外源茉莉酸甲酯对镉污染下草地早熟禾种子萌发及幼苗生长的影响
2022-07-08马志强种培芳
马志强, 史 毅, 种培芳
(甘肃农业大学林学院, 甘肃 兰州 730070)
随着全球气候变化和世界各国经济的快速发展,重金属污染已成为日益凸出的环境问题,尤其以土壤污染更为严重。据《全国土壤污染状况调查公报》报道[1],中国的土壤中重金属点位超标率已达16.1%,而镉以7.0%的点位超标率位居第一。
镉胁迫会降低种子发芽率,抑制植物光合作用与生长发育等[2],造成幼苗根茎长度、面积及数量下降[3]。当土壤中镉含量为200 mg·kg-1时,萝卜(RaphanussativusL.)的茎块、根长度、鲜重和干重明显下降[4]。土壤中的Cd2+进入植物体后,会在植物的各器官之间进行转移,进而累积到果实、籽粒中,其产量和质量会被影响[5]。为降低土壤中Cd2+对植物的影响,对镉污染土壤的修复成为了当前学者关注的热点之一。
为有效应对土壤镉污染,现有修复技术中基本可分为物理、化学及新兴的生物修复三类。其中物理、化学修复手段具有周期短、见效快、易推广等优点被广泛利用,但该技术实施的工程量大,实施所需经费较高,并且易对土壤造成二次污染,在一定程度上其推广利用被限制[6]。而重金属土壤的生物修复(Bioremediation)主要包括植物修复与微生物修复,因具有投资低、修复效果好、不易产生二次污染等优点,日益为人们所重视[6]。
刘倩等[7]将无性系金丝柳(Salixxaureo-pendula)作为镉污染土壤的修复品种,发现其对Cd污染土壤具有较强的修复能力,但其作为落叶乔木,具有生长速度缓慢、培育周期较长及后期维护成本较高等缺点,不适宜于大规模的治理镉污染土壤。而草坪草因具有生长速度较快,生物量较大、可反复修剪、修复效率快等特点的同时兼具景观效应,已在修复土壤重金属污染的领域占据重要地位[8]。已有研究将高羊茅(Festucaarundinacea)[8]及黑麦草(Loliumperenne)[9]作为镉污染土壤的修复草种,通过对植物的形态和相关生理指标进行分析,从而确定其对Cd污染土壤的修复效果。而草地早熟禾(Poapratensis)作为常见的多年生冷季型禾本科(Poaceae)草本植物,具有抗寒[10]、抗旱[11]等特点,已成为常见的建植草坪种之一,且草地早熟禾具有生物量大、根系发达等优点,具备成为治理镉污染土壤草种的潜力,但将草地早熟禾作为治理镉污染土壤草种的相关报道较少。
为增强草地早熟禾幼苗的抗镉能力,提升草地早熟禾对镉污染土壤的治理效果,施加外源缓解剂已作为当前的常用手段之一。现有的研究中,已有将油菜素内酯[12]、一氧化氮[13]、柠檬酸和EDTA等[14]作为缓解剂来增强草地早熟禾对镉的抗性,提升其对镉的吸附能力。茉莉酸甲酯(MeJA)作为茉莉酸类(jasmonates,JAs)[15]的一种,提自素馨花(Jasminumgrandiflorum)香精油中,具有挥发性,易在空气中传播,在植物间的通信联络中起重要作用[15],而茉莉酸类同样可以调控植物对非生物胁迫的反应[16],使用MeJA作为缓解剂,研究其对镉胁迫下草地早熟禾体内镉含量的影响具有现实意义。当前将草地早熟禾作为治理土壤重金属污染的草种研究中,多为单一地对生理指标进行分析[13,17],从而分析其对镉污染的防治效果,而植物-土壤作为一个共同体,对于外源缓解剂或镉胁迫的反应是相互关联的,土壤中的镉含量更是可以最直观的反映草地早熟禾对镉污染土壤的修复状况,而种子萌发期作为草地早熟禾在镉胁迫环境下生长发育过程中最脆弱的阶段[18],萌发期的生长状况对草地早熟禾后期的生长发育及镉污染防治效果起至关重要的作用,但是目前将种子萌发部分-幼苗生长部分-土壤镉含量三者进行结合,分析茉莉酸甲酯对草地早熟禾修复镉污染土壤效果的影响却鲜有报道。
本文以镉为污染源,外源MeJA作为缓解剂,草地早熟禾作为修复植物,通过分析草地早熟禾的种子萌发数据,对比溶液与土壤中生长的草地早熟禾体内的镉含量,计算土壤中的镉含量及富集系数,主要解决以下问题:(1)MeJA对镉污染下的草地早熟禾种子萌发是否具有缓解作用?(2)如果有,施用缓解剂的最适浓度应如何选择?(3)MeJA能否调控草地早熟禾修复Cd污染土壤的能力?
