631(AtbZIP2)、Bra020620(AtbZIP63)、Bra004550(AtbZIP55)、Bra020471(AtbZIP9)基因在互作网络中是应答春化反应的中心节点,表明其在应答春化反应调控网络中具有重要的作用。
2.7 白菜 bZIP 家族应答春化相关基因荧光定量PCR分析
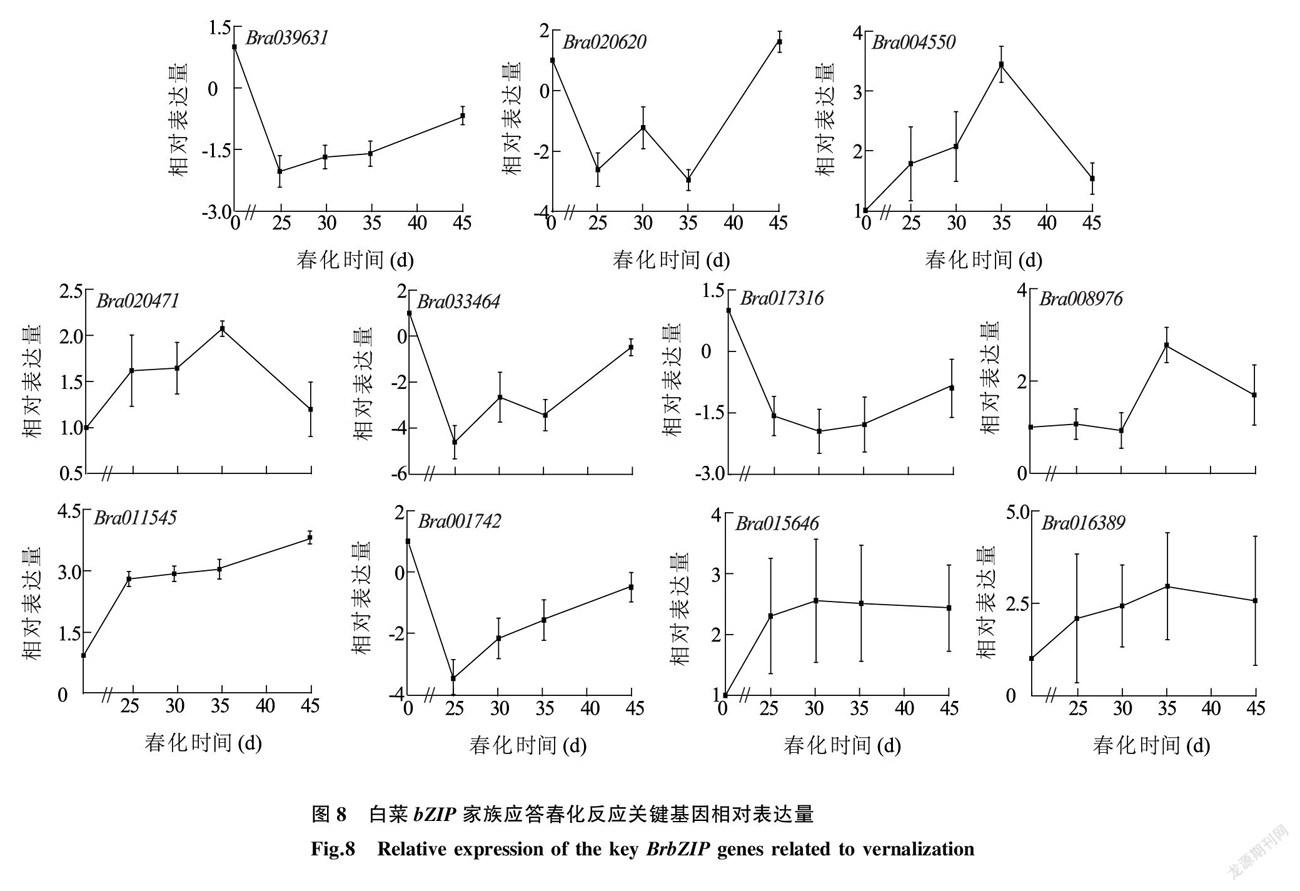
利用荧光定量PCR技术,进一步分析白菜 bZIP 家族应答春化反应关键基因的表达水平。结果(图8)显示,在春化35 d内,Bra039631、Bra020620、Bra033464、Bra001742、Bra017316基因的表达量均低于春化前,而其他6个基因的表达水平均高于春化前。以上结果表明,上述 BrbZIP 基因为白菜春化应答基因,在白菜春化过程中具有重要的作用。
3 讨 论
bZIP转录因子基因家族是植物中数量比较多的基因家族,不同植物家族成员数量不等,但在进化上比较保守 [3] 。本研究经过严格筛选鉴定,从白菜基因组中获得118个 bZIP 基因,数量是拟南芥 bZIP 成员的1.6倍,与白菜 PRX 家族基因数量比值类似 [23] ,表明 BrbZIP 家族基因成员在白菜基因组倍化过程中,同样发生了基因丢失事件。通过系统发育树分析将已鉴定的白菜与拟南芥 bZIP 基因成员分为5大簇11个小簇,与拟南芥根据结构分组的结果类似 [4] ,表明白菜与拟南芥的 bZIP 基因在进化上是保守的。
在物种进化过程中,基因家族通常经历串联复制或大规模的片段复制,以维持家族较大规模 [8, 26] 。白菜经历过2次全基因组复制,及1次全基因组三倍化事件 [27] 。然而,白菜118个 bZIP 基因只有2對串联重复,数量显著少于油菜(247个成员,15个基因复制对) [5] ,但白菜基因组内共线性基因对高达110组,表明基因复制事件在芸薹属作物进化中的复杂性,以及在 bZIP 基因家族扩展中的重要性 [5, 28] 。此外,通过组织表达模式分析也发现,一些重复的 BrbZIP 的转录水平也相似,这可能与它们高度相似的结构和顺式调控元件有关。
对于多基因家族来说,基因表达分析常常为功能预测提供有用的线索。本研究利用白菜组织转录组数据 [20] 分析了 BrbZIP 在各组织中的表达模式。结果表明,大部分 BrbZIP 在白菜根、茎、叶、花、荚中均有较高的表达丰度,且在根组织中高表达丰度基因数量最多,这可能与根部应答逆境相关 [1] ;在各组织高表达丰度( FPKM >500)的 BrbZIP 基因中,各组织仅共有13个 BrbZIP 基因,说明大部分 BrbZIP 基因具有组织表达特异性,且这些组成型表达 BrbZIP 基因在白菜生长发育过程中具有重要的作用 [8] 。
由于目前对 bZIP 基因应答春化反应的调控作用知之甚少,我们探讨了 bZIP 基因在白菜幼苗春化反应中的表达模式。比较春化前后样本的表达数据 [21] ,结果显示白菜 bZIP 家族基因中应答春化反应表达有上调也有下调,结果与大麦( Hordeum vulgare )中类似 [29] 。在白菜两个材料中应答春化反应 bZIP 基因有42个,共有差异表达基因18个,其中上调基因8个,下调基因10个,表明这些基因与春化反应密切相关。为了了解它们的功能,对白菜18个春化应答关键 bZIP 基因进行了基因互作网络分析及荧光定量PCR分析,发现这些基因表现出较复杂的互作关系,其中下调表达基因Bra039631与Bra020620,及上调表达基因Bra004550与Bra020471作为互作网络的中心节点,在白菜应答春化过程中发挥着重要作用。
参考文献:
