利用SSR分子标记技术分析甜菜多胚授粉系不同个体之间的遗传多样性
2022-05-30李月宏李乔乔吴则东
李月宏,李乔乔,吴则东*
(1.黑龙江大学现代农业与生态环境学院,哈尔滨 150080;2.黑龙江省普通高校甜菜遗传育种重点实验室,哈尔滨 150080)
0 引言
甜菜(BetavulgarisL.)属于藜科、甜菜属的二年生草本植物,是我国北方主要经济作物之一,甜菜是重要的糖料作物,产糖量非常高。甜菜是中温带地区的主要糖料作物,具有耐寒、耐旱、耐盐碱的特性,是一种抗逆性强、适应性广的作物,其可以在干旱和半干旱的地区广泛种植[1]。甜菜属共有12个种,其中属于普通甜菜种中的普通甜菜亚种又可以分为食用甜菜、叶用甜菜、饲用甜菜和糖用甜菜。2020年我国糖用甜菜种植面积约为213千公顷,产量达1198.4万吨。甜菜原产于欧洲的西部和南部沿海地区,从瑞典移植到西班牙,中国大面积引种糖用甜菜始于1906年,甜菜生产前景较好。种质资源是作物的遗传改良和基础研究的物质基础[2]。只有满足作物种质资源足够的数量和优异的质量以及种质研究所需知识的深度,才能使种质资源的利用效率进一步提高,使种业可持续发展[3]。甜菜育种的重要内容之一是优质种质资源的开发利用。通过对甜菜种质资源的识别,可以充分了解每种甜菜种质的表型特征,从而找到更好的甜菜种质并有效利用[4]。
我国甜菜种质资源最早都是从国外引进的多胚种子中进行选择而来,由于大部分甜菜种质资源具有自交不亲和性,引进的种子比较杂合,我国的甜菜育种家们为了适应我国甜菜育种生产的发展和制糖产业的需求,分别在不同地区进行选择。回顾我国甜菜育种的历史进程,自上个世纪50年代以来,为了选育优良品种,衍生出一些育种方法,从系统选择法到混合选择法及母系选择法[5],以及轮回选择法等等,但是这些育种方法选育出的品种均具有一定的异质性,后来开始利用雄性不育作为母本生产杂交种,而父本则是利用多胚授粉系,我国目前使用的甜菜多胚授粉系的主要选择方法就是母系选择法和混合轮回选择的方法,这两种方法选择出的授粉系在不同个体之间会存在一定的异质性,因此我们准备利用分子标记技术鉴定我国多胚授粉系的遗传多样性,也就是同一授粉系不同个体之间的差异性。 在育种过程中为了提高甜菜的育种效率,选择优良的杂交组合,获得优良的创新品种,需要进一步分析种质资源的遗传背景和内部遗传结构,这样可以高效的提高甜菜育种的工作效率,达到育种所需的种质资源品质[6]。
