中华蜜蜂越冬期抗寒生理生化指标研究
2022-07-14苏晓玲陈道印赵东绪华启云罗谷辉
苏晓玲,陈道印,赵东绪,华启云,罗谷辉
(金华市农业科学研究院,浙江金华 321017)
蜜蜂作为一种经济昆虫,不仅可以为人类提供营养丰富的蜂产品,而且对提高农作物产量、改善农产品品质和维护生态平衡具有十分重要的作用(Kleinetal., 2006)。低温是蜜蜂生存和繁衍的限制性因素之一,当外界温度较低和缺乏蜜源时,蜜蜂进入长达几个月的越冬期,而蜂群的安全越冬对蜂群的生存和翌年的养蜂生产至关重要;同时,蜜蜂作为我国自然生态系统和农业生态系统重要的授粉者,蜂群的成功越冬对于满足早春开花作物的授粉需求非常重要(Dokeetal., 2015)。
昆虫能够通过调节行为和改变体内生理生化反应来适应低温环境(Johannes and Heath, 2017)。蜜蜂以群体越冬,这是对低温环境的行为适应。当外界气温降到10℃以下时,蜂群内的蜜蜂形成一个球形蜂团(Phillipsetal., 1914),结团的蜜蜂通过振动飞行肌产生热量,维持蜂团外缘温度在6℃以上,从而使蜜蜂顺利越冬(Dokeetal., 2015)。另外,昆虫会发生一系列的生理生化反应,如降低过冷却点,减少体内含水量,排除体内冰核物质和积累抗寒物质等方式来抵抗冻害(陈豪等, 2010)。常志光等(2008)研究发现,卡尼鄂拉蜂A.m.carnicaPollmann, 1879体内水分、蛋白质和脂肪含量在越冬前期随气温下降逐渐增高,越冬后期随气温升高而降低,这几种物质含量与越冬蜂的抗寒性密切相关。秦明等(2017)研究表明,中华蜜蜂在应对寒冷环境刺激时主要通过降低体内游离水含量、提高糖原和小分子糖含量等方式降低体内过冷却点,提高自身耐寒性能。
长时间高温或低温胁迫可以诱导昆虫的氧化应激,诱发产生大量的活性氧,导致脂质过氧化损伤、蛋白羰基化损伤、DNA断裂等,最终导致机体功能障碍,昆虫的存活率下降(Rojasretal., 1996; Jiangetal., 2019)。抗氧化酶系统是昆虫等变温动物抵御温度胁迫的一个重要抗逆机制,正常情况下,细胞内自由基的产生与清除依靠超氧化物歧化酶(superoxide dismutase,SOD)、过氧化氢酶(catalase,CAT)和过氧化物酶(peroxidase,POD)等抗氧化酶的协调作用,使自由基维持在一个低水平(唐维媛等, 2016)。SOD能清除O2-形成H2O2,H2O2能与O2-形成毒性更强的HO-(Dubovskiyetal., 2008)。CAT和POD均能清除昆虫体内H2O2,CAT在H2O2浓度较高时发挥作用,而POD可以在低H2O2浓度的情况发挥高效的作用,二者共同作用将H2O2维持在较低水平(Kashiwagietal., 1997)。夏振宇等(2019)研究了中华蜜蜂短期低温胁迫下体内总抗氧化能力、SOD和POD酶活变化,证明了中华蜜蜂体内抗氧化酶在低温胁迫下协同发挥作用。