[1] 魏瑞敏,郑井元,刘 峰,等. 辣椒 bZIP 家族基因的鉴定与表达分析[J]. 园艺学报, 2018, 45(8): 1535-1550.
[2] LI D, FU F, ZHANG H, et al. Genome-wide systematic characterization of the bZIP transcriptional factor family in tomato ( Solanum lycopersicum L.) [J]. BMC Genomics, 2015, 16(1): 771.
[3] WOLFGANG D L, SNOEK B, BEREND S, et al. The Arabidopsis bZIP transcription factor family-an update [J]. Current Opinion in Plant Biology, 2018, 45: 36-49.
[4] JAKOBY M, WEISSHAAR B, DRGE-LASER W, et al. bZIP transcription factors in Arabidopsis [J]. Trends in Plant Science, 2002, 7(3): 106-111.
[5] YAN Z, XU D, JIA L, et al. Genome-wide identification and structural analysis of bZIP transcription factor genes in Brassica napus [J]. Genes, 2017, 8(11): 288.
[6] WANG Z, ZHU J, YUAN W, et al. Genome-wide characterization of bZIP transcription factors and their expression patterns in response to drought and salinity stress in Jatropha curcas [J]. International Journal of Biological Macromolecules, 2021, 181: 1207-1223.
[7] LI F, LIU J, GUO X, et al. Genome-wide survey, characterization, and expression analysis of bZIP transcription factors in Chenopodium quinoa [J]. BMC Plant Biology, 2020, 20(1): 405.
[8] ZHAO K, CHEN S, YAO W, et al. Genome-wide analysis and expression profile of the bZIP gene family in poplar [J]. BMC Plant Biology, 2021, 21(122): 122.
[9] JIN M, GAN S, JIAO J, et al. Genome-wide analysis of the bZIP gene family and the role of AchnABF1 from postharvest kiwifruit ( Actinidia chinensis cv. Hongyang) in osmotic and freezing stress adaptations [J]. Plant Science, 2021, 308: 110927.
