草地贪夜蛾幼虫自残捕食量分析
2022-07-14吕宝乾唐继洪刘彦龙闫三强
唐 雪,吕宝乾,卢 辉,唐继洪,刘彦龙,闫三强
(1. 贵州大学昆虫研究所,贵州省山地农业病虫害重点实验室,贵阳 550025;2. 中国热带农业科学院环境与植物保护研究所/农业农村部热带作物有害生物综合治理重点实验室海南省南繁生物安全与分子育种重点实验室/海南省热带农业有害生物监测与控制重点实验室,海口 571101)
草地贪夜蛾Spodopterafrugiperda属鳞翅目Lepidoptera夜蛾科Noctuidae灰翅夜蛾属Spodoptera,是一种原产于美洲热带和亚热带地区的重大迁飞性农业害虫(Hardkeetal., 2015)。其拥有强大的迁飞扩散和繁殖能力,自2016年1月开始入侵非洲后,两年之内便快速扩散至非洲44个国家(王磊等, 2019),年潜在玉米损失量最高可达2 103万吨(Rogeretal., 2017)。2019年1月在我国云南省江城县首次被发现,随后迅速席卷国内多个省市,截止2019年10月已在我国26个省(自治区、直辖市)1 538个县(区)入侵为害,对我国的玉米、水稻、小麦等粮食作物的生产造成了严重威胁(张智等, 2021)。作为一种多食性害虫,草地贪夜蛾可以取食76科353种植物,包括玉米、水稻、甘蔗、高粱、小麦、棉花、甜菜、卷心菜、甜椒、香蕉、番茄等多种作物(Montezanoetal., 2018年)。玉米作为最受其喜爱的寄主植物,幼虫通常躲藏在心叶中取食(Changetal., 2000年),为其提供了一些免受捕食的保护,而心叶中幼虫密度过高可能导致食物竞争,因此自相残杀率会增加。
自相残杀(Cannibalism)或种内自残(Intraspecific cannibalism),是指杀死和取食部分或全部同种其他个体的行为(Fox, 2003),对种群动态有着重要的影响(Pierce, 1995)。自相残杀在鳞翅目昆虫已有广泛报道,例如甜菜夜蛾Spodopteraexigua、棉铃虫Helicoverpaarmigera等(Horner and Dively, 2003; 王娟等, 2008; Tangetal., 2016)。有研究表明昆虫的自相残杀行为可能会提高存活率、发育速度及繁殖力进而使其在种群竞争中获得直接优势,更有利于自身的生存繁殖,高龄幼虫通过捕食低龄幼虫,其发育速度相对提高,从而获得直接优势和间接优势(Yeetal., 2014)。虽然种群间的自相残杀可以给昆虫种群带来许多好处,但它可能会增加病原体和寄生虫在种群之间传播的风险,特别是受感染的个体更容易被同类捕食(Siva Jothyetal., 2001; Williams and Hernandez, 2010)。