中华蜜蜂Apisceranacerana(简称“中蜂”)作为我国重要的本土资源,长期与我国当地的环境适应,具有较强的抗逆性。目前中蜂的饲养方式主要分为原始饲养与活框饲养。不同饲养方式的中蜂蜂群在自然越冬阶段的生理生化反应和抗氧化水平未见报道。本研究对越冬期活框饲养和原始饲养中蜂的蜂体水分、蛋白质、甘油等抗寒物质和SOD、POD、CAT等抗氧化指标进行测定,评价了越冬不同阶段的抗寒物质代谢水平和抗氧化水平,以期为中蜂抗寒生理机制提供依据;同时分析了不同饲养模式下中蜂对低温适应性的差异,以期为中蜂科学饲养管理提供理论依据。
1 材料与方法
1.1 试验蜂群
选择浙江省金华市罗店镇活框饲养和原始饲养的中华蜜蜂各5群,活框饲养蜂群采用郎氏标准箱饲养,越冬初期群势为3足框;原始饲养蜂群采用圆桶饲养,越冬初期群势为1.5足框。试验蜂群的群势为当地蜂场的常见群势。蜜蜂越冬前统一检查蜂群,补喂越冬饲料,越冬期间不进行操作。
1.2 试验方法
1.2.1样品采集
分别于越冬初期(2020年11月15日)、越冬中期(2020年12月15日)和越冬后期(2021年1月15日)进行工蜂取样,并以春繁期(2021年2月15日)蜜蜂作为对照样本。每个蜂群中随机取活蜂样本20头带回实验室,测定蜜蜂鲜重、游离水、蛋白质和糖原等抗寒生理指标。同时,取成年工蜂30头立即置于液氮冷冻,并于-80℃超低温冰箱内保存,用于测定蜜蜂SOD、POD和CAT等抗氧化指标。
1.2.2游离水含量测定
每群取工蜂10头,用电子天平称量10头蜜蜂鲜重(FW),然后将蜜蜂样本放入65℃电热恒温鼓风干燥箱中,48 h后测定10头蜜蜂干重(DW),计算蜂体的游离水含量(%)=[(FW-DW)/FW]×100(秦明等, 2017)。
1.2.3糖原含量测定
将蜜蜂解剖去掉蜜囊后,准确称取样品质量,加入0.75 mL提取液充分匀浆,按照糖原含量测试盒(苏州科铭生物技术有限公司)步骤测定糖原含量(秦明等, 2017)。每群测定3头,重复3次。
1.2.4蛋白质浓度测定
称取1头蜜蜂,加入2 mL PBS缓冲液,冰浴匀浆,8 000 g,4℃离心10 min,取上清,使用考马斯亮蓝法蛋白含量测试盒(苏州科铭生物技术有限公司)进行测定(Baoetal., 2020)。每群测定3头,重复3次。
1.2.5甘油含量测定
使用差重法准确称量蜜蜂样本重量,使用组织细胞甘油酶法测定试剂盒(北京普利莱基因技术有限公司)测定蜂体甘油含量。将标准品分别稀释至1 000、500、250、125、62.5和31.25 μmol/L后测定各标准管的吸光度,并绘制标准曲线y=0.0008x-0.0221,R2=0.9999,根据该方程将各样本的吸光度换算成甘油含量,再以每毫克蛋白浓度进行校正(徐凯等, 2018)。每次测定3头,重复3次。