1 材料与方法
1.1 草地早熟禾种子
供试草地早熟禾种子品种为午夜(Poapratensis‘Midnight’),选种依据为鲜靖苹等筛选出[17]的耐Cd草种,购于兰州市金惠种子经营部。
1.2 MeJA及镉浓度设置
使用CdCl2和蒸馏水配制Cd2+溶液,预实验中,Cd2+溶液浓度设置为50,75,100,150和200 μmol·L-1共5个浓度梯度,Cd2+浓度为50 μmol·L-1和75 μmol·L-1时,草地早熟禾种子萌发被促进,种子萌发数较CK分别提升27.24%和16.88%,Cd2+浓度为150和200 μmol·L-1时,相较于CK,种子萌发数分别下降42.71%和55.71%,草地早熟禾种子萌发受严重抑制,Cd2+浓度为100 μmol·L-1时,种子萌发数较CK下降35.23%,对草地早熟禾种子的生长胁迫的程度较轻,遂选取Cd2+浓度为100 μmol·L-1为胁迫浓度。预实验所得结果见图1。
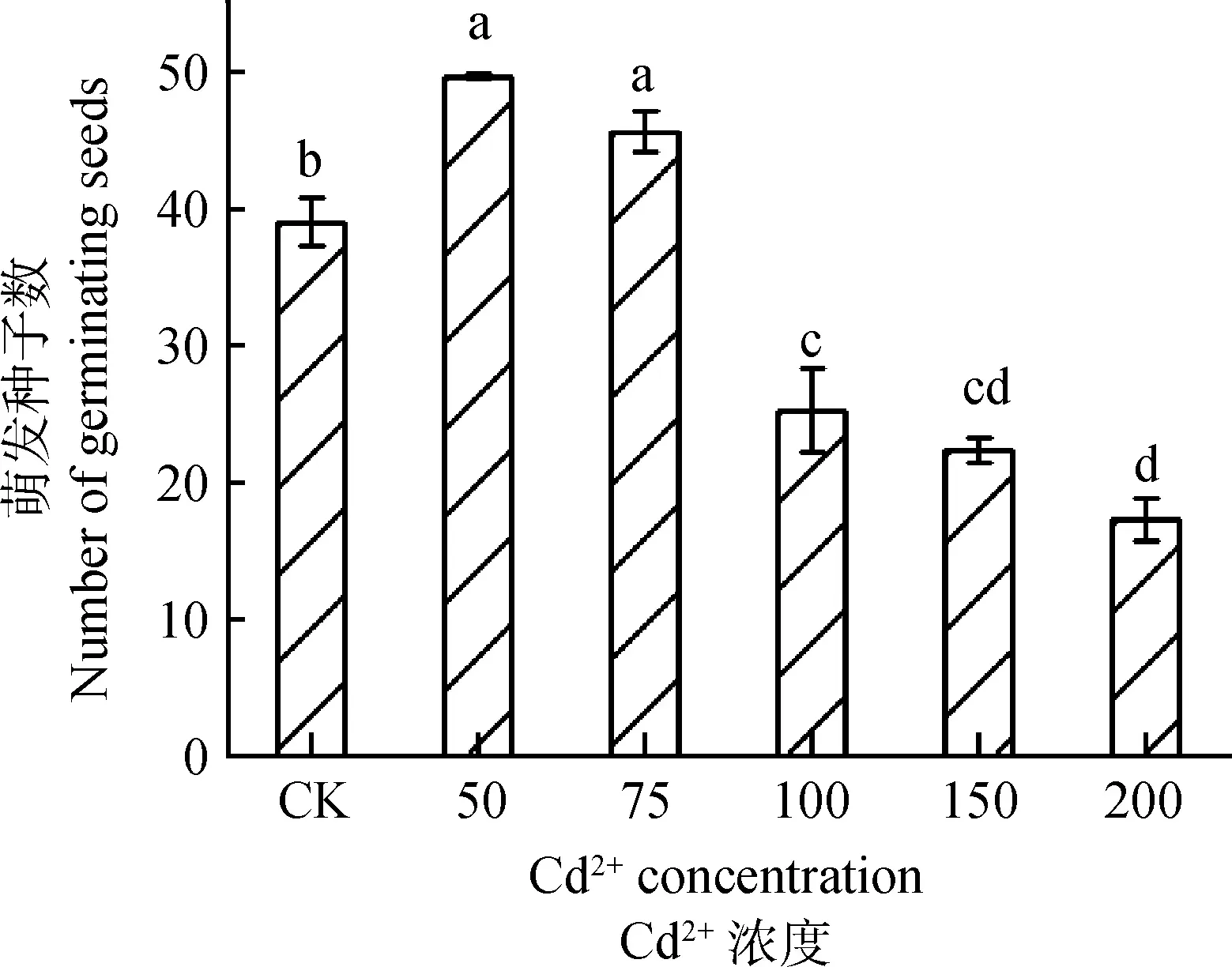
图1 不同Cd2+浓度对草地早熟禾种子萌发的影响Fig.1 Effects of different Cd2+ concentrations on Seed Germination of Poa pratensis
选定Cd2+浓度后,预实验中设定MeJA浓度为10,20,25,30,50,75,100,125,150 μmol·L-1共9个浓度梯度,MeJA溶液用乙醇溶解后加蒸馏水进行配制,发现施用浓度为10,25,50,75,100 μmol·L-1的MeJA溶液后,草地早熟禾发芽指数等差异明显,MeJA浓度高于100 μmol·L-1时,草地早熟禾生长均呈下降趋势。遂将MeJA溶液浓度梯度设置为0,10,25,50,75,100 μmol μmol·L-1,镉胁迫设置为Cd2+浓度100 μmol·L-1的镉离子溶液,CK全过程施用1/2Hoagland溶液进行培养,共7个处理,每个处理设3个重复,见表1。
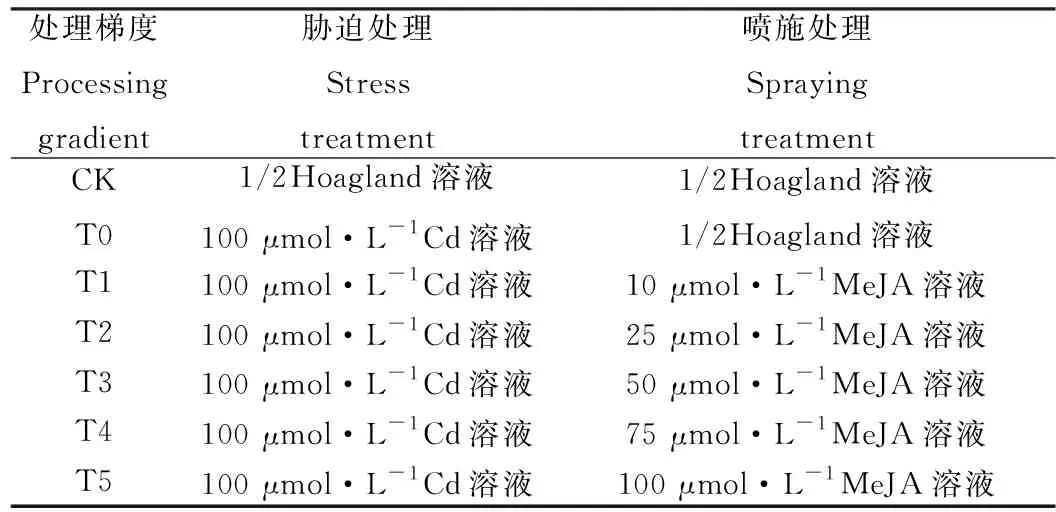
表1 镉处理浓度及外源MeJA施用浓度Table 1 Cadmium treatment concentration and exogenous MeJA application concentration
1.3 种子萌发的实验
破除种子休眠后,对种子进行消毒,晾干后进行处理,发芽方法选取为纸上发芽法[19],选取90 mm的培养皿21个,之后将浸有不同浓度梯度Cd处理液的滤纸平铺在这些培养皿中,滤纸上放置之前处理过的干燥种子,每个培养皿中均匀放置50枚种子,每个处理设置3个重复,培养温度设定为白天28℃,夜晚22℃,时间设定为白天16 h,夜晚8 h,光照度2 000 lx,每个培养皿称重之后记录,每天根据称重记录添加蒸发的蒸馏水,发芽标准为种子萌发并长成正常幼苗,3 d后在滤纸上添加MeJA溶液,喷施顺序添加蒸馏水后再施用MeJA溶液,添加容量为2 mL。每天对胚芽长、胚根长进行记录,25 d后停止发芽实验,测定并计算相关指标,相对含水量测定采用张志良[20]的方法,干物质含量测定参考文献[17]的方法。