种内自残作为影响昆虫种群消长和群落结构的重要因素,已被作为生态学和进化生物学重要组成部分而受到广泛研究(Yasudaetal., 2001)。而草地贪夜蛾幼虫不仅在高密度和食物资源缺少的情况下会发生自相残杀现象,在食物不受限制的情况下也会发生自残现象(Andowetal., 1999; Chapmanetal., 1999)。此外,当不同龄期的幼虫被关在一起时,自相残杀的比率甚至会更高(Chapmanetal., 1999)。种内的自相残杀行为通常是一个更大、更强壮的个体杀死并吃掉一个更小、更弱的同类(Rudolf and Antonovics, 2007)。而在已有的相关研究中,多集中在不同密度下同一龄期间幼虫的自相残杀(Chapmanetal., 1999)。例如王道通等(2020)通过设置4个密度(5、10、20和30头/瓶)下相同龄期草地贪夜蛾的自相残杀率比较,得出不同密度下的自相残杀行为主要集中在5龄幼虫中,而没有针对高龄幼虫对低龄幼虫的捕食及捕食量进行深入研究,不能全面地反映草地贪夜蛾幼虫残杀的行为习性。因此,本研究设计了不同龄期的幼虫捕食2龄幼虫和3龄幼虫,通过拟合捕食功能反应Holling Ⅱ模型对草地贪夜蛾高龄幼虫捕食低龄幼虫的能力进行初步研究,以分析不同龄期的草地贪夜蛾对低龄幼虫捕食量的差异,探讨龄期和密度对草地贪夜蛾幼虫自残行为的影响,为进一步阐明草地贪夜蛾自残行为规律背后的生态意义提供参考。
1 材料与方法
1.1 供试虫源
草地贪夜蛾幼虫于2020年5月采自海南省儋州市宝岛新村六坡队玉米田(30°29′N, 114°18′E),在实验室以新鲜玉米叶饲养3代后供试。实验室温度25±1℃、相对湿度70%±5%、光周期12 L ∶12 D。
1.2 主要仪器设备
智能人工气候箱,圆形塑料盒(D=10 cm,H=5 cm),小毛笔,标签纸,镊子,电子天平等。
1.3 试验方法
1.3.1不同龄期幼虫对2龄和3龄幼虫的捕食作用
分别选取草地贪夜蛾3、4、5、6龄幼虫各1头置于不同塑料盒(D=10 cm,H=5 cm)中,饥饿处理24 h,对2龄幼虫的捕食作用,设置2龄幼虫密度为20、40、60、80、100、120头/盒,共6个梯度水平,并放入玉米叶片,叶面积约为24 cm2。对于3龄幼虫的捕食作用,选取4、5、6龄幼虫各1头置于塑料盒中后,设置3龄幼虫密度为20、40、60、80、100头/盒共,5个梯度水平,并放入玉米叶片,叶面积同上。每个处理重复5次。试验虫均放在温度25±1℃、相对湿度70%±5%、光周期12 L ∶12 D的恒温培养箱内培养。24 h后记录2龄幼虫和3龄幼虫的死亡数量,盒中消失的虫和仅剩一半的虫体均视为被捕食。由于低龄幼虫极少出现自相残杀现象,所以本实验不考虑低龄幼虫之间的自相残杀。