1.2.6抗氧化指标测定
称取1头蜜蜂,加入1 mL提取液,冰浴匀浆,8 000 g,4℃离心10 min,取上清,分别使用超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)(苏州科铭生物技术有限公司)测试盒进行测定;称取1头蜜蜂,加入1 mL提取液,冰浴匀浆,10 000 g,4℃离心10 min,取上清,使用总抗氧化能力(FRAP法)(苏州科铭生物技术有限公司)测试盒进行测定,用蜂体蛋白质浓度进行校准(Wangetal., 2019)。每群测定3头,重复3次。
1.2.7气温数据获取
由金华市气象局提供样点附近气温数据。
1.3 数据统计与分析
试验数据采用SPSS 23.0软件进行单因素方差分析(one-way ANOVA),并采用LSD法进行多重比较分析,结果表示为平均值±标准差。P<0.05表示差异显著。
2 结果与分析
2.1 中蜂越冬期温度变化
根据金华市气象局气象数据统计,2020年11月-2021年2月取样点的月平均气温分别为15.3℃、7.9℃、6.4℃和12.1℃,月最低气温分别为7.3℃、-5.6℃、-6.4℃和1.3℃(图1)。可见,在蜜蜂越冬中期和后期外界环境温度较低。

图1 取样点温度变化Fig.1 Changes of temperature at the sampling point
2.2 越冬蜂体重
活框饲养中蜂和原始饲养中蜂体重(图2)在越冬期均呈“先升后降”趋势,但不同越冬阶段蜜蜂体重差异不显著(P>0.05);春繁期活框饲养中蜂体重显著增加至0.097 g(P<0.05),而原始饲养中蜂则显著降低至0.071 g(P<0.05),且两者达到差异显著水平(P<0.05)。
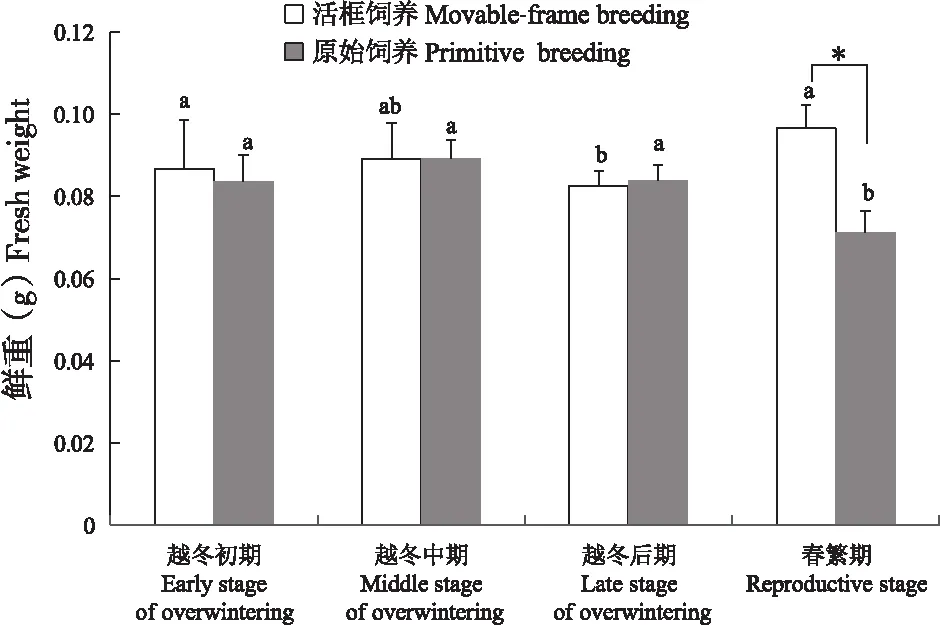
图2 中蜂体重Fig.2 Fresh weight of Apis cerana cerana注:柱上标有不同字母表示不同时间相同饲养方式中蜂差异显著(P<0.05);*表示不同饲养方式中蜂在同一时间差异显著。下图同。Note: Histograms with different lowercase letters indicated significantly different between different stages of the same breeding methods of Apis cerana cerana at 0.05 level. * indicated significantly different between different breeding methods of the same stages at 0.05 level. The same below.