1.4 幼苗的培养实验
选取花盆(直径23 cm,高14 cm)共21个,每个处理梯度设3个重复,每个花盆内用蛭石填满,将草地早熟禾种子均匀播撒在这些花盆中后用薄土覆盖,土壤选取为有机质含量较高的肥沃黑土,之后每天用少量蒸馏水对种子进行一次浇灌,室温培养,间苗时间段为幼苗地上长度约1 cm时,间苗后密度为每盆20株幼苗,分布状况为不集中地均匀分布。间苗前不添加胁迫,间苗后改用1/2Hoagland营养溶液继续培育,将CdCl2·1/2H2O加入营养液中用来配制浓度为100 μmol·L-1的含镉营养液胁迫幼苗生长,CK全过程采用1/2Hoagland营养溶进行培育。采用叶面喷施法在草地早熟禾幼苗叶片上均匀喷施MeJA溶液,喷施至叶面滴水,喷施间隔时间为3 d,CK叶片表面喷施蒸馏水。Cd2+溶液浇灌于土壤表面,喷施时间间隔为3 d,施用容量为50 ml。Cd2+溶液与MeJA溶液隔天喷施。经过15 d的处理后,采集样本,样本在采集时注意减掉叶尖及根部,选取其相同部分作为试验材料。
1.5 苗长、根长和鲜重的测定
发芽苗长、胚芽及胚根长采用Epson Expression 11000XL扫描,使用winRHIZO对扫描所得数据进行计算。鲜重用吸水纸吸干水分后称取重量,3次重复。
1.6 Cd指标的测定
使用火焰原子吸收分光光度计对[21]叶片及根中镉含量进行测量,溶液及土壤中镉含量的测定方法为石墨炉原子吸收光谱法(GFAAS),具体测定方法分别参照文献[22-23]。对草地早熟禾的根叶部分的鲜重和干重分开称重并记录,用尺子对地上部分的伸长长度进行测定,伸长长度测定时在处理前对草地早熟禾统一修剪并记录长度,处理7 d后测量并记录长度。
1.7 相关指标计算
式中:Gt为第t天的发芽种子数;Dt为对应的发芽种子天数。
式中:D为从种子置床起算的天数,有的规定置床之日为0,有的规定置床之时为1;
n为相应各天的发芽粒数
(5)活力指数(Vi)=发芽指数(Gi)×幼苗重量(S)
(7)转移系数=草地早熟禾叶中的Cd浓度÷草地早熟禾根中镉浓度
(8)富集系数=草地早熟禾体内Cd浓度÷土壤中Cd浓度
(9)伸长长度=Cd处理前草地早熟禾地上部分高度-Cd处理前草地早熟禾地上部分长度。
式中:RWC(relative water content)为相对含水量,Wf(fresh weight)为鲜重,Wt(saturated weight)为叶片饱和重,Wd(dry weight)为干重
式中:m0为烘干2 h后的植物恒重,m1为20株幼苗洗净剪碎后放入铝盒后的重量,m2为铝盒内样品3次烘干后的恒重。
1.8 数据处理
采用SPSS26.0分析数据并用“平均值±标准误差”表示测定结果,分别对镉胁迫下施用不同浓度的MeJA处理进行单因素方差分析,绘图采用Origin2018作图。
2 结果与分析
2.1 外源茉莉酸甲酯对镉胁迫下草地早熟禾种子萌发的影响
