贵州省东方蜜蜂Apis cerana线粒体DNA遗传多样性研究
2022-07-14赵金玉龙见坤DAVIDTembongCham陈祥盛常志敏
赵金玉,龙见坤,DAVID Tembong Cham,陈祥盛,常志敏,*
(1. 贵州大学动物科学学院,高原山地动物遗传育种与繁殖教育部重点实验室,贵阳 550025;2. 贵州大学动物科学学院,贵州省动物遗传育种与繁殖重点实验室,贵阳 550025;3. 贵州大学昆虫研究所,贵阳 550025;4. 贵州大学昆虫资源开发利用省级特色重点实验室,贵阳 550025)
东方蜜蜂Apiscerana又称中华蜜蜂Apisceranacerana,是我国分布最广、数量最多及饲养量最大的一个地理亚种,属中国独有蜜蜂品种,除新疆外,全国各地均有分布,尤其是南方的丘陵及山区(国家畜禽遗传资源委员会, 2011)。东方蜜蜂具有西方蜜蜂所不可比拟的优势,如抗寒御敌能力强,其7℃左右即可正常采集;另外,东方蜜蜂善于利用零星蜜源植物,采蜜期长,非常适合山区饲养。近年来,国家较重视东方蜜蜂的种质遗传资源保护,目前已划建多个保护区及保种场。贵州省是东方蜜蜂重要分布区,近5年来,其蜂群数量增至约51万群(韦小平等, 2020)。
目前,我国东方蜜蜂有北方型、华南型、华中型、云贵高原型、长白山型、海南型、滇南型、阿坝型及西藏型9个类型(国家畜禽遗传资源委员会, 2011)。东方蜜蜂遗传分化现状是其种质资源保护及开发利用的基础。近年来,mt DNA被广泛应用于东方蜜蜂遗传多样性研究。谭垦等(2004)对云南及北京地区东方蜜蜂mt DNA COⅡ基因进行分析,发现其存在碱基差异且属于不同类群。丁桂玲等(2008)对中国25个省100余个样点的东方蜜蜂mt DNA tRNAleu-COⅡ非编码区进行研究,获得18个单倍型及12个多态位点,并认为西北地区单倍型数量较多,蜜蜂多样性较丰富。