2.3 越冬蜂游离水含量
活框饲养中蜂和原始饲养中蜂蜂体游离水含量(图3)在越冬期呈降低趋势,在越冬后期降至最低。活框饲养中蜂体内游离水含量在越冬初期达到最高为70.86%,在越冬后期降至68.86%,春繁期升高至70.90%(P<0.05),3个阶段游离水含量差异显著(P<0.05)。而原始饲养中蜂越冬初期游离水含量为76.64%,越冬后期降至75.08%,春繁期继续降低至72.19%,各个阶段差异不显著(P>0.05)。越冬期活框饲养中蜂蜂体游离水含量均低于原始饲养中蜂,且两者差异显著(P<0.05)。
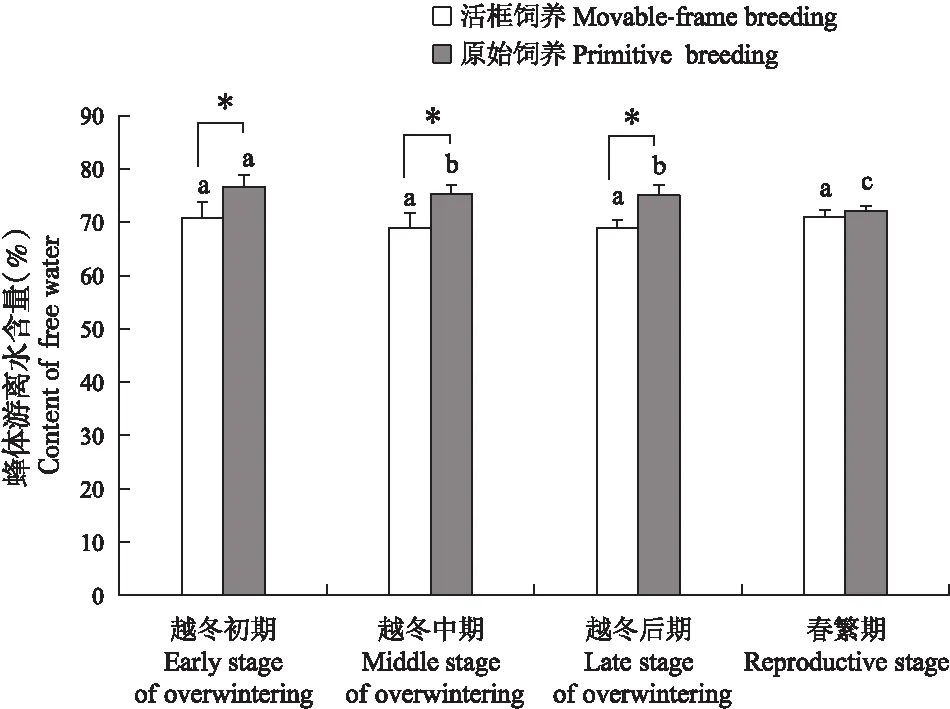
图3 中蜂游离水含量Fig.3 Content of free water of Apis cerana cerana
2.4 越冬蜂蛋白质浓度
活框饲养中蜂和原始饲养中蜂蛋白质浓度(图4)均呈“升-降-升”趋势,越冬中期蛋白质浓度最高,且与其他时期蛋白质浓度存在显著差异(P<0.05)。活框饲养中蜂蛋白质浓度由越冬初期的16.23 mg/g显著上升至越冬中期的19.72 mg/g(P<0.05),在越冬后期下降至14.77 mg/g(P<0.05),春繁期蛋白质浓度为15.53 mg/g,与越冬初期、越冬后期差异不显著(P>0.05)。原始饲养中蜂蛋白质浓度由越冬初期15.72 mg/g显著上升至越冬中期的18.24 mg/g(P<0.05),在越冬后期下降至14.76 mg/g(P<0.05),春繁期蛋白质浓度为16.80 mg/g,较越冬后期显著上升(P<0.05)。相同阶段的两种饲养方式中蜂蛋白质浓度差异不显著(P>0.05)。
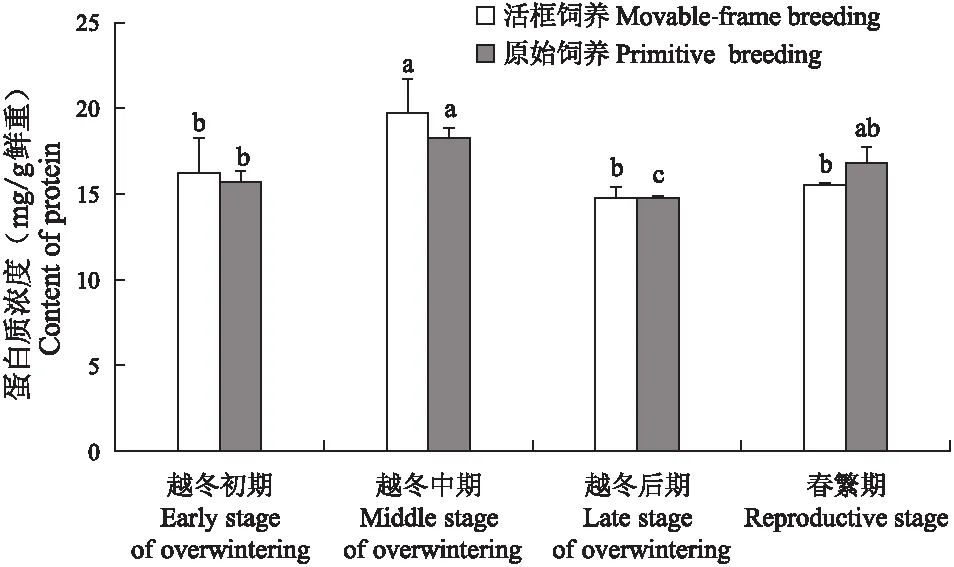
图4 中蜂蛋白质浓度Fig.4 Content of protein of Apis cerana cerana
2.5 越冬蜂糖原含量
两种饲养方式的中蜂糖原含量(图5)均呈“先降后升”趋势,越冬中期与越冬后期糖原含量最低。活框饲养中蜂的糖原含量由越冬初期的8.30 mg/g下降至越冬中期的2.08 mg/g(P<0.05),至越冬后期糖原含量下降至最低1.57 mg/g,春繁期上升至3.69 mg/g(P<0.05)。原始饲养中蜂糖原含量由越冬初期的10.07 mg/g下降至越冬中期的2.99 mg/g(P<0.05),由越冬后期的3.03 mg/g上升至春繁期的6.11 mg/g(P<0.05)。活框饲养中蜂的糖原含量在越冬阶段和春繁期均低于原始饲养中蜂,越冬期差异不显著(P>0.05),春繁期差异显著(P<0.05)。
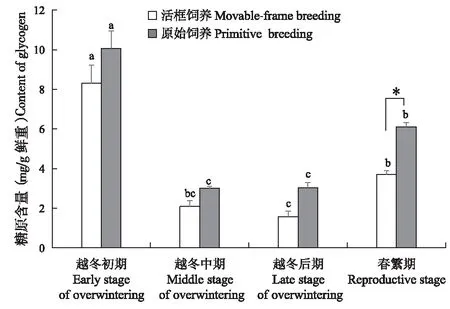
图5 中蜂糖原含量Fig.5 Contert of glycogen of Apis cerana cerana