1.3.2不同龄期幼虫间的理论最大捕食量
对不同龄期的幼虫进行称重比较,记草地贪夜蛾2龄幼虫体重为1,以此计算不同龄期的幼虫相对于2龄幼虫的比值。利用Holling Ⅱ型圆盘方程进行模拟后得出3、4、5、6龄幼虫对2龄幼虫和4、5、6龄幼虫对3龄幼虫的最大理论捕食量,利用捕食量和体重的关系计算出每个龄期幼虫的最大理论捕食质量,根据各龄期的体重则可以换算出相应的捕食量。
1.4 分析方法
采用Holling Ⅱ(Holling, 1959)型圆盘方程进行拟合:
(1)
式中:Na—被捕食的猎物数量;A—捕食者的瞬时攻击率;N—猎物密度;Tr—总的试验时间(本实验以天为单位,故Tr=1);Th—处理1头猎物所需要的时间。
其中A和Th为待估参数。由于(1)式为非线性方程,将(1)式转化为以下线性方程:
(2)
将实验数据代入(2)式,用最小二乘法即可求得A和Th。将A和Th代入(1)式,即为拟合的草地贪夜蛾捕食2龄幼虫或3龄幼虫的功能反应模型。
要计算草地贪夜蛾理论最大捕食量时,将(1)变形为(T=1):
(3)
当N→∞时,Na有最大值即为1/Th,即为草地贪夜蛾的每日理论最大捕食量。用Excel 2010 处理数据,Graphpad Prism 8.02分析并作图。
2 结果与分析
2.1 草地贪夜蛾不同龄期幼虫对2龄幼虫的捕食功能反应
草地贪夜蛾3、4、5、6龄幼虫对2龄幼虫的日捕食量随2龄幼虫密度的升高而逐渐增加,当猎物密度增加一定水平时,草地贪夜蛾幼虫的日捕食量趋于平稳,捕食功能反应符合Holling Ⅱ型圆盘方程(图1)。草地贪夜蛾各高龄幼虫对2龄幼虫的捕食功能反应方程与Holling Ⅱ型圆盘方程模型拟合度较高,R2均在0.9以上(表1)。草地贪夜蛾3、4、5、6龄幼虫对2龄幼虫的瞬时攻击率依次为0.580、1.016、1.111、1.081;对2龄幼虫的处理时间依次为0.013、0.012、0.008、0.010 d。草地贪夜蛾3、4、5、6龄幼虫对2龄幼虫的日捕食量随着龄期的增加而增加,其中5龄幼虫对2龄幼虫的日最大捕食量最高,为117.92头,3龄幼虫的捕食量最小,为76.47头。而6龄幼虫的捕食量为96.05头,低于5龄幼虫,这可能与草地贪夜蛾6龄幼虫即将进入化蛹阶段有关,同时能说明草地贪夜蛾5龄幼虫的自残现象最为明显。
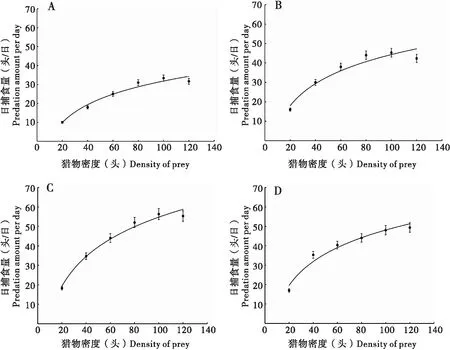