刘振国等(2015)分析了我国20个地理种群东方蜜蜂mt DNA COⅡ基因遗传多样性,结果表明不同地理种群间存在显著遗传分化。谷瑛等(2016)对重庆139群东方蜜蜂mt DNA tRNAleu-COⅡ片段非编码区进行分析,发现不同生境种群间存在基因交流。曹联飞等(2017)基于tRNAleu-COⅡ序列对浙江省94个样点东方蜜蜂进行相关分析,发现20个单倍型。李彪等(2018)基于mt DNA tRNAleu-COⅡ非编码区对唐家河自然保护区37个蜂群进行分析,发现14个单倍型,指出单倍型分布与地理、气候等因素有较大关系。刘意秋等(2019)基于COⅠ片段分析了陕、甘、宁、青、晋5省东方蜜蜂遗传分化状况,定义了21个单倍型,并指出该地区种群总体遗传分化不显著。曹联飞等(2020)基于mt DNA tRNAleu-COⅡ序列对浙江金华地区东方蜜蜂遗传多样性分析,共发现14个单倍型,且发现该地区存在显著遗传分化。
贵州省地处云贵高原与湘桂丘陵之间,属温暖湿润季风高原山地气候区,西部为高原地貌,最典型的威宁地区,平均海拔2 200 m以上,其气候、植被与全省差异较大,年均气温13℃以下,降雨量较小,为低温少雨区,日照较丰富(徐祖荫, 1986);东部主要为山地和丘陵,植被茂盛、受外界干扰小、有保存较好的东方蜜蜂资源。因此,贵州蜜蜂资源是研究复杂地形下东方蜜蜂遗传分化的良好对象。近年来,仅于瀛龙等(2017)利用微卫星标记法对贵州省9个样点东方蜜蜂遗传多样性进行研究,认为贵州省东方蜜蜂未发生遗传分化。但由于涉及样点较少,贵州省东方蜜蜂是否存在遗传分化有待进一步研究。
1 材料与方法
1.1 样本采集
样本点选择标准:为减少实验误差,采样在蜜源植物丰富,种群群势较强的季节进行,并结合海拔、经纬度及距离等因素,优先选择野生蜂群及原始饲养蜂群,其次选择无人工育王蜂群。共采集79个理想样点,具体采样信息(见图1,表1),样本采集后暂存于装有无水乙醇的15 mL试管中,带回实验室后更换无水乙醇并保存于-20℃冰箱待用。