2.6 越冬蜂甘油含量
两种饲养方式的中蜂甘油含量(图6)呈“先升后降”趋势,越冬后期甘油含量最高,在相同阶段不同饲养方式中蜂之间甘油含量差异均达到显著水平(P<0.05)。活框饲养中蜂甘油含量在越冬初期(362.50 μmol/mg prot)与越冬中期(350.46 μmol/mg prot)无显著差异(P>0.05),在越冬后期显著上升至483.11 μmol/mg prot(P<0.05),于春繁期显著下降至117.96 μmol/mg prot(P<0.05)。原始饲养中蜂甘油含量由越冬初期的236.62 μmol/mg prot、越冬中期的245.26 μmol/mg prot上升至越冬后期的319.92 μmol/mg prot,差异显著(P<0.05),在越冬后期又显著下降至263.04 μmol/mg prot(P<0.05)。活框饲养中蜂越冬期甘油水平均显著高于原始饲养中蜂(P<0.05),春繁期显著低于原始饲养中蜂(P<0.05)。
2.7 抗氧化指标
2.7.1SOD、CAT和POD酶活变化
越冬期和春繁期活框饲养中蜂SOD(图7)、CAT(图8)和POD(图9)变化趋势一致,呈“降-升-降”趋势,但不同酶活的变化幅度不一致。在越冬初期SOD、CAT和POD酶活较高,越冬中期出现显著下降(P<0.05),分别由194.53、49.32 和60.43 U/mg prot下降至117.95、43.08和28.19 U /mg prot,且差异显著(P<0.05);越冬后期酶活性分别升高至149.07(P<0.05)、48.89和75.26 U/mg prot(P<0.05),随后春繁期再降低至69.81(P<0.05)、43.15和72.31 U/mg prot。

图7 中蜂SOD酶活性Fig.7 Enzymatic activity of SOD of Apis cerana cerana

图8 中蜂CAT酶活性 Fig.8 Enzymatic activity of CAT of Apis cerana cerana

图9 中蜂POD酶活性Fig.9 Enzymatic activity of POD of Apis cerana cerana
原始饲养中蜂SOD呈“先升后降”趋势,由越冬初期的153.41上升至越冬中期的203.89(P<0.05),越冬后期显著下降至161.07 U/mg prot(P<0.05),直至在春繁期降至最低的77.15 U/mg prot(P<0.05)。原始饲养中蜂CAT与POD酶活呈“先降后升”趋势,分别由越冬初期的50.71和101.24 U/mg prot下降至越冬中期的46.24和22.10 U/mg prot(P<0.05),在春繁期上升至53.81(P<0.05)和72.93 U/mg prot(P<0.05)。
