家蚕Bmlark基因启动子活性及转录调控元件分析
2022-07-14牛康康彭玉玲冯启理
吴 桐,牛康康,彭玉玲,冯启理
(华南师范大学生命科学学院,广州市昆虫发育调控与应用研究重点实验室,广东省昆虫发育生物学与应用技术重点实验室,广州 510631)
Lark基因最初是从果蝇羽化节律缺失的突变体中鉴定出来的(Newbyetal., 1993),是果蝇昼夜节律起搏器输出路径的组成成分(Zhangetal., 2000; Wangetal., 2005)。Lark在时间和空间的信息传输,导致行为变化的过程中发挥着重要作用,如调节神经兴奋性,这可能与LARK靶向调控编码钾离子通道的mRNA有关(Huangetal., 2009),或者通过调节信号分子如激酶或者磷酸酶从而控制离子通道和兴奋性。LARK表达水平的节律性变化对于成虫羽化的昼夜节律调节非常重要,该蛋白质在介导蜕皮的时钟调节中起抑制作用,lark基因产物增加会导致迟羽化表型(Newby and Jackson, 1996; Gerardetal., 1998; Iwai and Takeda, 2007)。
Lark在母体遗传和胚胎发育过程中发挥着重要的作用。McNeil(1999)等人发现在果蝇受精卵中lark开始进行转录之前(0~2 h),在前胚层胚胎中即含有丰富的larkmRNA,暗示了其作为母体遗传信号发挥着作用。在卵巢管发育的卵室中的营养细胞中也可以检测到lark表达,缺少lark母体遗传成分的未受精卵子和受精卵都表现出“脆弱”表型,大多数胚胎在胚盘形成之前停止进一步发育,还会导致营养细胞细胞质倾倒和肌动蛋白细胞骨架组织的缺陷(Mcneiletal., 2009)。在本实验室前期工作中,在家蚕Bombyxmori中用CRISPR/Cas9基因编辑技术敲除Bmlark基因后,纯合子突变体Bmlark-/-的胚胎死亡,不能正常孵化出蚁蚕。
lark对神经元发育和功能也至关重要。有研究表明,LARK在整个胚胎中枢神经系统及成虫神经系统细胞中表达(Newbyetal., 1993)。在果蝇的中枢神经系统(CNS)和腹神经系统(VNS)中检测到LARK的存在,在含有甲壳类动物心脏活性肽CCAP(Crustacean cardioactive peptide)的神经元中,LARK定位于细胞质中(Huangetal., 2007),而在其他神经元中定位于细胞核。在其他组织中LARK也存在于细胞核和细胞质中,暗示了LARK具有复杂的功能,可能在RNA加工、转运、翻译等过程中起着调控作用(Kojimaetal., 2007; Huangetal., 2014)。
Bmlark作为具有重要生物学功能的基因,其转录调控机制尚不明确。本研究通过构建Bmlark基因启动子双萤光素酶报告系统,鉴定其调控活性区及可能与之结合的调控蛋白,为进一步研究Bmlark基因转录调控的分子机制提供线索。
1 材料与方法
1.1 材料
家蚕Bm12卵巢细胞株来自中山大学徐卫华教授实验室;胎牛血清(FBS)和Grace液体培养基购自美国Gibco公司。无血清培养基Opti-MEM购自美国Invitrogen公司。X-tremeGENETMHP DNA转染试剂购自美国SIGMA公司。一步法克隆试剂盒ClonExpress©ⅡOne StepCloning Kit购自美国Vazyme公司。大肠杆菌菌株DH5α购自深圳康体生命科技有限公司。dNTP (10 mM)、rTaq DNA聚合酶、限制性内切酶XhoⅠ、HindⅢ购自Ttakara公司。凝胶DNA小量回收试剂盒购自Magen公司。质粒提取试剂盒购自天根深化科技有限公司。双萤光素酶报告基因检测试剂盒购自上海翊圣生物科技有限公司。核蛋白提取试剂盒和LightShift©Chemiluminescent EMSA Kit购自美国Thermo公司。DNA引物合成及基因测序由北京擎科新业生物生物技术有限公司完成。蛋白质谱分析由广州唯誉智合科技有限公司完成。
1.2 方法
1.2.1家蚕Bmlark基因启动子序列的PCR扩增