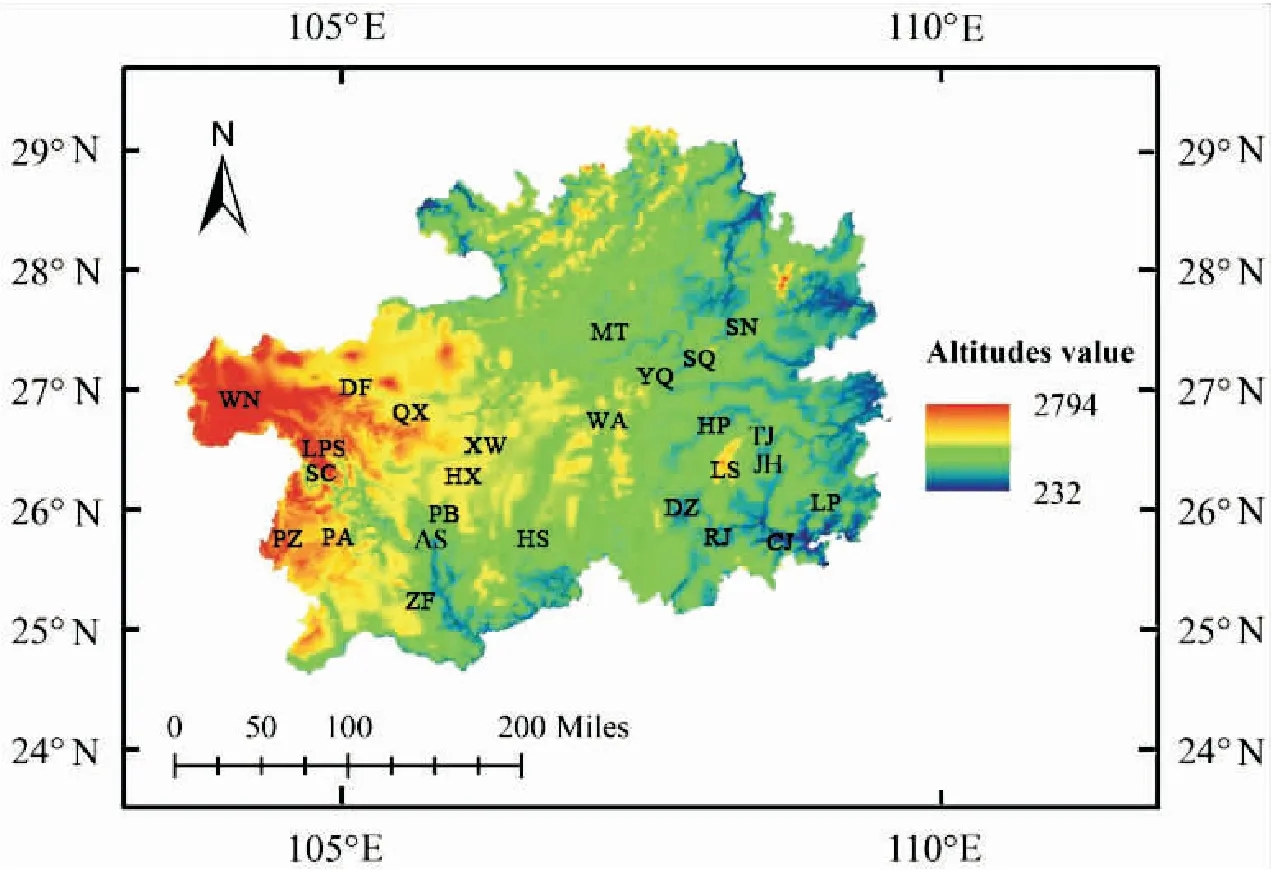
图1 东方蜜蜂样本采集信息图Fig.1 Information of Apis cerana sample collection

表1 东方蜜蜂样本采集信息表Table 1 Information of Apis cerana sample collection
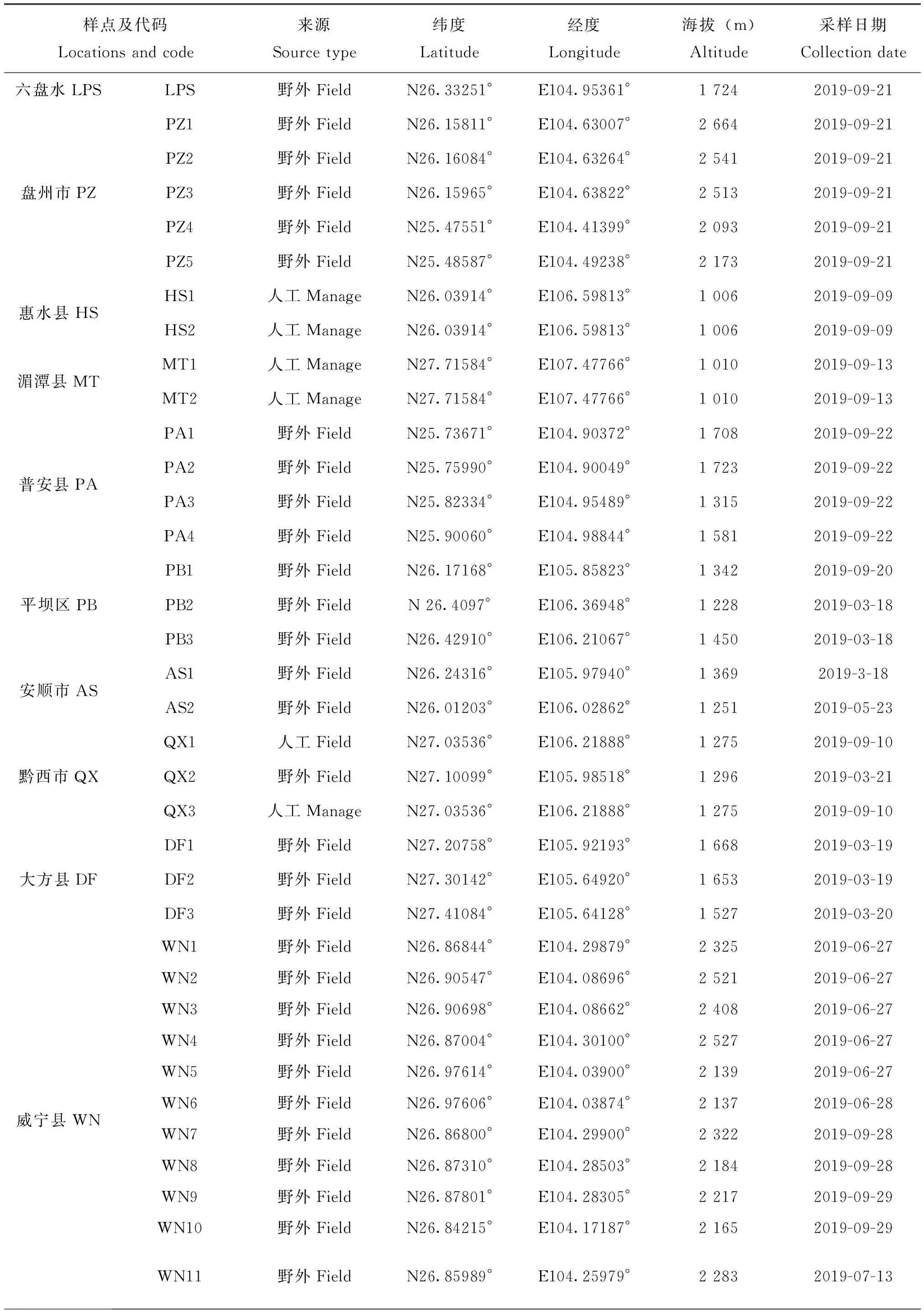
续表1 Continued table 1
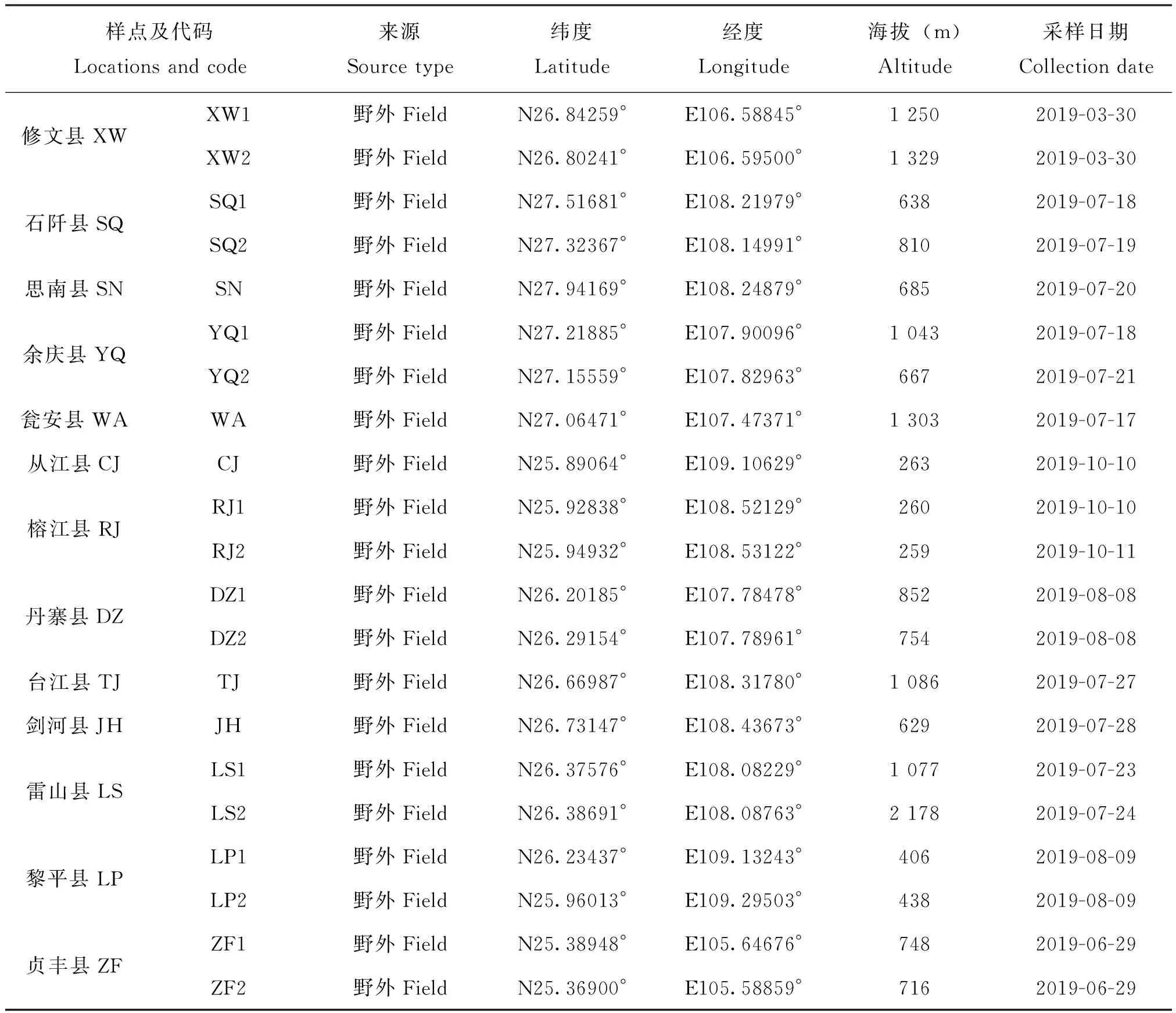
续表1 Continued table 1
1.2 试验方法
1.2.1总DNA提取
每群随机抽取1头无水乙醇保存的蜜蜂样本,取其胸部,尽量除去外表几丁质,置于1.5 mL离心管中加入TL Buffer后充分研磨,添加OB蛋白酶后55℃水浴4 h。具体方法及步骤参照动物基因组DNA快速提取试剂盒DP302(天根生化科技有限公司)。
1.2.2序列扩增与测序
依据东方蜜蜂mt DNA全基因组(Genbank Acession No: GQ162109)设计COI-COII片段的引物(Tanetal., 2016)。具体引物信息为R:5′-CTATACCTCGACGATACTCAG-3′,F:5′-TCAGGG TATTCATAGGATC-3′,并对mt DNA COI-COII序列进行扩增。PCR反应体系为20 μL,其中2×Taq MasterMix 10 μL,引物F及引物R各1 μL,DNA模板2 μL,ddH2O 6 μL。反应程序为:94℃ 5 min(预变性)、95℃ 30 s(变性)、54℃ 30 s(退火)、64℃ 30 s(延伸)、34个循环,72℃ 5 min(终延伸)。PCR产物送生工生物工程股份有限公司进行双向测序。