两种饲养方式中蜂的SOD活性在越冬期差异显著(P<0.05),春繁期差异不显著(P>0.05);CAT活性与SOD相反;POD活性在越冬初期和越冬后期差异显著(P<0.05)。
2.7.2总抗氧化能力(T-AOC)
两种饲养方式之间的总抗氧化能力(图10)在相同阶段差异显著(P<0.05)。活框饲养中蜂越冬初期T-AOC含量为132.26 nmol Trolox/mg prot,越冬中期上升至149.99 nmol Trolox/mg prot(P<0.05),在越冬后期上升至220.73 nmol Trolox/mg prot(P<0.05),在春繁期稳定至213.27 nmol Trolox/mg prot(P>0.05)。原始饲养中蜂T-AOC含量由越冬初期的54.60上升至越冬中期与越冬后期的79.97与78.48 nmol Trolox/mg prot,春繁期显著上升至234.34 nmol Trolox/mg prot(P<0.05)。活框饲养中蜂的T-AOC在越冬阶段高于原始饲养中蜂,差异显著(P<0.05);在春繁期低于原始饲养中蜂,差异显著(P<0.05)。
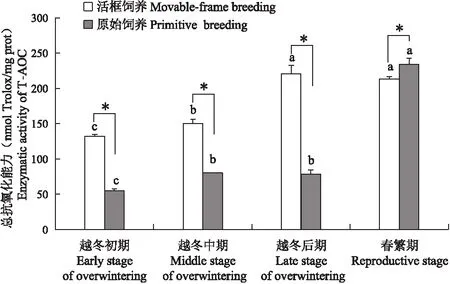
图10 中蜂总抗氧化能力Fig.10 Enzymatic activity of T-AOC of Apis cerana cerana
3 结论与讨论
3.1 中华蜜蜂越冬期间抗寒物质变化
越冬不同阶段蜜蜂的蛋白质含量呈“先升后降”趋势,表明越冬初期中蜂蛋白质等抗寒物质得到积累,到了越冬后期又被大量消耗。春繁期蜜蜂蛋白质含量升高,这可能是由于外界温度回升,蜜粉源植物开花泌蜜,蜜蜂采食蜂蜜和花粉后体内营养物质得以补充。两种饲养方式中蜂体重在春繁期表现出不同升降趋势,且差异显著,这可能是由于不同的饲养方式影响了蜂群的采集效率,但这需要进一步的实验论证。徐凯等(2018)比较了中蜂和意蜂Apismellifera低温处理前后17种血淋巴氨基酸含量,多数氨基酸在低温处理后(0℃)有一定的积累。本研究中蜂体蛋白质浓度在越冬初期到中期显著增加,越冬后期显著下降,这可能是蜜蜂在越冬初期进行越冬能量储备,在越冬中后期低温状态下,蛋白质分解成为氨基酸参与到蜜蜂的低温调控中,蛋白质是参与蜜蜂低温调控的重要物质。
越冬昆虫常在越冬前将体内游离水含量转化为结合水,降低虫体游离水含量,以提高过冷却能力而顺利越冬(Wolfeetal., 2002)。本研究中,蜜蜂游离水含量在越冬中期和后期出现下降,与任金龙等(2021)研究的意大利蝗卵在较冷的1-2月游离水含量和总含水量降低的结论一致,表明中蜂在低温环境体内游离水含量会显著降低,这也许是蜜蜂顺利越冬的重要抗寒机制(秦明等, 2017)。