在美国国家生物技术信息中心(National Center for Biotechnology Information,NCBI)数据库获得包含启动子区域的家蚕Bmlark基因序列(-1 232~ +211)(GenBank Accession No.: NC_051360.1)。通过Primer premier 5.0软件设计扩增该DNA序列的PCR引物,在上、下游引物的5′端分别引入与线性化载体两端一致的16 bp同源重组序列及XhoⅠ和HindⅢ酶切位点序列(表1)。以家蚕基因组DNA作为PCR模板进行目的片段的扩增。PCR反应体系(20 μL)为:2×taq Master 10 μL、DNA模板2.5 μL、上下游引物各0.5 μL、ddH2O 6.5 μL。PCR反应条件:95℃预变性3 min,95℃变性30 s,53℃退火30 s,72℃延伸90 s,共35个循环,72℃终延伸5 min。PCR产物经1%琼脂糖凝胶电泳检测PCR产物大小正确后,切胶并用DNA琼脂糖凝胶回收试剂盒回收纯化,扩增正确的PCR产物命名为P1(-1 232~ +221)。设计引物对P1进行系列截短PCR扩增,得到的目的PCR产物依次命名为P2(-529~ +211)、P3(-220~+211)、P4(-124~ +211)、P5(-92~ +211)、P6(-66~ +211)和P7(-8~ +211)。所有PCR产物均经过测序验证其正确性。

表1 扩增Bmlark基因启动子区各序列片段的PCR引物Table 1 PCR primers for amplifying each sequence fragment in the promoter region of Bmlark gene
1.2.2 家蚕Bmlark基因启动子萤光素酶报告质粒的构建
用限制性内切酶XhoⅠ和HindⅢ对pGL3-Basic质粒进行酶切,并切胶回收纯化。家蚕Bmlark基因启动子PCR产物与线性化的pGL3-Basic质粒进行连接反应,反应条件参照vazyme ClonExpress©ⅡOne StepCloning Kit说明书。重组产物转化大肠杆菌DH5α感受态细胞,然后涂布于氨苄青霉素抗性平板培养基中37℃培养过夜。挑单克隆菌落进行扩增培养,提取质粒DNA进行目的片段的PCR鉴定,将大小正确的PCR产物进行测序。测序正确的重组质粒分别命名为pLP1(-1 232~ +221)、pLP2(-529~ +211)、pLP3(-220~ +211)、pLP4(-124~ +211)、pLP5(-92~ +211)、pLP6(-66~ +211)和pLP7(-8~ +211)。
1.2.3细胞培养与转染
用含有10%胎牛血清的Grace昆虫培养基,于27℃~28℃恒温培养中贴壁培养家蚕Bm12细胞。在24孔细胞培养板里加入105mL的细胞,培养过夜,至细胞密度80%~90%时,进行转染。取0.1 μg各Bmlark基因启动子萤光素酶报告质粒、0.01 μg内参质粒SV40、转染试剂0.3 μL,加入25 μL Opti-MEM培养基中充分混匀,室温静置15~20 min,然后转染Bm12细胞。每组设置3个重复,同时设置pGL3-Basic空载体转染组作为阴性对照。
1.2.4启动子活性的萤光素酶测定
萤光素酶活性测定参照上海翊圣生物科技有限公司生产的双萤光素酶报告基因检测试剂盒说明书进行。上述细胞转染48 h后,每孔加200 μL裂解液,4℃摇床轻微震荡5 min,充分裂解细胞,10 000 rpm离心1 min,取20 μL上清和100 μL萤光素酶底物加入1.5 mL离心管中混匀,用Promega Glo Max发光检测仪测定萤光素酶活性M1后,加入100 μL反应终止液,继续测定海参萤光素酶发光值M2,M1/M2的置即为萤光素酶的相对活性(Relative Luciferase Activity,RLA),同一转染试验重复3次,取平均值。
1.2.5Bm12细胞核蛋白提取步骤
贴壁细胞生长密度至90%~100%后,将预冷的PBS直接加入到细胞培养瓶中清洗细胞两次,弃上清。每瓶加入360 μL胞浆提取缓冲液,均匀后冰上静置5 min。用吸头吹打几次后转移到预冷的1.5 mL离心管中,涡旋大力震荡15 s后,4℃最高速离心5 min;弃上清,用0.5 mL预冷PBS重悬沉淀,10 000 rpm离心5 min后弃上清,每样品加入150 μL核提取缓冲液,涡旋大力震荡15 s,冰上孵育1 min,重复4次;迅速将核提取物转入到预冷的离心管套管中,14 000 rpm,离心30 s,收集沉淀的核蛋白,保存于-80℃中。