1.2.3数据分析
以地级市行政区划为单位,将贵州省79个样点分为9个地区样点,包括毕节地区(BJ)、六盘水地区(LPS)、黔西南地区(QXN)、黔南地区(QN)、黔东南地区(QDN)、铜仁地区(TR)、遵义地区(ZY)、贵阳地区(GY)、安顺地区(AS)。
使用Mega 6.05(Kumaretal., 2016)对9个地区样点进行比对、计算碱基组成、统计保守位点及变异位点、计算Kinura2-Parameter(K2P)平均遗传距离等;运用DNAsp 5.0(Rozasetal., 2017)检测并统计单倍型数目及分布,进而计算单倍型多样度及核苷酸多样度;然后使用DNAsp 5.0依据Tauna’s值(D)及Fu’s Fs值(Fs)进行中性检验,当两者均呈一个相对较大的负值时,表明种群可能经历过群体扩张;分别采用Mega 6.05 和Network 4.61(Bandeltetal., 1999)软件ML法构建单倍型系统发育树及单倍型网络中介图。
2 结果与分析
2.1 核苷酸碱基组成及变异位点
对贵州省9个地区样点东方蜜蜂mt DNA COI-COII序列测序校正后,获得片段长度约760 bp,碱基A、T、C、G平均含量分别为39.2%、43.1%、11.0%、6.7%,A+T平均含量为82.3%,C+G平均含量为17.7%,表现出明显的A+T偏移性。该片段中发现简约信息位点19个及单突变位点14个,共33个变异位点,其中变异位点A/G有10个,T/A有7个,T/C有8个,C/T 有6个,C/A有1个,T/G有1个。
2.2 单倍型分析