甘油是昆虫体内重要的低温保护剂,甘油很容易穿过细胞膜,可以减少细胞膜上的渗透压力,起到稳定蛋白质和保护细胞膜的作用。在低温下机体细胞合成并积累一定的甘油,用以缓解冰冻与低温对细胞的机械损伤(Kostaletal., 2007; Johannes and Heath, 2017)。本研究中蜜蜂的甘油水平在越冬阶段持续增加,表明中蜂为了抵御越冬期低温环境使体内甘油水平得到了积累。徐凯等(2018)研究表明,10℃低温处理后蜂体甘油水平显著上调,本研究与其结果一致。越冬结束后,随着外界温度的升高,甘油水平降到较低水平,甘油含量变化与温度变化趋势相反,表明甘油是蜜蜂应对低温环境的一种调节物质,起到了低温保护作用。
糖原是昆虫越冬期的重要能量来源,在冬季温度降低时昆虫开始储存糖原,以保证顺利度过越冬期(景晓红和康乐, 2004)。昆虫抗寒物质的积累会提高昆虫抗寒能力,如甘油、海藻糖、葡萄糖和果糖以及氨基酸等物质(赵静等, 2008)。在本试验中发现,在越冬初期糖原含量显著高于其他阶段,表明中蜂在越冬前积累了大量的糖原,在越冬中期和后期糖原含量显著下降,可能蜜蜂在抵御外界寒冷时消耗了大量的糖原,致使体内糖原含量较低(欧阳芳等, 2014)。Rozsypal等(2013)研究表明,9月份刚开始滞育的苹果蠹蛾毛虫的糖原含量比7月份非滞育毛虫增加了一倍,这些糖原在越冬期被大量消耗,本研究结果与上述结果一致。在春繁期糖原含量显著增加,可能是有外界蜜源后,蜜蜂大量采食后体内糖原水平得以补充的结果。
综上所述,本研究中蜜蜂在自然越冬时通过降低体内游离水含量,增加甘油含量来增加抗寒能力,同时贮备大量糖原和蛋白质供越冬期能量消耗。
3.2 中华蜜蜂越冬期间抗氧化指标变化
昆虫体内广泛存在由SOD、CAT和POD等组成的抗氧化酶系统,在一定程度上影响到昆虫的抗逆能力(Siesetal., 1993)。SOD酶是抗活性氧自由基的第一道防线,能够催化生物体内的O2-生成H2O2,CAT酶与POD酶能够进一步催化H2O2进行分解成H2O和O2。本研究中,越冬初期SOD、CAT和POD酶活性处于较高水平,可能是低温诱导了蜜蜂体内SOD等抗氧化酶的升高,蜜蜂通过提高抗氧化酶的活性来降低体内氧化应激刺激,适应低温环境,这与夏振宇等(2019)研究的低温胁迫诱导了中华蜜蜂体内SOD活性的升高的结论一致。进入越冬中期,气温大幅降低,蜜蜂为维持生存,减少能量消耗,使酶活性降低。刘楠楠等(2020)研究欧洲黑蜂Apismelliferamellifera和新疆黑蜂的SOD和CAT酶活力均在越冬中期达到最低值,本研究中酶活力变化与上述结果一致。越冬中期酶活的抑制现象表明外界低温环境可能超过蜜蜂的耐受温度导致活性氧调节失衡,此时蜜蜂体内存在过量的氧自由基可能对蜜蜂造成了一定氧化损伤。越冬后期温度开始上升,蜜蜂体内3种抗氧化酶活性开始上升,清除过量O2-,恢复体内活性氧动态平衡,这与谌苗苗等(2020)研究结果类似。本研究发现,春繁期间SOD酶活性较越冬后期出现显著下降,而CAT和POD并没有显著差异。推测这可能是因为春繁时外界温度升高,蜜蜂体内O2-的产生与清除恢复动态平衡,因此SOD酶活处于较低水平,而蜜蜂体内H2O2仍未恢复到越冬前水平,导致CAT和POD酶活仍处于较高水平,这需要进一步测定蜂体的O2-和H2O2水平进行验证。