1.2.6电泳迁移率转移法(DNA mobility shift assay,EMSA)检测
制备6%非变性聚丙烯酰胺凝胶。用0.5×TBE缓冲液,100 V电压、冰浴中预电泳1 h。DNA-蛋白结合反应 :通过北京擎科新业生物生物技术有限公司合成Bmlark启动子32 bp(-124~-92)的寡核苷酸序列作为探针并进行生物素标记,同时合成未生物素标记的的竞争探针,探针序列For:5′-AATAATGTCGCCAAATATTATGCCGA AAAATAA-3′,Rev:5′-TTATTTTTCGGCATAATAT TTGGCGACATTATT-3′。DNA-蛋白结合反应混合液包括探针2 μL,核蛋白8 μL,2 μL 10×结合缓冲液,1 μL 50%甘油,1 μL 1% NP-40,1μL poly(dI-dC)封闭核蛋白非特异性结合,以及促进DNA和核蛋白结合的MgCl2、KCl和EDTA。上述混合液室温28℃放置20 min,作为对照,同时进行单纯探针反应和特异性竞争反应(在结合反应体系的基础上分别50倍和100倍未标记的探针)。上样后,于0.5×TBE缓冲液,100 V电压进行电泳2 h。电泳后转膜,转膜完成后,将膜取出,置于玻璃皿中,溴酚蓝面向上,将膜放置于紫外灯(波长254 nm)下进行交联0.8 min。根据Thermo LightShift©Chemiluminescent EMSA Kit的方法进行化学发光检测生物素标记的DNA探针与蛋白结合的情况。
1.3 统计学处理
利用GraphPad Prism 7软件对实验数据进行分析和展示,萤光素酶的相对活性以“平均值±标准差”表示,对各启动子片段相对萤光素酶活性采用t检验,*P<0.05表示最低的显著意义,**P<0.01表示中等程度的显著意义,***P<0.001表示最高的显著意义,P>0.05表示差异不显著。
2 结果与分析
2.1 Bmlark基因启动子转录活性检测
以家蚕基因组DNA为模板,上游引物Bmlark-F(-1 232)和下游引物Bmlark-R (+211)进行PCR扩增,获得一约1.4 Kb的PCR产物P1(-1 232~ +211)(图1-A)。经测序验证,表明该PCR产物为Bmlark启动子目的片段。
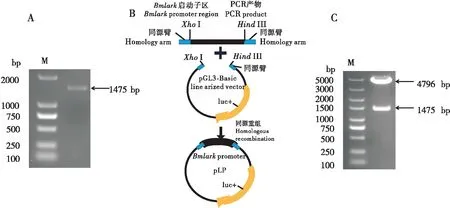
图1 家蚕Bmlark基因启动子萤光素酶报告质粒的构建Fig.1 Construction of the luciferase reporter plasmid of the Bmlark gene promoter in silkworm注:A,家蚕Bmlark基因启动子序列P1(-1 232~ +211)的PCR扩增产物的凝胶电泳分析;M,DL2000 DNA标准分子量;B,利用同源重组构建家蚕Bmlark基因启动子萤光素酶报告质粒示意图;C,家蚕Bmlark基因启动子萤光素酶报告质粒pLP1(-1 232~ +211)的Xho Ⅰ和Hind Ⅲ双酶切鉴定分析;M,DL5000 DNA标准分子量。Note: A, Gel electrophoresis analysis of the PCR amplified product of the promoter sequence P1 (-1 232~ +211) of the silkworm Bmlark gene; M, DL2000 DNA standard molecular weight; B, Schematic diagram of constructing the luciferase reporter plasmid of the silkworm Bmlark gene promoter by homologous recombination; C, Xho Ⅰ and Hind Ⅲ double enzyme digestion analysis of the luciferase reporter plasmid pLP1 (-1 232~ +211) of the Bmlark gene promoter; M, DL5000 DNA standard molecular weight.