贵州省9个地区样点共检测到33种单倍型,分别命名为H1-H33。其中,共享单倍型11个(H1、H2、H3、H4、H5、H11、H13、H14、H16、H21、H23),占全部单倍型的33.33%;独有单倍型22个,占所有单倍型的66.67%(见表2)。在9个地区样点中,从单倍型出现的种类及数量来看,毕节地区最多,包括H1、H11、H14、H19、H20、H21、H22、H23、H24、H25、H26 11种单倍型,11种单倍型分布在17个样点;其次为黔东南地区,包括H1、H3、H5、H6、H7、H8、H28、H30、H31、H32 10种单倍型,分布在19个样点。

表2 33个单倍型分布情况Table 2 Distribution of 33 haplotypes
11个共享单倍型中,H1广泛分布在9个地区。从其出现频率来看,H1分布最多,其中六盘水地区分布4个样点(SC1、PZ1、PZ4、PZ5)、毕节地区分布5个样点(QX1、DF3、QX2、WN9、WN10)、黔东南地区分布8个样点(CJ、RJ1、RJ3、DZ1、JH、LP1、LP2、ZF1)等9个地区共分布33个样点;其次为H3,分布在六盘水地区(SC4、LPS)、黔南地区(HS1)、贵阳地区(HX5)共分布4个样点;H4、H11及H14均分布在2个样点,H4分布在贵阳地区(HX8)及六盘水地区(SC5);H11分布在六盘水地区(SC3)及毕节地区(WN11);H14分布在黔南地区(HS2)及毕节地区(WN2)。部分单倍型出现次数较多,但仅存在相邻较近样点,如H5仅分布在黔东南地区(HP3、DZ2、SS、LS2)4个较近样点,H13分布在六盘水地区(PZ2、PZ3)2个较近样点,H16、H21及H23均分布在相邻样点。