2.1.1茉莉酸甲酯对镉胁迫下草地草地早熟禾种子萌发的影响 图2显示了施用外源MeJA后草地早熟禾发芽数据的变化趋势。图2A中,T0处理时发芽率与发芽势较CK分别下降了65.67%与74.31%,差异极显著(P<0.01),发芽率与发芽势除CK外,均在T3处理时升至最高点,较T0处理提升117.39%和114.29%,差异显著(P<0.05);图2B中,平均发芽速度在T0处理时明显上升,T4处理时,平均发芽速度较T0处理显著下降4.22%(P<0.05),与T3处理梯度差异不显著(P>0.05),活力指数在CK时最高,T0处理时,活力指数较CK下降了89.18%,差异极显著(P<0.01),T3处理时,活力指数较T0处理显著上升492.63%(P<0.05),与T4处理梯度间的差异不显著(P>0.05)。
图2C中,草地早熟禾发芽指数在T0处理时最低,较CK降低73.08%,差异显著(P<0.05),除CK外,发芽指数在T3处理时升至最高,较T0处理提升了184.66%,差异显著(P<0.05),T1、T2和T5处理梯度间差异不显著;种子死亡率在T0处理时最高,较CK提升84.61%,差异显著(P<0.05),T3处理较T0处理显著降低51.92%(P<0.05)。
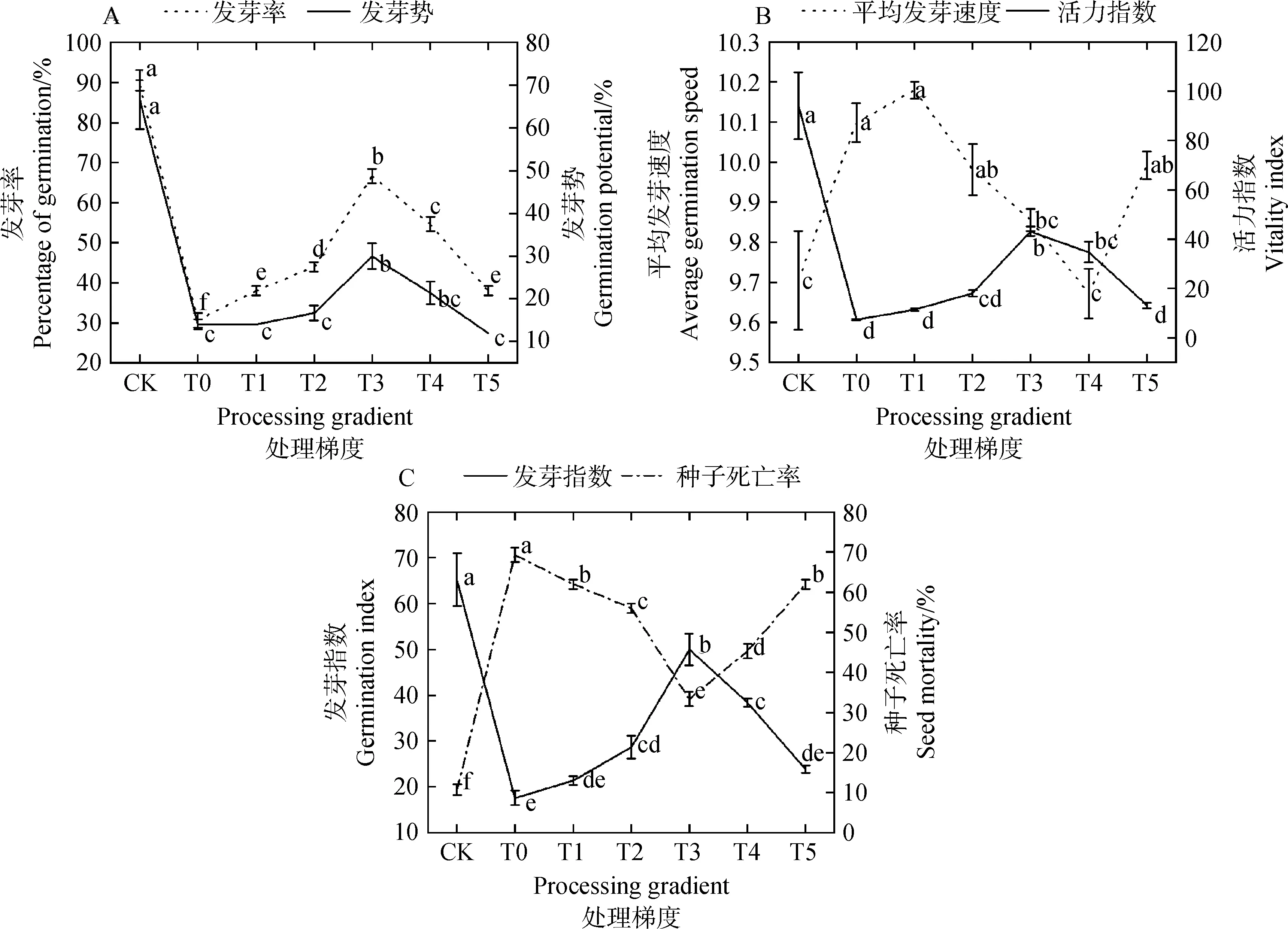
图2 茉莉酸甲酯对镉胁迫下草地早熟禾种子萌发的影响Fig.2 Effect of Methyl Jasmonate on seed germination of Poa pratensis under cadmium stress注:图中不同小写字母表示数据平均值之间显著差异(P<0.05),下同Note:Different lowercase letters indicate significant difference between the average values of data at the 0.05 level,the same as below
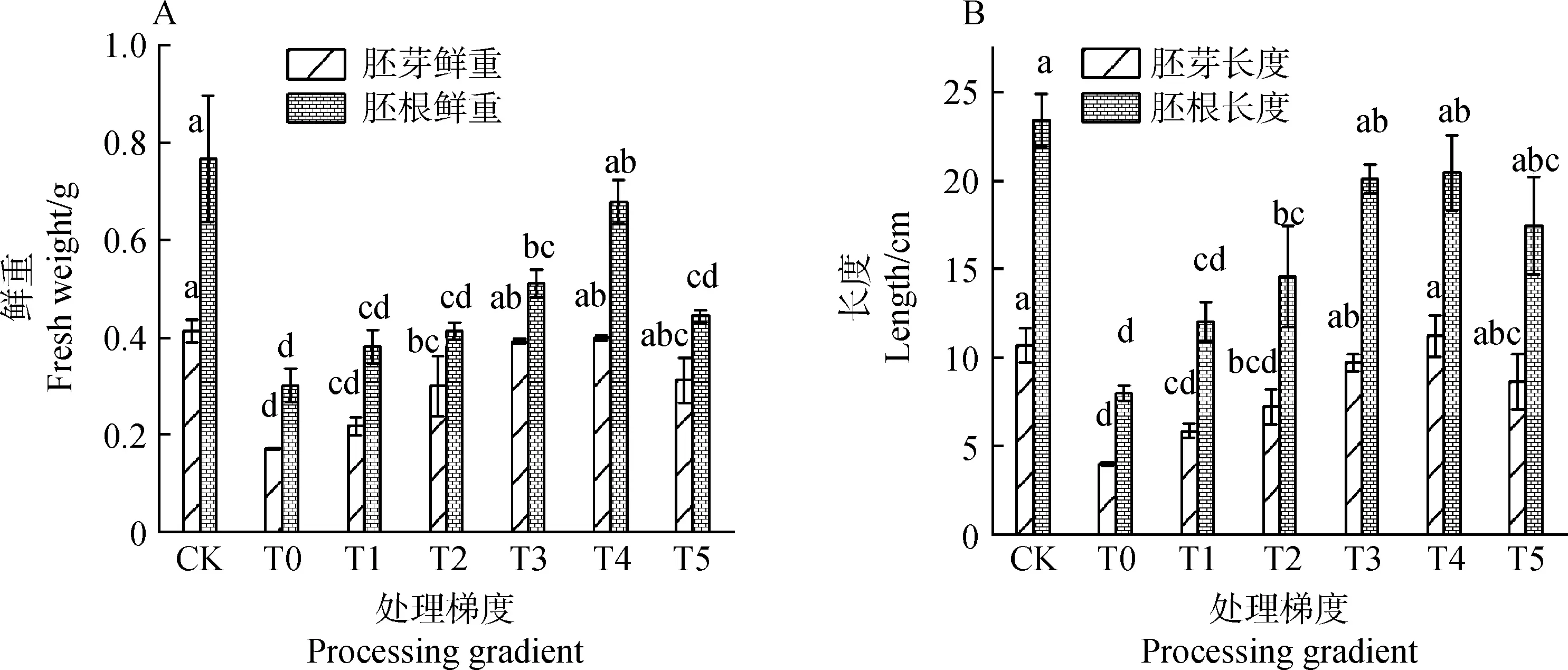
2.1.2茉莉酸甲酯对镉胁迫下草地早熟禾重量、长度、相对含水量和干物质量的影响(种子萌发) 图3为种子萌发中草地早熟禾重量及长度的变化趋势图。图3A中,处理梯度为T3和T4时,胚芽鲜重较T0处理增加了128.54%和132.42%,差异显著(P<0.05),T3较T4处理差异不显著(P>0.05),胚根鲜重较T0处理分别提升69.21%和124.73%,差异显著(P<0.05);图3B中,除CK外,T3和T4处理下胚芽长度及胚根长度较长,较T0处理,胚芽长度分别提升了142.91%和180.55%,胚根长度分别提升了151.78%和156.42%,差异显著(P<0.05),两梯度间胚根长度差异不显著(P>0.05);图3C中,T0处理下,相对含水量和干物质量较CK显著下降了8.76%和8.52%(P<0.05),除CK外,T4处理下,相对含水量升至最高,较T0处理显著提升7.36%(P<0.05),处理梯度为T2~T4时,3个处理梯度间的干物质量差异不显著(P>0.05),且与CK差异不显著(P>0.05)。
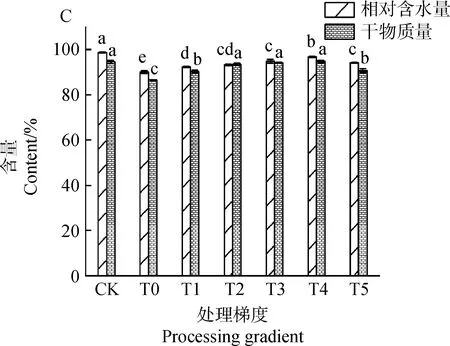