上述PCR产物的两端设计有与pGL3-Basic萤光素酶报告载体多克隆位点同源的接头序列。利用XhoⅠ和HindⅢ两个酶切位点,酶切使pGL3-Basic载体线性化,随后将Bmlark基因启动子片段P1(-1 232~ +211),通过同源重组的方式克隆至pGL3-Basic质粒中(图1-B)。转化大肠杆菌细胞后提取质粒后双酶切验证,结果显示,该1 475bp目的片段已重组到pGL3-Basic中。测序结果显示,插入序列完全正确,表明含家蚕Bmlark基因启动子片段的萤光素酶报告质粒pLP1(-1 232~ +211)构建成功。
为验证克隆得到的家蚕Bmlark基因启动子是否具有转录活性,将报告质粒pLP1和pGL3-Basic空载体分别与SV40海肾萤光素酶报告质粒(内参)共转染家蚕Bm12细胞。萤光素酶活性检测结果显示,表达pLP1细胞中的萤光素酶活性比pGL3-Basic阴性对照增加约461倍(P=0.0002,图2),表明家蚕Bmlark基因的-1 232~ +211 bp区域具有启动子活性。
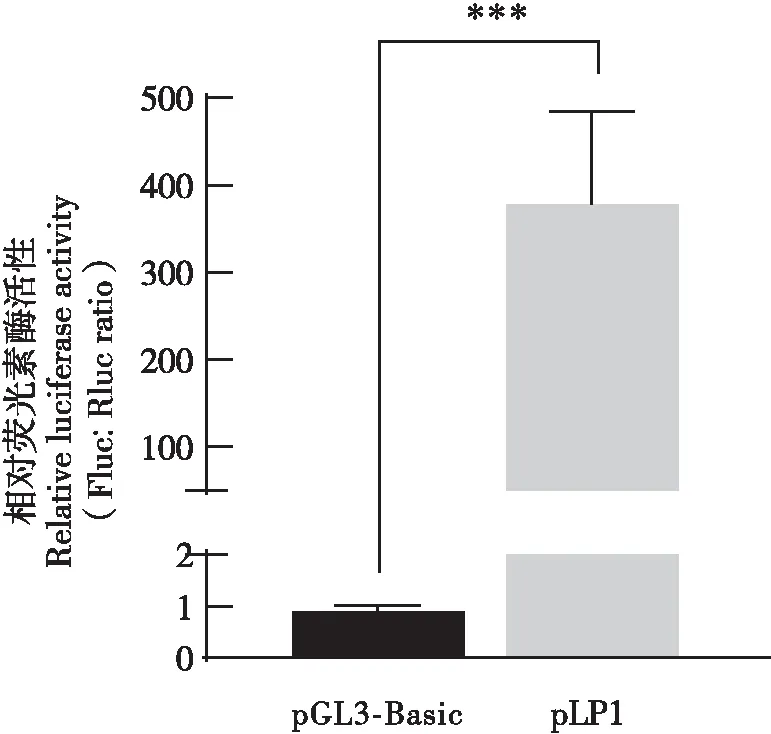
图2 家蚕Bmlark基因启动子萤光素酶报告质粒pLP1(-1 232~ +211)转录活性检测Fig.2 Detection of the transcriptional activity of the Bmlark gene promoter luciferase reporter plasmid pLP1 (-1 232~ +211)注:萤光素酶的相对活性以“平均值±标准差”表示,n=3,***P<0.001。Note: Relative activity of luciferase was expressed as "mean±standard deviation", n=3, ***P<0.001.