[10] CHRISTOPH W, LORENZO P, JEBASINGH S, et al. The Arabidopsis bZIP11 transcription factor links low-energy signalling to auxin-mediated control of primary root growth [J]. PLoS Genetics, 2017, 13(2): e1006607.
[11] OYAMA T, SHIMURA Y, OKADA K. The Arabidopsis HY5 gene encodes a bZIP protein that regulates stimulus-induced development of root and hypocotyl [J]. Genes Development, 1997, 11(22): 2983-2995.
[12] HOU X, HU W, SHEN L, et al. Global identification of DELLA target genes during Arabidopsis flower development [J]. Plant Physiology, 2008, 147(3): 1126-1142.
[13] 徐 偉. 小麦 bZIP 基因 TaGBF 参与植物开花调控机制研究[D]. 济南: 山东大学, 2015.
[14] NAN H, CAO D, ZHANG D, et al. GmFT2a and GmFT5a redundantly and differentially regulate flowering through interaction with and upregulation of the bZIP transcription factor GmFDL19 in soybean [J]. PLoS One, 2014, 9(5): e97669.
[15] TAKESHIMA R, NAN H, HARIGAI K, et al. Functional divergence between soybean FLOWERING LOCUS T orthologues, FT2a and FT5a, in post-flowering stem growth [J]. Journal of Experimental Botany, 2019, 70(15): 3941-3953.
[16] LI M, HUA W, YUAN Y, et al. Capsella rubella TGA4, a bZIP transcription factor, causes delayed flowering in Arabidopsis thaliana [J]. Archives of Biological Sciences, 2015, 68: 141.
[17] 周 波,王 宇,孙 梅,等. 津田芜菁bZIP蛋白HY5 cDNA的克隆及表达特性[J]. 分子植物育种, 2008, 6(1): 59-64.
[18] LIU X, LI Z, HOU Y, et al. Protein interactomic analysis of SAPKs and ABA-inducible bZIPs revealed key roles of SAPK10 in rice flowering [J]. International Journal of Molecular Sciences, 2019, 20(6): 1427.
[19] CERISE M, GIAUME F, GALLI M, et al. OsFD4 promotes the rice floral transition via florigen activation complex formation in the shoot apical meristem [J]. New Phytologist, 2020, 229(1): 429-443.
[20] TONG C, WANG X, YU J, et al. Comprehensive analysis of RNA-seq data reveals the complexity of the transcriptome in Brassica rapa [J]. BMC Genomics, 2013, 14: 689.
[21] DAI Y, ZHANG S, SUN X, et al. Comparative transcriptome analysis of gene expression and regulatory characteristics associated with different vernalization periods in Brassica rapa [J]. Genes, 2020, 11(4): 392.
[22] CHEN G, WANG J, WANG H, et al. Genome-wide analysis of proline-rich extension-like receptor protein kinase (PERK) in Brassica rapa and its association with the pollen development [J]. BMC Genomics, 2020, 21: 401.
[23] 陈国户,王 浩,李 广,等. 白菜 PRX 基因家族的鉴定与生物信息学分析[J]. 浙江大学学报, 2020, 46(6): 677-686.
[24] CHEN C, CHEN H, ZHANG Y, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data [J]. Molecular Plant, 2020, 13(8): 1194-1202.
[25] 孙 宇,刘志鑫,叶 子,等. 杧果 RAV 基因家族的全基因组分析[J]. 江苏农业学报, 2021, 37(4): 957-967.
[26] CANNON S, MITRA A, BAUMGARTEN A, et al. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana [J]. BMC Plant Biology, 2004, 4: 10.
[27] CHENG F, MANDKOV T, WU J, et al. Deciphering the diploid ancestral genome of the mesohexaploid Brassica rapa [J]. The Plant Cell, 2013, 25: 1541-1554.
[28] FLAGEL L, WENDEL J. Gene duplication and evolutionary novelty in plants [J]. New Phytologist, 2009, 183(3): 557-564.
[29] POURABED E, GOLMOHAMADI F, MONFARED P, et al. Basic leucine zipper family in barley: genome-wide characterization of members and expression analysis [J]. Molecular Biotechnology, 2015, 57(1): 12-26.
(责任编辑:张震林)
收稿日期:2021-09-10
基金项目:国家自然科學基金项目(31801853);国家大学生创新创业基金项目(202110364080);安徽省大学生创新创业基金项目(S202010364238、S202010364239);安徽省博士后基金项目(2019B320)
作者简介:马李广(2000-),男,广东河源人,学士,研究方向为蔬菜种质资源与遗传育种。(E-mail)1554727563@qq.com。张贺龙为共同第一作者。
通讯作者:陈国户,(E-mail)cgh@ahau.edu.cn