图3 茉莉酸甲酯对镉胁迫下草地早熟禾重量、长度、相对含水量和干物质量的影响(种子萌发)Fig.3 Effects of Methyl Jasmonate on weight,length,relative water content and dry matter quality of Poa pratensis under cadmium stress (seed germination)
2.2 外源茉莉酸甲酯对植物镉含量及土壤指标的影响
2.2.1外源茉莉酸甲酯对草地早熟禾体内镉含量的影响 表3体现了土壤中生长的草地早熟禾,在施用MeJA后其叶片及根系镉含量的变化趋势。发现施用MeJA后,叶片及根系镉含量上升,转移系数增加,土壤镉含量下降,处理梯度为T4时,叶片镉含量和转移系数最高,较T0处理显著提升112.80%和62.07%(P<0.05),根系镉含量也在此处理梯度时最高,较T0处理提升32.45%,差异显著(P<0.05),与T2、T3和T5处理时根系镉含量差异不显著,土壤镉含量在T3处理时最低,较T0处理下降23.38%,差异显著(P<0.05),相较于T2,T3和T5处理时土壤镉含量差异不显著。
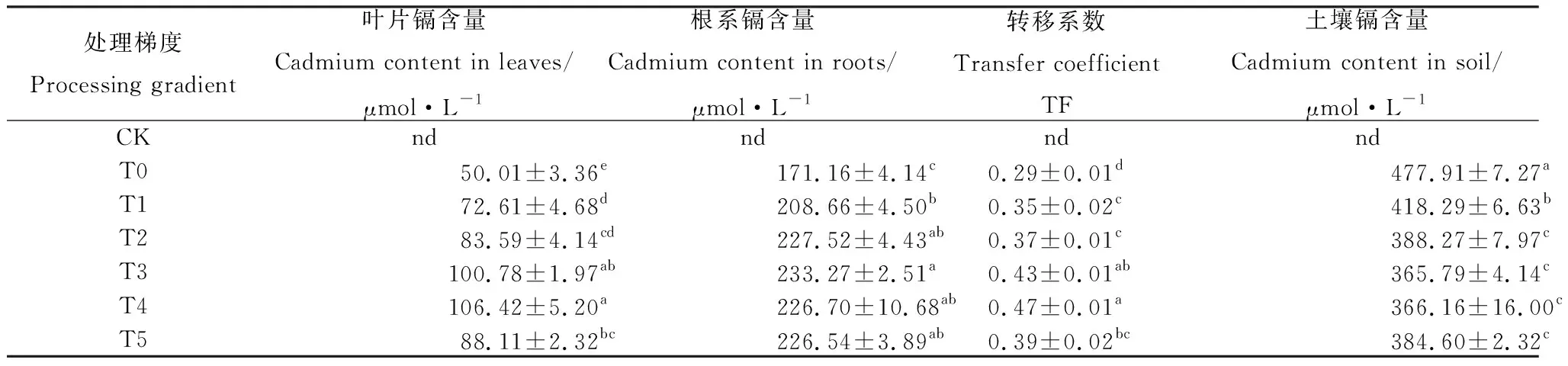
表3 外源茉莉酸甲酯对镉胁迫下草地早熟禾幼苗根系及叶片镉含量的影响Table 3 Effects of Exogenous Methyl Jasmonate on cadmium content in roots and leaves of Poa pratensis seedlings under cadmium stress
2.1.3茉莉酸甲酯对镉胁迫下草地早熟禾体内镉含量的影响(种子萌发) 由表2可知,T4处理下,胚芽镉含量、转移系数和溶液中的镉含量升至最高,较T0处理分别提升36.75%,63.33%和23.10%,而胚根中镉含量在T4处理时较T0处理下降了16.88%,均差异显著(P<0.05)
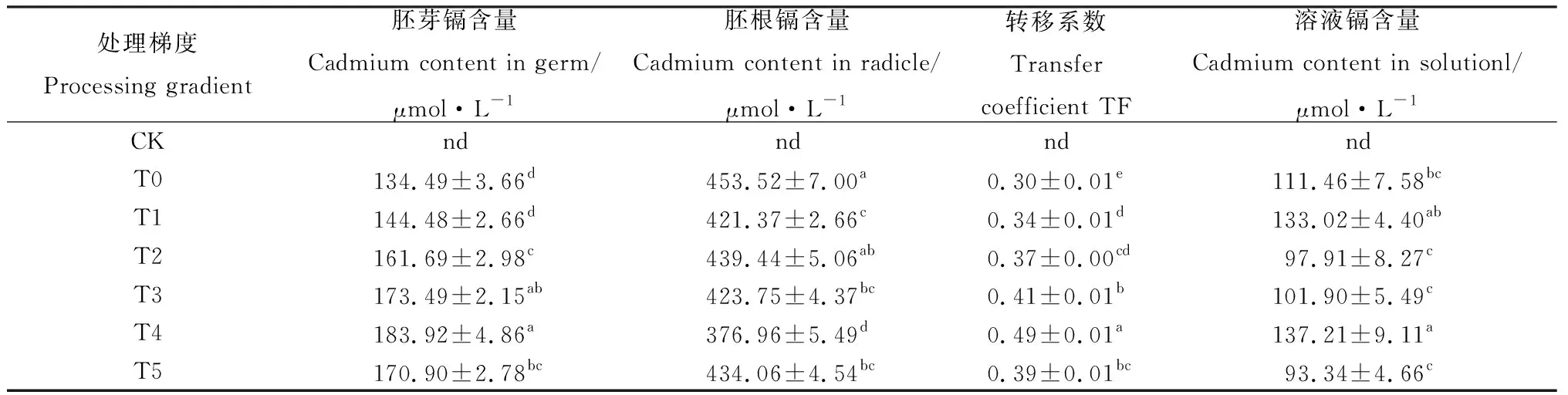
表2 外源茉莉酸甲酯对镉胁迫下草地早熟禾胚根及胚芽镉含量的影响(种子萌发)Table 2 Effects of Exogenous Methyl Jasmonate on cadmium content in radicle and embryo of Poa pratensis under cadmium stress (seed germination)
2.2.2外源茉莉酸甲酯对草地早熟禾伸长长度、根系长度及重量的影响 图4为镉胁迫下草地早熟禾在施用MeJA后重量及长度的变化趋势。图4A发现,T0处理下伸长长度及根系长度较CK显著下降49.95%和40.10%(P<0.05),草地早熟禾幼苗处理梯度为T4时,较T0处理其伸长长度提升了60.42%,差异显著(P<0.05),除CK外,根系长度T4处理时最长,较T0处理显著提升49.96%(P<0.05),与CK、T3处理时根系长度的差异不显著(P>0.05);图4B中,除CK外,T4处理下,叶片及根系鲜重最重,其中叶片鲜重较T0处理提升51.85%,差异显著,根系鲜重T4处理时较T0处理差异不显著(P>0.05),提升了12.92%,较其他处理梯度下根系长度的变化差异也不显著(P>0.05)。