2.2 Bmlark基因启动子转录调控活性区的鉴定
为进一步定位Bmlark基因启动子的转录调控活性区,以pLP1(-1 232~ +211)为模板,采用移步截短法设计引物对pLP1(-1 232~ +211)进行系列截短扩增,得到6个大小与预期一致的截短片段:P2(-529~ +211)、P3(-220~ +211)、P4(-124~ +211)、P5(-92~ +211)、P6(-66~ +211)、P7(-8~ +211)(图3-A),测序结果显示序列正确。经同源重组分别把这些片段插入到pGL3-Basic载体。对阳性重组质粒进行双酶切鉴定(图3-B)和测序,结果显示插入序列完全正确,证明系列截短的Bmlark基因启动子萤光素酶报告质粒pLP2(-529~ +211)、pLP3(-220~ +211)、pLP4(-124~ +211)、pLP5(-92~ +211)、pLP6(-66~ +211)、pLP7(-8~ +211)构建成功。
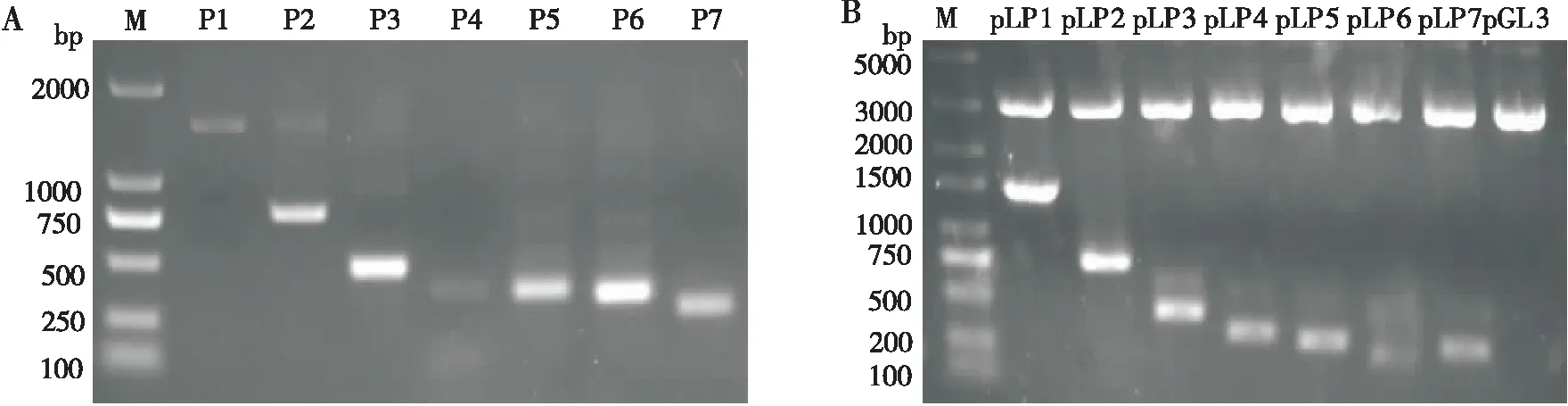
图3 系列截短的Bmlark基因启动子萤光素酶报告质粒的构建Fig.3 Construction of a series of truncated Bmlark gene promoter luciferase reporter plasmids注:A,系列截短的家蚕Bmlark基因启动子序列PCR扩增产物的凝胶电泳;M,DL2000 DNA标准分子量;B,系列截短的家蚕Bmlark基因启动子萤光素酶报告质粒XhoⅠ和Hind Ⅲ双酶切鉴定;M,DL5000 DNA标准分子量。Note: A, Gel electrophoresis of PCR amplified products of a series of truncated silkworm Bmlark gene promoter sequence; M, DL2000 DNA standard molecular weight; B, A series of truncated silkworm Bmlark gene promoter luciferase reporter plasmids XhoⅠ and Hind Ⅲ double restriction digestion identification; M, DL5000 DNA standard molecular weight.