22个独有单倍型主要分布在毕节、黔东南、贵阳等地区,其中毕节地区有H19、H20、H22、H24、H25、H26 6种,分布在QX3、DF1、WN3、WN4、WN1、WN8 6个样点;黔东南有H6-H8、H28、H30-H32 7种,分布在HP2、HP4、HP5、TJ、CG、LS1 7个样点;六盘水地区有H9、H10、H12 3种,分布在SC2、SC6及SC7 3个样点;安顺地区有H17、H18 2种,分布在PB1及AS2 2个样点;贵阳、铜仁、遵义及黔西南地区均有1种,分别为H27、H29、H15、H33,分别分布在XW2、SN、MT1、ZF1 4个样点;黔南地区未发现独有单倍型。
2.3 核苷酸多样度及中性检验
总体平均单倍型多样度为0.802,其中黔南地区最高,为1.000,遵义地区最低,为0.500;总体平均核苷酸多样度为0.014,其中六盘水地区最高,为0.102,黔西南及安顺地区最低,均为0.002(表3)。
中性检验结果显示,黔东南、黔南、安顺、毕节地区的Tauna’s值(D)及Fu’s Fs值(Fs)均为负数,分别为-0.162和-1.330、-0.797和-1.514、-1.048和-0.186、-0.707和-0.227,表明上述地区东方蜜蜂可能经历过种群扩张,其余地区均有正有负(表3)。

表3 核苷酸多样度及中性检验结果Table 3 Nucleotide diversity and neutral test results
2.4 遗传距离分析
9个地区样点间平均遗传距离介于0.0012~0.0044之间,其中毕节与黔东南地区间最大,为0.0044;安顺与黔南、遵义地区间最小,均为0.0012(表4)。
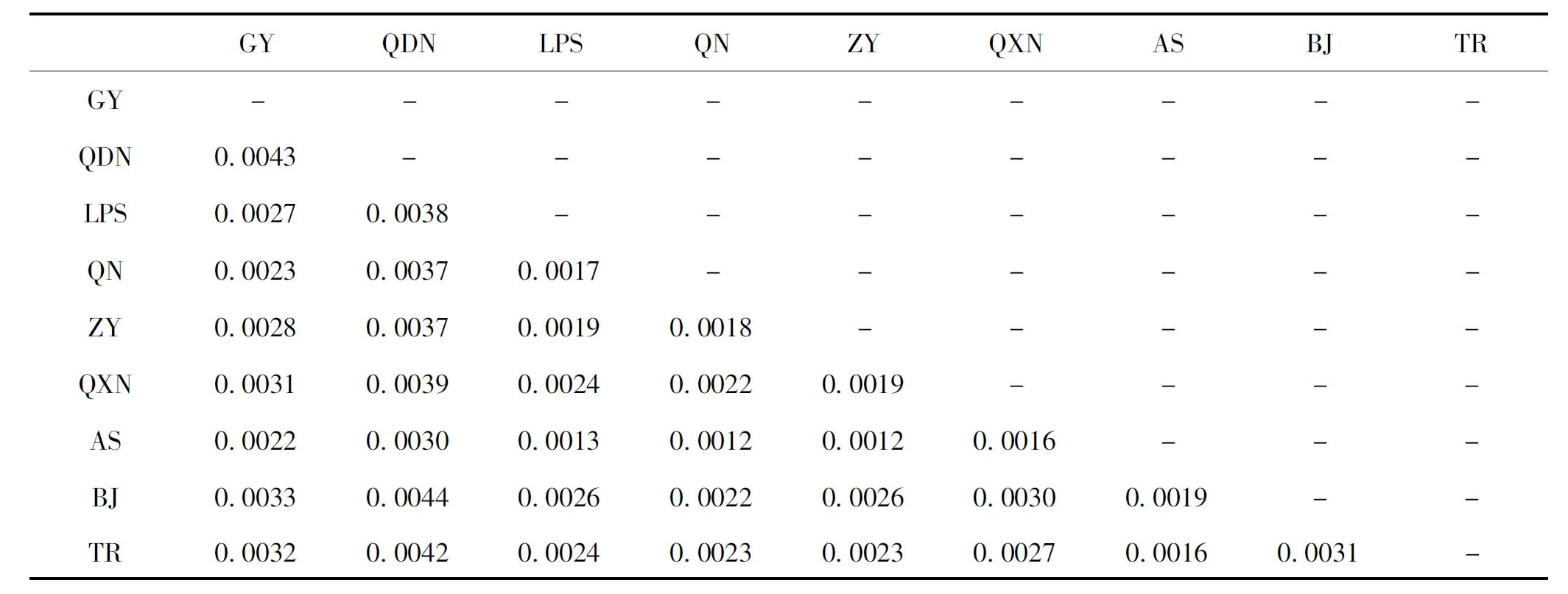
表4 9个地区样点间的K2P平均遗传距离Table 4 Average genetic distance of K2P among 9 sampling locations of Apis cerana
2.5 基于单倍型系统进化分析