图1 草地贪夜蛾不同龄期幼虫对2龄幼虫的捕食功能反应Fig.1 Functional responses of Spodoptera frugiperda at different stages to the second instar larvae注:A、B、C、D分别代表草地贪夜蛾3龄、4龄、5龄、6龄幼虫对2龄幼虫的捕食功能反应,图中数值为平均值±标准误。Note: A, B, C and D showed the functional responses of the 3rd, 4th, 5th and 6th instar larvae of S.frugiperda a to the 2nd instar larvae respectively, data were means±SE.

表1 草地贪夜蛾不同龄期幼虫对2龄幼虫的捕食功能反应Table 1 Functional responses of Spodoptera frugiperda at different stages to the second instar larvae
2.2 草地贪夜蛾不同龄期幼虫对3龄幼虫的捕食功能反应
草地贪夜蛾4、5、6龄幼虫对3龄幼虫的捕食量随着3龄幼虫密度的增加而增加,当3龄幼虫的密度增加到一定限度时,草地贪夜蛾各高龄幼虫捕食量增加的速度逐渐变缓(图2);同一猎物密度,草地贪夜蛾5龄幼虫对3龄幼虫的捕食量均高于4龄和6龄幼虫对3龄幼虫的捕食量。由Hollling Ⅱ型公式拟合草地贪夜蛾各龄期的捕食量计算出草地贪夜蛾高龄幼虫对3龄幼虫捕食的功能反应方程及其参数如表2所示:4龄幼虫对3龄幼虫的捕食功能反应方程为Na=0.740N/(1+0.026N),对猎物的瞬时攻击率a为0.740,处理单头猎物所需要花费的时间Th为0.026 d,最大捕食量为38.83头/d;5龄幼虫对3龄幼虫的捕食功能反应方程为Na=1.345N/(1+0.017N),对猎物的瞬时攻击率a为1.345,处理单头猎物所需要花费的时间Th为0.017 d,捕食上限59.49头/d;6龄幼虫对3龄幼虫的捕食功能反应方程为Na=1.111N/(1+0.017N),对猎物的瞬时攻击率a为1.111,处理单头猎物所需要花费的时间Th与5龄幼虫相同,为0.017 d,捕食上限为58.87头/d。
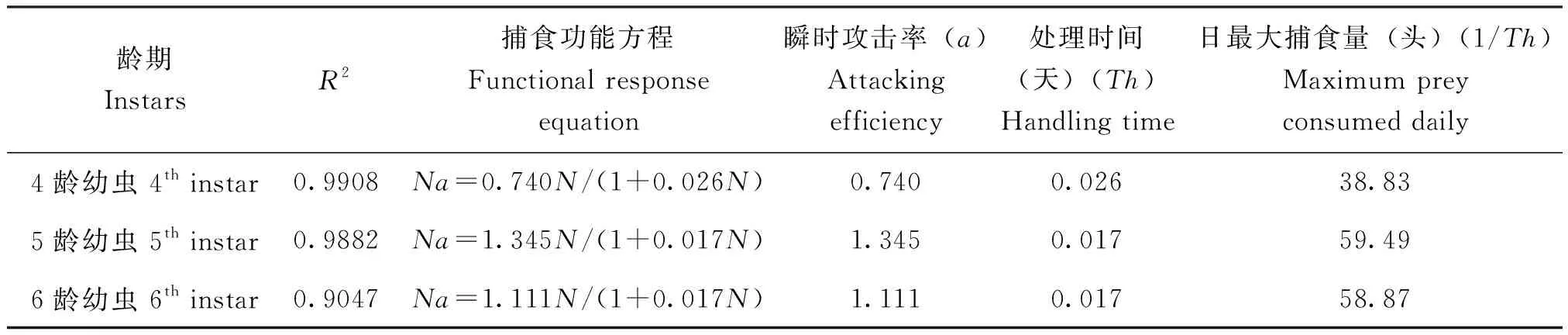
表2 草地贪夜蛾不同龄期幼虫对3龄幼虫的捕食功能反应Table 2 Functional responses of Spodoptera frugiperda at different stages to the third instar larvae
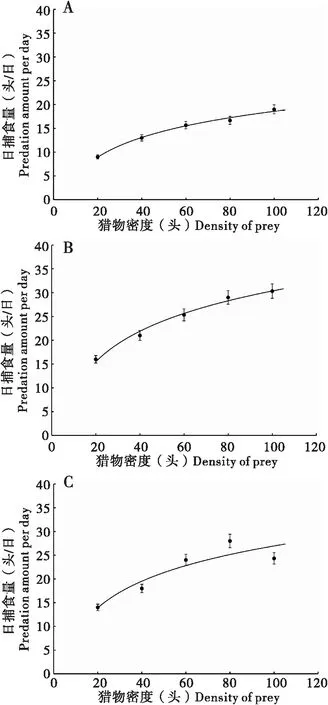