将不同片段长度的重组质粒与内参SV40质粒共转染Bm12细胞,以pGL3-Basic作为阴性对照同步转染,48 h后进行双萤光素酶检测。质粒pLP1、pLP2、pLP3、pLP4、pLP5、pLP6的萤光素酶表达水平均较pGL3-Basic显著增加。其中,pLP4的萤光素酶表达最高,较pLP3升高了约13倍(P=0.0004),暗示在-220~ -124 bp启动子区域可能存在转录抑制因子结合位点或负向调控元件。pLP5的萤光素酶表达水平较pLP4下降低了约5倍(P=0.0007),pLP6的萤光素酶表达水平较pLP5又下降了约6倍(P=0.0007),暗示在-124~ -92 bp可能存在转录激活因子结合位点或正调控元件(图4)。

图4 系列截短的家蚕Bmlark基因启动子萤光素酶报告质粒的转录活性检测Fig.4 Transcriptional activity detection of a series of truncated silkworm Bmlark gene promoter luciferase reporter plasmids注:萤光素酶的相对活性以“平均值±标准差”表示,n=3,***P<0.001。Note: Relative activity of luciferase was expressed as “mean±standard deviation”, n=3, ***P<0.001.
2.3 启动子结合蛋白的EMSA分析
为了鉴定是否有核蛋白结合到该-124/-92 bp潜在的转录激活因子结合位点或正调控元件序列,从Bm12细胞中提取的核蛋白后,以生物素标记的该片段(32 bp)探针(For:5′-AATAATGTCGC CAAATATTATGCCGAAAAATAA-3′,Rev:5′-TTAT TTTTCGGCATAATATTTGGCGACATTATT-3′)进行EMSA分析。结果显示有4个核蛋白与探针结合(图5)。当在反应体系中加入竞争探针(非标记的同样探针)的浓度增加时,蛋白1、蛋白2、蛋白4的结合信号消失,蛋白3结合信号在逐渐减弱但不能被探针完全竞争。结果表明,这些蛋白与探针的结合是特异性的。

图5 电泳迁移率转移法(EMSA)分析Bmlark基因启动子的-124~ -92 bp序列探针与Bm12细胞核蛋白的结合Fig.5 Electrophoretic mobility transfer method (EMSA)analysis of Bmlark gene promoter -124~ -92 bp sequence probe binding to Bm12 nuclear protein注:用生物素标记Bmlark基因启动子的-124~ -92 bp序列探针,未标记探针作为冷探针。Note: -124~ -92 bp sequence probe of Bmlark gene promoter was labeled with biotin, and the unlabeled probe was used as a cold probe.