总抗氧化能力(T-AOC)是体现抗氧化体系的抗氧化能力总和,包含了直接作用在自由基或是间接消耗掉容易生成自由基的物质,防止自由基在机体内发生进一步反应,是反映生物体内抗氧化能力的重要指标(曹善茂等, 2018)。本研究两种饲养方式中蜂的T-AOC在整个越冬阶段不断增加,表明蜜蜂在越冬期通过不断提高抗氧化能力来应对环境低温刺激,这与欧洲黑蜂和新疆黑蜂的T-AOC在自然越冬时的变化规律一致(刘楠楠等, 2020)。
3.3 中蜂不同饲养方式抗寒物质和抗氧化水平比较
活框饲养中蜂在越冬初期和中期体重和蛋白质含量均高于原始饲养中蜂,且活框饲养中蜂体重在春繁期显著大于原始饲养中蜂(P<0.05)。蜜蜂体重与蜜蜂的个体大小呈正相关,蜜蜂体重和个体受到蜂王的生产性能、蜂群营养状况和巢房大小等因素的影响,而蜜蜂个体大在抗病力、生产力和抗逆性等方面更有优势(胡宗文等, 2020)。秦明等(2017)发现中蜂体内游离水含量显著低于意蜂,游离水越低,抗寒性越强。本研究中越冬期各个阶段活框饲养中蜂游离水含量均显著低于原始饲养中蜂(P<0.05),甘油含量显著高于原始饲养中蜂(P<0.05),表明活框饲养的中蜂个体表现出了对低温环境更强的抗逆性,有利于蜂群的顺利越冬。
越冬中后期外界温度达到全年最低水平,此时活框饲养中蜂的SOD和CAT酶活性均低于原始饲养中蜂,而POD活性却高于原始饲养中蜂,且总抗氧化能力也高于原始饲养中蜂,这可能表明活框饲养中蜂体内氧自由基水平更低,但蜜蜂体内还存在多种抗氧化损伤的机制或其他抗氧化物,它们协同发挥作用以避免冷刺激造成自身损伤。春繁期活框饲养中蜂的总抗氧化能力与3种抗氧化酶活性均出现了下降,而原始饲养的中蜂的总抗与POD酶活性仍出现了升高,这表明不同的饲养方式会影响蜜蜂体内的氧化自由基的生成,进一步影响到了蜜蜂体内的抗氧化能力。而本研究中活框饲养中蜂越冬群势较大,且在春繁期群势发展更快,因此,建议蜂农实际生产中可以通过饲养大群,合并弱群进行越冬,提高越冬成功率,加快春繁速度。
蜜蜂以群体越冬,蜜蜂的抗寒能力除了受到个体抗寒水平的影响,还受到蜜蜂群体的影响,群体对小环境的温湿度调节能力对蜂群抗寒力有重要作用,而群体大的蜂群具有更强的温湿度调节能力。原始饲养中蜂因不受干预,自由发展,越冬群势普遍较小;而活框饲养中蜂通过调整巢脾、培育越冬蜂等管理操作增加了越冬蜂群群势,因此可能具有更强的温度调节能力和抗寒能力。但两种饲养方式蜂群内部的温湿度缺少试验数据,蜜蜂的群体抗寒力还需进一步研究。