随着DNA分子标记技术的快速发展,农作物品种的鉴定已经转移到基因层面[7]。最常见的分子标记技术包括 RFLP[8]、RAPD[9]、AFLP[10]、ISSR[11]、SSR[12]、SRAP[13]和 SNP[14]。SSR 为简单重复序列(simple sequence repeat),SSR 标记又称为微卫星 (Microsatellites)标记,是由 Moore 等于 1991 年建立的基于 PCR 的一种分子标记技术。SSR技术由于其遗传共显性和技术简单,被广泛用于分析甜菜的遗传多样性,是最常见的分子标记技术之一[15]。本试验利用SSR分子标记技术对甜菜多胚授粉系不同个体之间的遗传多样性进行分析。了解我国多胚授粉系不同个体之间的混杂程度,为将来更好的品种选育打下基础。
1 材料与方法
1.1 实验材料
本研究中所用4份甜菜授粉系(每份授粉系包括50个单株)的编号、名称以及来源详情见表1。
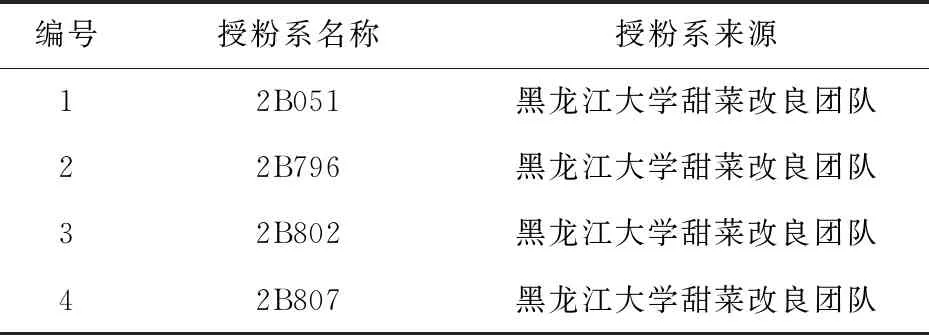
表1 4份甜菜授粉系材料的编号、名称及来源
1.2 实验方法
1.2.1 DNA提取与检测
在甜菜嫩叶时期,采集4份甜菜多胚授粉系材料的新鲜嫩叶,每份授粉系提取50个个体,将采集的甜菜样品按照其编号分别装入PC管中,放入液氮中使其达到液氮的温度,再用震荡研磨机打碎,采用 CTAB 法提取基因组DNA,利用超微量分光光度计测定提取DNA的浓度和纯度。
1.2.2 PCR扩增体系和程序
SSR 引物选用实验室已筛选并验证的多态性好、条带清晰的15对 SSR 引物(表 2),均由上海生工生物工程有限公司合成。
PCR 扩增体系为5 μL,其中包括DNA模板1.0 μL,2.5 μL Mix,1.1 μL ddH2O以及0.4 μL Primer。
参考刘蕊等[16]所使用的程序,稍作修改,PCR扩增程序为94℃,预变性3 min;95℃变性15 s,退火(引物不同,退火温度不同)20 s,72℃延伸30 s,35个循环;72℃延伸5 min。
1.2.3 电泳检测
使用8%的非变性聚丙烯酰胺凝胶对PCR产物进行电泳检测。每孔点样量为1.5 μL,缓冲液为0.5×TBE,恒压180 v,电泳90 min。
1.2.4 数据统计与分析
电泳结束后,采用Gelred核酸染料进行凝胶染色[17],观察并统计条带。利用50 bp DNA Marker 作为分子量参考,每个SSR 位点,不同授粉系的单株在相同迁移率上有带记为 1,无带记为 0,建立 SSR 分子标记的 1/0 矩阵数据库。利用MEGA7.0进行数据处理,采用UPGMA方式进行聚类分析,得到4份多胚授粉系的遗传关系树状图。
2 结果与分析
2.1 SSR引物多态性