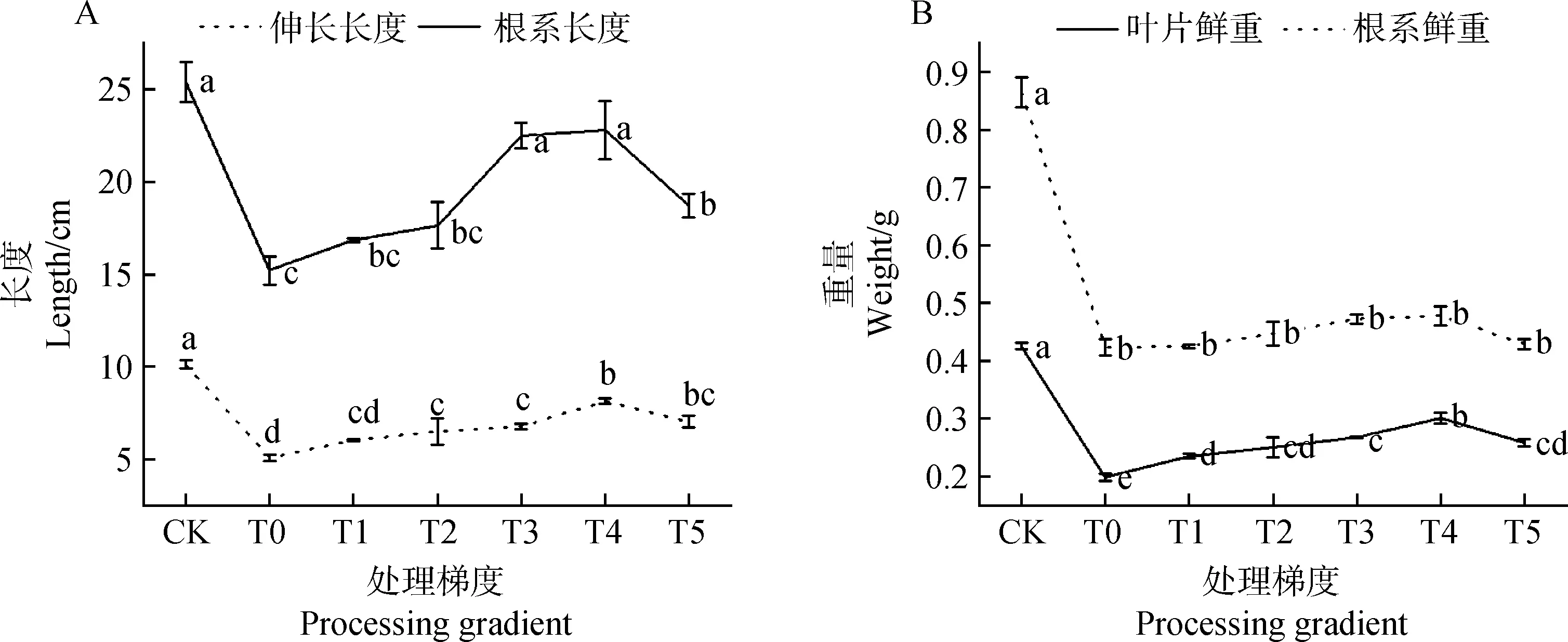
图4 茉莉酸甲酯对镉胁迫下草地早熟禾伸长长度、根系长度及鲜重的影响(土壤生长)Fig.4 Effects of Methyl Jasmonate on elongation length,root length and fresh weight of Poa pratensis under cadmium stress (soil growth)
2.2.3外源茉莉酸甲酯对土壤酸碱度和草地早熟禾富集系数的影响 图5中,施用MeJA后,土壤pH值较T0处理明显下降,富集系数明显上升,处理梯度为T3~T4时,pH值较T0处理分别显著下降7.23%和7.36%,差异显著(P<0.05),两者间差异不显著(P>0.05),富集系数较T0处理分别显著上升97.14%和97.80%(P<0.05),两者间差异不显著(P>0.05)。
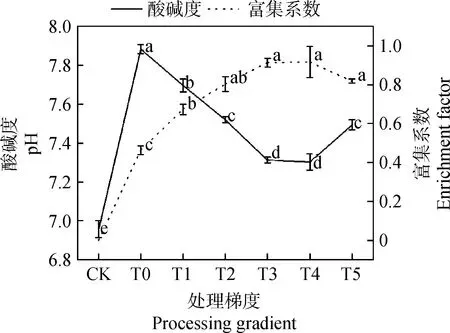
图5 外源茉莉酸甲酯对草地早熟禾修复镉污染土壤的影响Fig.5 Effect of Exogenous Methyl Jasmonate on the remediation of cadmium contaminated soil by Poa pratensis
3 讨论
3.1 MeJA对镉胁迫下草地早熟禾种子萌发的影响
植物种子萌发是一个非常复杂的过程,在萌发过程中受许多因素的影响。种子的发芽率反映了种子的总体发芽能力,发芽指数、活力指数则体现了种子的萌发速度、整齐程度及种子的萌发质量[24]。本研究中,T0处理时,种子发芽率、发芽指数与活力指数均相较于CK下降明显,与王勇[25]等人的研究结果不一致,推测有两个原因,一是只有少部分种子的种皮阻挡了Cd对种子的入侵,其余种子在萌发过程中,受Cd2+胁迫,表现为种子萌发的停止;二是种子品种的选择不同,导致种子萌发的数据差异较大。施用MeJA后,发芽率、发芽势与活力指数均在T4处理时最高,但发芽势在施用MeJA后,与T0处理相比不明显,活力指数的变化趋势也较为平缓,可能是发芽势体现了13 d内的种子活力程度,MeJA施用时间较短,防御基因的组成型表达并未形成,发芽率表现了种子25 d内萌发的活力程度,T4处理时发芽率与T0处理差异明显,说明MeJA累积增加后,种子体内的防御基因的组成型表达形成,进一步证明了MeJA的累积有利于缓解非生物胁迫对种子萌发造成的损害,Seo[26]的观点也证明了此推测的合理性。本研究中,种子死亡率最高点出现在T0处理时,T3处理明显降低了种子死亡率,表明施用适宜浓度的MeJA可以缓解种子因镉胁迫而造成的死亡率上升。平均发芽速度可以作为种子萌发期耐性的负向指标[27],即平均发芽速度愈高,种子萌发状况愈差,本研究中,T0处理时,平均发芽速度最高,施用MeJA后发芽速度降低。结合上述分析,表明喷施低浓度的MeJA作为非生物因素促进了草地早熟禾种子萌发,而过高浓度的MeJA对种子萌发起抑制作用,与前人[28]研究结果相似。