将蛋白1~4切胶,并分别进行质谱检测。质谱结果显示protein 1、protein 2、protein 3、protein 4分别有177、124、104、95个候选蛋白(见S1),将获得的蛋白通过Omic Share网站及Gene Ontology数据库分析获得GO富集分析,这些蛋白主要与生殖过程、新陈代谢、生物调节相关,具有核酸及蛋白结合和催化等活性(图6)。根据蛋白分子量,是否细胞核蛋白以及是否具有DNA结合结构域等因素,鉴定出4个可能与该-124~-92 bp序列结合的蛋白,分别为TAF7(NC_051358.1),Ets transcription factor(NC_051374.1),protein abrupt isoform X1(NW_021011484.1)和forkhead box protein N3(XM_004928387.4)(表2)。今后将对这4个候选蛋白或转录因子进行进一步的实验验证。

图6 GO富集分析Fig.6 GO enrichment analysis注:将质谱分析获得的共500个蛋白质向Gene Ontology数据库的各个term映射,选取蛋白质显著富集的GO term,通过GO显著性分析能确定质谱检测到的蛋白质行使的主要生物学功能。Note: Map a total of 500 proteins obtained by mass spectrometry to each term in the Gene Ontology database, selected GO terms that were significantly enriched in different proteins, and used GO significance analysis to determine the main biological functions of the proteins detected by mass spectrometry. Cellular process,细胞过程;Single-organism process,单个组织过程;Metabolic process,代谢过程;Biological regulation,生物调控;Regulation of biological process,生物过程调控;Cellular component organization or biogenesis,细胞组分组织或合成;Response to stimulus,刺激反应;Localization,定位;Multicellular organismal process,多细胞生物过程;Developmental process,发育过程;Positive regulation of biological process,生物过程正调控;Negative regulation of biological process,生物过程负调控;Reproduction,生殖;Reproductive process,生殖过程;Multi-organism process,多组织过程;Signaling,信号发生;Locomotion,运动;Behavior,行为;Immune system process,免疫系统过程;Growth,生长;Biological adhesion,生物附着;Rhythmic process,生物节律过程;Presynaptic process involved in synaptic transmission,突触传导中的突触前过程;Cell,细胞;Cell part,细胞部分;Organelle,细胞器;Organelle part,细胞器部分;Macromolecular complex,高分子复合物;Membrane,膜;Membrane-enclosed lumen,有膜腔室;Membrane part,膜部分;Extracellular region,胞外区;synapse,突触;Synapse part,突触部分;Cell junction,细胞连接;Extracellular region part,胞外区部分;Supramolecular fiber,超分子纤维;Extracellular matrix,细胞外基质;Extracellular matrix component,细胞外基质部分;Binding,结合;Catalytic activity,催化活性;Transporter activity,转运活性;Transcription factor activity,转录因子活性;Protein binding,蛋白结合;Molecular transducer activity,分子传感器活性;Nuclei acid binding transcription factor activity,核酸结合转录分子活性;Structural molecule activity,结构分子活性;Electron carrier activity,电子携带活性;Molecular function regulator,分子功能调控;Signal transducer activity,信号传导活性;Protein tag,蛋白标签;Translation regulator activity,翻译调控活性。