图2 草地贪夜蛾不同龄期幼虫对3龄幼虫的捕食功能反应Fig.2 Functional responses of Spodoptera frugiperda at different stages to the third instar larvae注:A、B、C分别代表草地贪夜蛾4龄、5龄、6龄幼虫对2龄幼虫的捕食功能反应, 图中数值为平均值±标准误。Note: A, B and C showed the functional responses of S.frugiperda at the fourth instar, the fifth and the sixth instar to the second instar larvae respectively, data were means±SE.
2.3 不同龄期幼虫间最大理论捕食量
不同龄期幼虫体重如表3所示,通过利用Holling Ⅱ型圆盘方程进行模拟,得出3、4、5、6龄幼虫对2龄幼虫和4、5、6龄幼虫对3龄幼虫的最大理论捕食量,利用捕食量和体重的关系计算出每个龄期幼虫的最大理论捕食质量,并做出散点图(图3),拟合方程为y=47.231x0.4591。各个龄期间的最大理论捕食质量如图3所示,根据各龄期的体重换算出相应的捕食量。
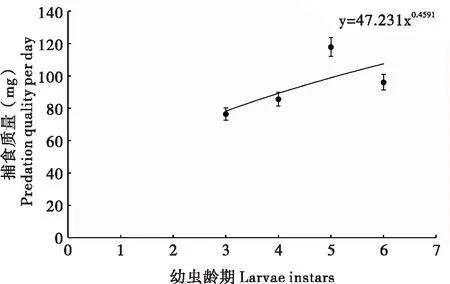
图3 草地贪夜蛾不同龄期幼虫的最大捕食质量Fig.3 The maximum predation quality of larvae at different instars of Spodoptera frugiperda
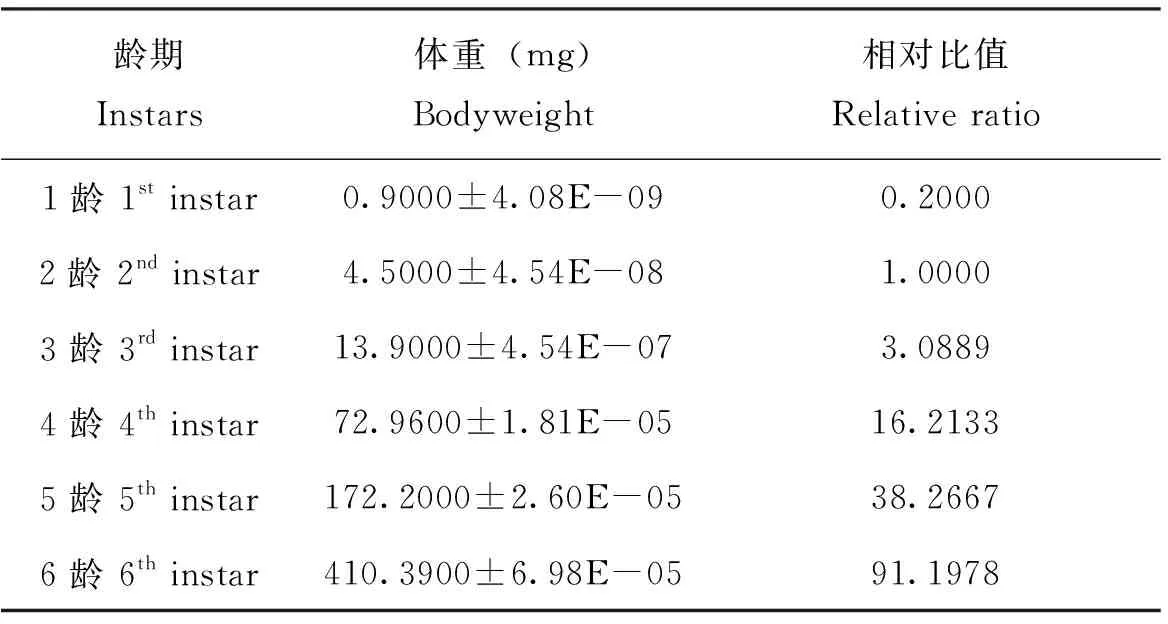
表3 草地贪夜蛾不同龄期幼虫体重Table 3 Larvae body weight of Spodoptera frugiperda at different stages
根据拟合方程y=47.231x0.4591及龄期与体重关系,计算出各龄期对应的捕食量如表4所示,6龄幼虫对低于其龄期的幼虫的捕食量依次为1龄(494.42头)> 2龄(107.52头)> 3龄(34.81头)> 4龄(6.63头)> 5龄(2.81头);5龄幼虫对低于其龄期的幼虫的捕食量依次为1龄(446.28头)> 2龄(98.88头)> 3龄(32.01头)> 4龄(6.10头);4龄幼虫对低于其龄期的幼虫的捕食量依次为1龄(391.06头)> 2龄(89.26头)> 3龄(28.90头);3龄幼虫对低于其龄期的幼虫的捕食量依次为1龄(324.64头)> 2龄(78.21头)。由于没有直接观察到3龄以下幼虫存在自相残杀现象,所以3龄以下幼虫对同类的捕食不作考虑。
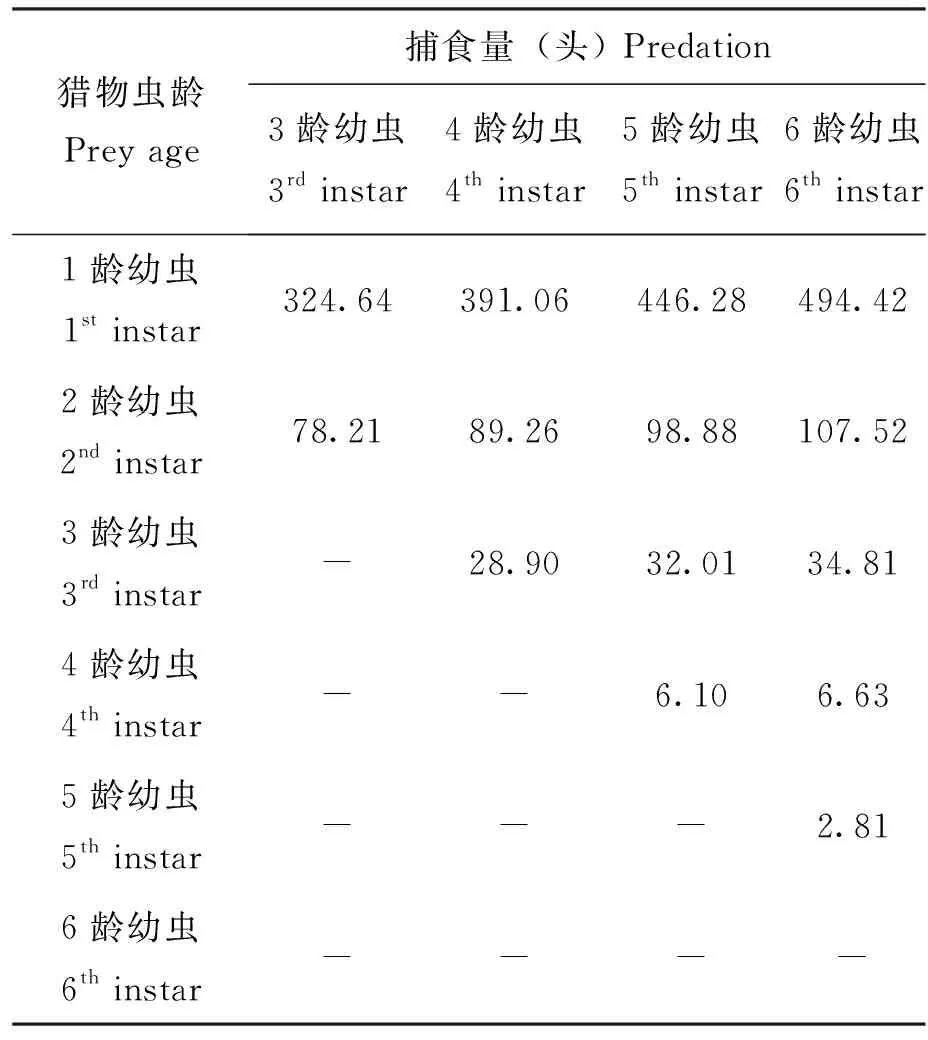