3.2 MeJA对镉胁迫下草地早熟禾重量与长度的影响
干重、鲜重、胚芽长与胚根长直观地反映了植物体的生长状况,植物伸长长度反映植物生长发育程度同时,也反映植物对环境的耐性[29]。草地早熟禾在添加Cd溶液后,其干、鲜重、胚芽、胚根长及伸长长度均相较CK差异明显,可能此过程中Cd作为植物生长发育的有害元素[30],诱导活性氧产生的同时,使得膜质过氧化,Cd2+与生长细胞相结合,对生物大分子造成损伤,光合作用等生理过程发生紊乱,使得植株的生长被严重抑制[31]。施用MeJA后,较T0处理,植物的干重、鲜重、胚芽长、根系长均上升,对修复重金属污染的土壤而言,保持较高的地上部分长度是提高修复效率的根本措施之一[8],证明MeJA可以缓解镉胁迫对草地早熟禾生长带来的损害。推测草地早熟禾在应对镉胁迫时,茉莉酸合成途径(ctadecenoic途径)被影响,植物体内源JA含量上升,防御基因中多种表达基因上调[32],而当植物体内Cd含量过高时,植物体产出的内源JA含量不足以抵抗Cd对植物带来的损害,使得Cd2+进入植物体内,对草地早熟禾产生毒害作用,表现为Cd对植物的“低促高抑”[33]。此时加入适量MeJA后,相较于T0处理下植物体内产生的内源JA,外源MeJA的加入进一步增强了茉莉酸合成途径的信号转导,植物还原系统的活性提升,提高了植物的抗性。而土壤培养环境下,草地早熟禾的重量及长度数据基本高于种子萌发过程中的草地早熟禾数据,原因可能是土壤中的腐殖质为植物的生长提供了养分,或者是土壤为植物提供大部分生长过程中所需的营养元素[34],使得土壤中生长的草地早熟禾优于培养皿中培育的草地早熟禾。
3.3 MeJA对镉胁迫下草地早熟禾相对含水量和干物质含量的影响
相对含水量(RWC)是组织水重占饱和组织水重的百分率,反映了细胞壁的紧张程度[35],同时也反映了叶片水分与蒸腾之间的平衡关系[36]。T0处理时,草地早熟禾的相对含水量下降,与CK差异明显,原因是Cd2+取代了叶片细胞中的水分子,使得相对含水量下降,亦或是Cd2+破坏了叶片水分与蒸腾间的平衡关系,抑制了草地早熟禾水孔蛋白基因表达,植物水分吸收被抑制。而T2,T3和T5处理下,草地早熟禾相对含水量之间差异不明显,但较T0处理明显上升,推测植物体内在已受Cd2+的破坏后,LOX被激活,LOX使得生成的JAs进一步被激活,使得JAs积累[37],外源MeJA的加入使得更多JAs累积,植物体内分子活力重新被唤醒,细胞壁的紧张程度得到缓解,叶片水分与蒸腾的关系的到了平衡,表现为相对含水量的上升。干物质量为光合作用产物的质量,同时为生物脱水后的质量,反映植物的生长状况。发现施用MeJA后干物质含量较T0处理呈上升趋势,与前人[38]研究结果一致,且T2、T3和T5处理时干物质量与CK差异不大,进一步证明了MeJA对植物的生长有缓解作用,可以缓解非生物胁迫对植物带来的损害。
3.4 MeJA对草地早熟禾体内镉含量的影响