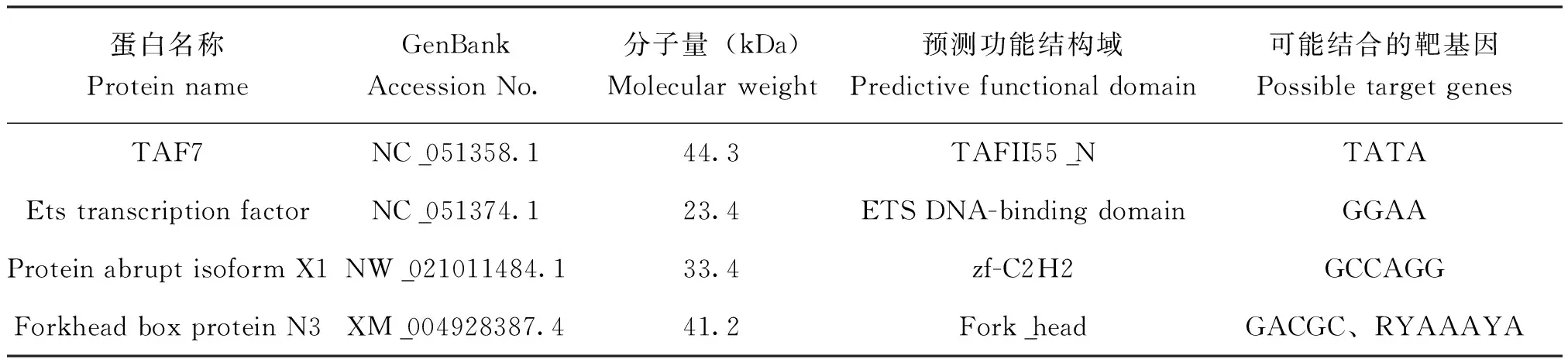
表2 候选4个蛋白的相关信息Table 2 Information about the 4 candidate proteins
3 结论与讨论
有研究发现,Bmlark是昼夜节律中输出通路的重要组成部分,对胚胎发育有着极其重要的作用,参于神经元发育和功能(Sundrametal., 2012),其表达水平受转录因子调控,但对于Bmlark的转录调控机制的研究尚无报道。本研究拟通过寻找Bmlark基因启动子的关键转录调控元件及结合蛋白,探索Bmlark基因的转录调控机制。
利用双萤光素酶报告基因系统鉴定基因启动子及调控元件,具有检测快、灵敏度高、操作简单、结果确切等优点。应用该方法,本研究对家蚕Bmlark基因的启动子区域进行了克隆并构建了家蚕Bmlark基因萤光素酶报告基因pLP1。与作为内参的SV40共同转染Bm12细胞后,确定家蚕Bmlark基因的-1 232~ +211 bp区域具有启动子活性,并进一步通过截短试验发现,在-124~ -92 bp启动子区域可能存在正向调控元件,可增强Bmlark基因的启动子活性。而在区域-220~ -124 bp可能存在转录抑制因子结合位点或负调控元件。这些结果表明,Bmlark基因的转录可能同时受到正和负的协同调控。经Jaspar在线网站预测,区域-124~ -92 bp包含转录因子Cad、Dbx、Ade-B等转录因子结合位点,区域-220~ -124 bp包含转录因子Su(Hw)、Br、H2.0等转录因子结合位点,但预测结合转录因子与EMSA获取的转录因子没有重合。
为了找到与-124~ -92 bp区域结合的调控蛋白,利用Bm12中提取的核蛋白进行EMSA分析,发现有4个蛋白结合条带protein1、protein2、protein3、protein4。经过甄别分析后,筛选出TAF7、Ets transcription factor、protein abrupt isoform X1、forkhead box protein N3等4个蛋白最有可能与该调控区结合,参与对Bmlark基因转录的调控。TAF7在RNA聚合酶II中可以通过结合TAF1来调节转录活性(Gegonneetal., 2008),是转录启动的流量控制器(Gegonneetal., 2013)。但也有研究指出,TAF7也可以作为一个独立的转录因子起作用(Devaiahetal., 2010)。TAF7对胚胎发育至关重要,是细胞增殖的关键调节因子,缺失TAF7会导致胚胎致死,TAF7在人类体内积极参与调节MHC I类基因的转录(Gegonneetal., 2012)。Ets transcription factor具有RNA聚合酶II顺式调节区序列特异的DNA结合活性,控制细胞周期、边界卵泡细胞迁移等过程,对神经系统、心脏、气管和眼睛的发育至关重要,参与表皮生长因子受体信号通路,成纤维细胞生长因子受体等信号通路(Poon and Kim, 2017; Sizemore and Pitarresi, 2017)。Protein abrupt isoform X1与蜕皮激素tai结合时会负调控蜕皮激素的信号传导。它调节器官和多种组织形态发生,介导神经元分化以及细胞迁移(Julieetal., 2011; Turkeletal., 2013; Turkeletal., 2015)。Forkhead box protein N3具有RNA聚合酶II顺式调节区序列特异性DNA结合活性(Rogersetal., 2019),参与器官形态发生、唾液腺形态发生、细胞周期调控以及碳水化合物和脂质代谢的发育(Kongetal., 2019)。虽然从蛋白的结构及EMSA分析结果来看,这些蛋白有可能与这个启动子片段结合,但直接证据仍需要通过获得这些蛋白后,再通过EMSA实验和其他方法加以验证。
综上所述,本研究成功构建了家蚕Bmlark基因启动子双萤光素酶报告系统,并发现-124~-92 bp区域可能存在转录激活因子结合位点或正调控元件,-220~ -124 bp可能存在转录抑制因子结合位点或负调控元件,并筛选出了可能结合在-124~ -92 bp正调控区域的蛋白。研究为进一步深入研究Bmlark基因转录调控提供了线索。