本研究通过15对SSR引物对4份甜菜多胚授粉系进行扩增,15对SSR引物共扩增出63条带,且均为多态性条带,其中TC55扩增条带最多为6条,LNX47为2条,其余引物扩增的条带总数为2~3条,实验中用到的引物名称及其序列见表2。如图1为引物L59对50份单株材料的扩增条带图,总共扩增出3条带,多态带3条。扩增结果表明,这15对引物可以用于甜菜不同单株之间的遗传多样性分析。

表2 SSR引物信息
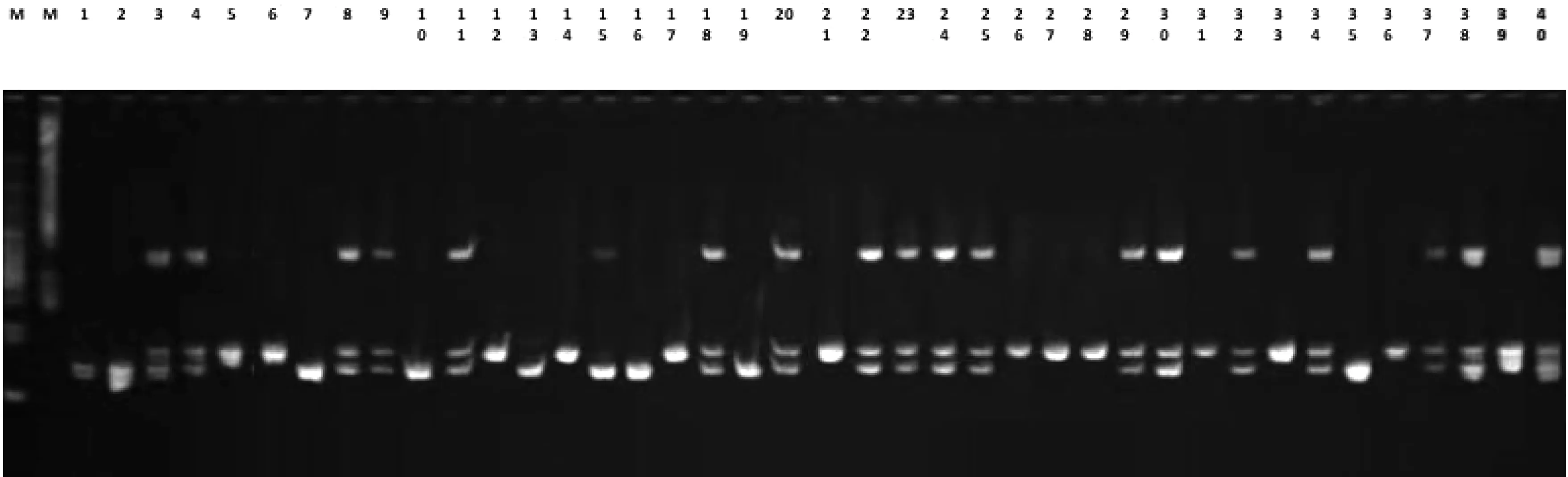
图1 引物L59对2B796的40个甜菜单株的PCR扩增图谱
2.2 遗传距离及聚类分析
利用MEGA 7.0计算同一授粉系不同个体之间的遗传距离。结果显示授粉系2B051的不同单株间的最大遗传距离为0.563,最小为0.141,平均遗传距离0.350,授粉系2B051的聚类分析图见图2(另外三个授粉系的聚类图略),根据遗传相似矩阵,在遗传距离为0.195处,可将50个单株分为2个大类群,类群Ⅰ包括6个单株,类群Ⅱ包括44个单株。类群Ⅰ、Ⅱ又可以分为多个亚群,类群Ⅰ在遗传距离为0.185时又可以将类群Ⅰ分为2个亚群(Ⅰ-1、Ⅰ-2),亚群Ⅰ-1共有43个单株,亚群Ⅰ-2共有1个单株。类群Ⅱ在遗传距离为0.150时可以分为2个亚群(Ⅱ-1、Ⅱ-2),亚群Ⅱ-1共有1个单株,亚群Ⅱ-2共有5个单株。
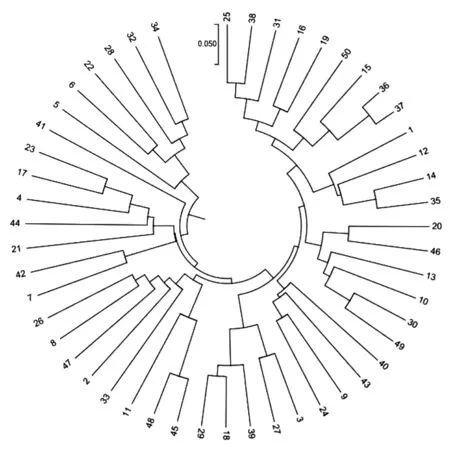
图2 2B051的50个单株聚类分析图
授粉系2B796的不同单株间最大的遗传距离为0.538,最小遗传距离为0.115,平均遗传距离为0.356。根据遗传距离越大,其两者的亲缘关系越疏远的理论基础分析,以遗传距离为0.175标准,将50个单株分为8个类群,第1个类群共有2个单株,第2类群共有4个单株,第3个类群共有3个单株,第4个类群共有2个单株,第5个类群,共有10个单株,第6个类群共有8个单株,第7个类群共有10个单株,第8个类群,共有11个类群。
授粉系2B802的遗传距离在0.132~0.528 之间,平均遗传距离为0.342。当遗传距离为0.200时,授粉系2B802可以分为2个大类群,当遗传距离为0.1750时,可以分为5个类群分别为类群一包括单株3共一个单株,类群二包括单株39、单株34、单株40共3个单株,类群三包括单株36、单株27共单株,类群四包括11个单株8、12、16、4、18、24、22、23、17、41、43,类群五包括单株10、14、46、45、13、32、21、48共8个单株。1 、44、30、15、19、2、36、20、33、9、49、37、42、5、29、7、6、28、31、38、25、26、50、11、47,共25个单株。