植物根系作为重金属等一些污染物进入植物体内的门户,在应对重金属胁迫及吸收过程中起至关重要的作用[39]。本研究发现,培养皿及土壤中培育的草地早熟禾在相同浓度的Cd胁迫下,地下部性状(根系鲜重及根长)对镉胁迫的敏感程度明显高于地上部性状(叶片鲜重及长度)。草地早熟禾地下部根系的镉含量均大于地上部叶片的镉含量,与铁梅等[40]的研究结果相同,证明草地早熟禾作为Cd污染土壤的修复草种确实具有一定潜力。
转移系数作为反映植物系统中重金属元素由地下部向地上部转移难易程度的指标,可以衡量植物在应对重金属胁迫时的修复能力[41]。本研究中,草地早熟禾在T0处理时,其转移系数在0.29附近,培养皿中培育的草地早熟禾在此时根部Cd浓度最高,而相同的转移系数下,土壤中此时根部的Cd含量较低,推测原因为培养皿中过多的Cd2+只有一小部分通过蒸发等分散出培养环境,其余均被植物所吸收,累积在植物生长点附近,对根部造成损伤[25],而有机质含量较高的土壤对Cd2+具有较好的吸附作用[42],减缓了Cd对植物根系的损伤,减少了植物体中的Cd2+的含量,培养皿中培育的草地早熟禾由于没有土壤吸附Cd2+,大量Cd2+转移进入植物根系,根系鲜重及长度均低于土壤培育的草地早熟禾证明了此推测的合理性。施用MeJA后,转移系数上升,原因可能是茉莉酸甲酯促进了草地早熟禾自身的解毒机制,加快了Cd2+由根部向叶片的转移速度,使叶片中镉含量上升。
3.5 MeJA对土壤pH值及镉含量的影响
富集系数可衡量植物对重金属积累能力强弱,表达为植物中某种重金属的含量与土壤中某种重金属含量的比值,富集系数越大,证明该植物对金属的富集能力越强[43-44]。本文研究发现,T5处理下,草地早熟禾的富集系数为0.4左右,而在施用MeJA后,富集系数较T0处理明显提升,且T2~T5处理时富集系数间差异不明显,证明此处理下的MeJA较好地提升了草地早熟禾的抗逆能力,促进了草地早熟禾对Cd2+的吸收。结合转移系数进行分析后,得出施用此浓度的MeJA使得草地早熟禾能够有效富集土壤中Cd的同时,其体内对Cd的转移能力也被提升。
研究表明,土壤的pH是影响土壤吸附镉的重要因素[45-47],可以控制土壤中的Cd2+的吸附与转移[45],土壤的pH值与Cd2+呈正相关,表现为pH值升高,土壤对Cd的吸附量逐渐增加[46],费志军等[47]的研究也证明了这一点。本研究中,土壤pH值下降时,土壤中的Cd含量下降,T5处理时,土壤pH值上升,土壤中的Cd含量也呈上升趋势,原因可能是适当浓度的MeJA提升了草地早熟禾对Cd2+的吸附能力,表现为土壤中Cd含量下降,土壤pH值下调,而过高浓度的MeJA对草地早熟禾吸附Cd2+起负作用,使得土壤中Cd含量上升,土壤pH值也呈上升趋势。
4 结论
本研究中,T1~T4处理下发芽指标的变化具体表现为发芽率、发芽势和活力指数上升,平均发芽速度下降;草地早熟禾在T1~T4处理时,叶片及根系的长度和重量均呈上升趋势,转移系数与富集系数也呈上升趋势;而土壤中Cd含量与pH值均呈下降趋势,表明适当浓度MeJA可促进草地早熟禾对土壤中Cd的吸收,降低土壤中Cd含量。这些变化趋势的最高点的出现区间基本为T3~T4处理时,表明该浓度的MeJA对草地早熟禾治理镉污染土壤具有促进作用。