表4 草地贪夜蛾不同龄期幼虫间最大理论捕食量Table 4 The maximum theoretical predation between different instar larvae of Spodoptera frugiperda
3 结论与讨论
自相残杀行为是种群内捕食者和被捕食者相互作用的结果,在种群动态和种群结构中扮演着重要的角色(Heetal., 2021)。自相残杀通常是对食物短缺、高密度、同种个体之间的大小差异(Porrettaetal., 2018)、环境温度(Startetal., 2018)甚至是饲养盒材料差异的反应(Okanoetal., 2018)。本研究明确了草地贪夜蛾在食物充足时仍会对同类幼虫进行捕食,采用Holling Ⅱ型圆盘方程进行拟合,确定了3、4、5、6龄幼虫对2龄幼虫和4、5、6龄幼虫对3龄幼虫的最大理论捕食量。结果表明,草地贪夜蛾3、4、5、6龄幼虫对2龄幼虫的捕食量大小依次为5龄>6龄>4龄>3龄;4、5、6龄幼虫对3龄幼虫的最大理论捕食量大小依次为5龄>6龄>4龄。草地贪夜蛾幼虫对同类低龄幼虫的捕食量先随着龄期的增加而增加,后又显现出下降趋势,6龄幼虫的捕食量低于5龄幼虫,这可能和6龄幼虫其本身活性不高准备化蛹有密切关系,这一现象和已知研究的甜菜夜蛾的种内捕食基本一致(Soniaetal., 2010)。通过分析得知,草地贪夜蛾在4龄以前,捕食量随着猎物密度的增加而呈现缓慢增长,4龄之后,捕食量随着猎物密度的增加而快速增加,表明草地贪夜蛾种内捕食与龄期有着极大关系。邓盼等(2015)在对棉铃虫自相残杀习性与其龄期和食物营养关系的研究中表明棉铃虫自相残杀主要发生在4龄和5龄幼虫之间,而非3龄和6龄幼虫。同样,Elvira等(2010)通过比较低密度和高密度下5龄和6龄甜菜夜蛾的自相残杀率,发现5龄幼虫自相残杀率高于6龄幼虫。而本研究则是通过Holling Ⅱ型圆盘方程模拟在密度无限增大的情况下,可得出理论最大捕食量,其R2值均为0.9以上,但是现实情况下密度是有限的,其最大值可能远远小于理论值。此外,本研究只是确定草地贪夜蛾对不同密度下低龄同类幼虫的最大捕食量,但喂食不同玉米叶部位、寄主种类和温度等因素是否会影响其对低龄幼虫的捕食量还需进行深入研究。
本试验利用各龄期幼虫之间体重的比值,结合3、4、5、6龄幼虫对2龄幼虫的捕食量,计算出不同龄期幼虫的最大捕食质量,从而得出草地贪夜蛾任意两个不同龄期之间高龄幼虫对低龄幼虫的捕食量。通过对高龄幼虫对低龄幼虫的捕食行为进行解析,试探明草地贪夜蛾幼虫的种内自残行为规律,其中龄期和低龄幼虫的密度是影响其自残行为的两大重要因素。同时还要进一步的设计实验,详细的探究出草地贪夜蛾幼虫的种间自残行为规律,为更好地理解种内自残现象背后的生态意义和生理机制,可能有助于开发新的策略来控制害虫。