基于ML法构建33个单倍型系统发育树,结果显示所有单倍型聚为2个大类群。一个类群包括H6、H15、H18及H23 4种单倍型,另外一大枝除3个小枝(H22及H25为一枝、H27及H28为一枝、H4及H29为一枝)外,均各自单独聚类在一起(图2)。

图2 基于ML法构建的单倍型系统发育树Fig.2 Construction of haplotype phylogenetic tree based on ML method
2.6 单倍型网络中介图
构建9个地区样点中33个单倍型网络中介图,其中H1面积最大,颜色类型最多,表明H1出现次数最多,分布最广,存在贵州省9个地区样点群体中,黔东南群体中分布最丰富,H1为最原始单倍型,其余单倍型均起源于此。同时可根据单倍型起源情况,将其分为3个大类,第一类包括H6、H15、H18、H23及H19,此大类和单倍型进化树基本一致;第二类包括H22、H25、H27-28及H14;其余单倍型为第三类(图3)。
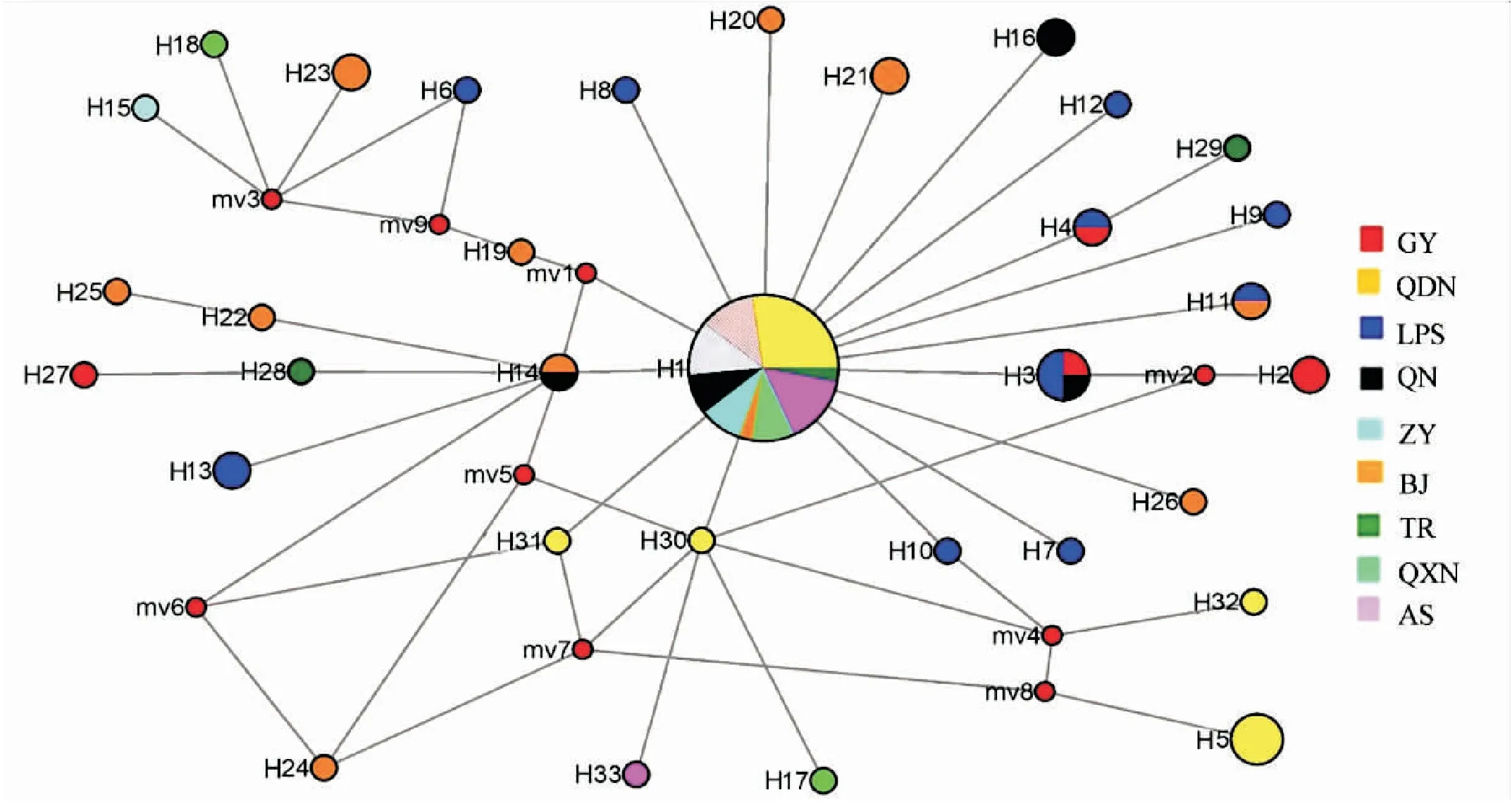
图3 单倍型网络中介图Fig.3 Haplotype network mediation diagram注:通过单倍型网络中介图可直观看出33个单倍型之间的亲缘关系,不同颜色代表不同样点种群,圆圈面积代表所含样本数量大小。Note: The relationship among 33 haplotypes could be obtainedby the haplotype network intermediary diagram. Different colors represented different geographical populations, and the circle area represented the number of samples.