授粉系2B807的遗传距离在0.140~0.509 之间,其平均遗传距离是0.352。当遗传距离为0.200时,授粉系2B807的50个单株可以分为2大类群,类群Ⅰ包括49个单株,类群Ⅱ只有1个单株。当遗传距离在0.195时,类群Ⅰ可以分为亚群Ⅰ-1、亚群Ⅰ-2。亚群Ⅰ-1包括48个单株,亚群Ⅰ-2包括1个单株。
综合4个授粉系的信息表明,同一授粉系不同个体之间存在较大的差异性,同一授粉系内部遗传差异较大。
3 讨论与结论
种质资源的遗传多样性主要是指种群内个体间遗传的差异,是生物遗传进化和再生产的基础,是遗传多样性的核心和重要组成部分,对甜菜品种改良和新品种选育有直接影响的是种质遗传多样性的丰富程度[18]。李满红等指出甜菜新品种种植和生产的瓶颈问题是缺乏优良的种质资源,特别是缺乏高产、高糖和多种适合抗病机械化作业的资源材料[19]。
本实验通过15对引物分别对4份甜菜授粉系50个单株进行PCR扩增及引物多态性分析,结果表明四份甜菜授粉系内部具有极其丰富的多态性。通过遗传距离及聚类分析结果表明,甜菜2B051的遗传距离在0.141~0.563之间,甜菜2B796的遗传距离在0.115~0.538之间,甜菜2B802的遗传距离在0.132~0.528 之间,甜菜2B807的遗传距离在0.140~0.509 之间,其平均遗传距离分别为0.350、0.356、0.342、0.352。表明这四种甜菜多胚授粉系不同单株之间的遗传距离大,遗传多样性高,亲缘关系较远。聚类分别将四份授粉系的50个单株分为多个类群,由此进一步说明同一授粉系的不同单株之间的遗传多样性高。
近年来发现国外甜菜品种的一致性非常好,但是国产甜菜品种的一致性较差,原因可能是由于国外甜菜品种的父母本比较纯,母本为较纯的自交系,而父本也可能具有自交可育基因,经过多代自交而成。而我国多胚种质资源中缺乏自交可育基因,因此大都通过母系选择或者混合轮回选择的方法创造多胚授粉系,通过本次实验发现这种方法创造的授粉系内部遗传差异较大,纯度低,这也造成了我国自育的甜菜品种一致性较差。
在育种工作中,可以有目的地选择和使用甜菜材料。决定甜菜育种成败的关键是亲本选配,在选择亲本材料作为杂交育种亲本时,除考虑材料间的亲缘关系外,还要考虑其复杂的遗传组分,尽可能挑选亲缘关系相对较远,遗传背景不同的材料作为父母本,这样才能提高杂种优势,有助于选育出更好的后代杂交种[20]。在今后的的育种中,我们要更多的引入具有自交可育基因的多胚种质资源,创造甜菜多胚自交系,对指导甜菜杂交育种工作与未来的甜菜育种研究具有重要指导意义。
致谢
本研究由财政部和农业农村部国家现代农业产业技术体系(CARS-170111)资助。