3 结论与讨论
3.1 遗传多样性分析
本研究基于COⅠ-COⅡ片段对采自贵州省9个地区样点东方蜜蜂进行遗传多样性分析,结果表明,东方蜜蜂mt DNA序列在9个地区样点间存在一定变异。
碱基组成分析表明COⅠ-COⅡ序列中,A+T平均含量为82.3%,此片段为A+T富集区,该结果与前人研究结果相似,该序列碱基偏移性明显。Tan等(2011)研究表明亚洲地区东方蜜蜂该片段A+T平均含量为77.5%;冯毅楠等(2016)指出云南地区东方蜜蜂COⅠ-COⅡ片段A+T平均含量为82.4%;徐国威等(2014)指出四川省东方蜜蜂COⅠ-COⅡ片段A+T平均含量为82.8%。综上可知,贵州省与云南省、四川省东方蜜蜂的mt DNA COⅠ-COⅡ片段A+T平均含量相近。张卓等(2015)研究发现黑带食蚜蝇EpisyrphusbalteatusCOⅠ-COⅡ序列A+T平均含量为70.3%,其含量明显低于东方蜜蜂。
单倍型分析表明,9个地区样点中COⅠ-COⅡ基因共存在33个单倍型,11个为共享单倍型,22个为独有单倍型,平均总体单倍型多样度为0.802,其中遵义地区单倍型多样度为0.500,在所有样点中相对偏低,王俊杰(2018)研究发现遵义地区单倍型多样度在所有样本中也相对偏低,为0.400,可见该地区的东方蜜蜂种群与外界交流相对较少,也有学者认为这可能是东方蜜蜂与该地区特殊地理气候相适应而产生的一种结果(田嵩浩, 2014)。黔西南地区单倍型多样度也相对较低,其余7个样点单倍型遗传多样度均大于0.800。
3.2 遗传距离分析
9个地区样点间平均遗传距离介于0.0012~0.0044之间,其中黔东南地区与其它8个地区平均遗传距离相对较大,均大于0.0030;安顺地区与其它8个地区平均遗传距离相对较小,均小于0.0030。徐国威等(2014)研究表明四川省东方蜜蜂平均遗传距离介于0.0018~0.0049之间;冯毅楠等(2016)研究表明云南地区东方蜜蜂平均遗传距离介于0.0004~0.0012之间。综上,贵州省东方蜜蜂平均遗传距离大于云南省,略小于四川省。在实际距离上,毕节地区与黔东南地区相离最远,而遗传距离也最大;黔西南、贵阳与黔东南相离较远,其遗传距离也相对较大。黔东南地区与其它地区整体遗传较大,这可能与人为措施有关,如蜂群引进、人工育王、放蜂普遍等因素(张明华等, 2019; 徐祖荫, 2019)。安顺地区位于贵州省中部,其实际距离与其它地区相对较近,与其它地区遗传距离也最小。综上,贵州省东方蜜蜂各样点种群间遗传距离与实际距离可能存在一定相关性。
3.3 系统发育关系分析
单倍型系统发育树显示,33个单倍型聚为2个大类,第一大类包含H6、H15、H18及H23 4种单倍型,分别对应黔东南州黄平县纸坊乡、遵义市湄潭县云贵山、安顺市唐家湾、毕节市威宁县观风海样点;其余种群均包含在第二类群中。综上,东方蜜蜂种群未能按照相应地理区域聚类,这可能与盲目引种、放蜂等有关,张明华等(2019)调查发现每年初春两广及福建地区华南型中蜂被引入贵州地区,徐祖荫(2019)调查发现贵州地区有引进越南东方蜜蜂的现象,徐祖荫(2019)研究表明盲目引种,会导致当地蜂种混乱及退化,生产性能降低等现象。
33个单倍型网络图显示,单倍型H1扩散最广,出现频率最高,分布于以地级市为单位划分的所有样点,其次为H14,出现在毕节、黔南、铜仁及贵阳4个样点。根据逆祖理论(Gyllensten, 1991),可将H1认定为起源单倍型或祖先单倍型。
