2 种带鱼科鱼类线粒体COI 基因片段的比较分析
2022-05-18斯舒谨褚梦洁王忠明杨天燕朱文斌
斯舒谨,褚梦洁,袁 帆,王忠明,杨天燕,朱文斌
(1.浙江海洋大学水产学院,浙江舟山 316022;2.浙江海洋大学海洋与渔业研究所,浙江省海洋水产研究所,浙江省海洋渔业资源可持续利用技术研究重点实验室,农业农村部重点渔场渔业资源科学观测实验站,浙江舟山 316021)
带鱼Trichiurus lepturus 和沙带鱼Lepturacanthus savala 分别为隶属于鲈形目Perciformes,带鱼科Trichiuridae,带鱼属Trichiurus 和沙带鱼属Lepturacanthus 是海洋中上层凶猛肉食性鱼类[1-2]。带鱼在我国沿海均有分布,和大黄鱼、小黄鱼及乌贼并称为中国的“四大海产”,也是东海渔场最重要的经济鱼类之一,历史上渔获量曾占全国总产量的80%以上[3-4]。沙带鱼产量相较带鱼少,分布于日本至澳大利亚、马来半岛及印度等近海水域,国内则常见于东海和南海[1-2],因背鳍金黄,福建沿海又有“黄鳍带鱼”或“黄金带鱼”的俗称。20 世纪70 年代中期,随着海洋捕捞强度的日益增加及环境的破坏,东海渔业资源面临巨大压力[5]。以带鱼为例,2008-2015 年间东海带鱼资源量发生剧烈波动,呈现持续下降趋势。具体表现在群体组成低龄化和小型化、性成熟提前和初次性成熟最小肛长趋小以及单位渔获量下降等方面[6-7]。近年来,随着拖网和帆张网等主要带鱼捕捞作业方式减船转产、伏季休渔延长、产卵场保护区设立、执法力度加大,带鱼资源状况有好转的趋势。
线粒体DNA(mitochondrial DNA,mtDNA)严格遵守母系遗传、具有分子结构简单稳定、可溯源性强等特点,随着DNA 测序技术和分子信息学的快速发展,mtDNA 在鱼类系统学、种间杂交渐渗、群体遗传学等方面得到广泛应用,成为当今分子生物学研究中的一个热门领域[8-10]。细胞色素c 氧化酶亚基I(cytochrome c oxidase subunit I,COI)基因是线粒体DNA 中13 个蛋白质编码基因之一,因携带的遗传信息量大、进化速率适中、解析能力强,主要应用于群体遗传结构和系统发育分析、种类鉴定以及DNA 条形码研究等领域[11-12]。
本研究分别选取我国舟山带鱼和北海沙带鱼为研究对象,采用线粒体COI 基因片段序列作为分子标记,探讨2 种带鱼科鱼类线粒体COI 基因片段序列的差异性,以期为带鱼和沙带鱼的种质资源保护研究和开发利用提供遗传背景资料,并为带鱼科鱼类分类和系统进化提供分子依据。
1 材料与方法
1.1 实验材料
本研究实验材料为2019 年7-9 月采集自舟山近海的带鱼和广西沿海的沙带鱼,2 种鱼类样本共计52 尾(图1)。所有样品取背部新鲜肌肉组织存放于5 mL 离心管,加入无水乙醇置于-20 ℃保存备用。样本数量、采集时间和地点信息如表1 所示。

图1 2 种带鱼科鱼类的采样图(括号数字代表样本数量)Fig.1 The sampling locations of two Trichiuridae fishes (numbers in brackets represent the number of samples)
1.2 基因组DNA 提取及PCR 扩增
剪取适量背部肌肉组织,采用传统的Tris 酚-氯仿法[13]提取带鱼和沙带鱼基因组DNA,将75%乙醇沉淀后经自然风干的DNA溶解于100 μL 灭菌水中,并放于4 ℃保存备用。用于扩增COI 基因序列的正反向引物分别为COI-F:5′-TCAACCAACCACAAAGACATTGGCAC-3′和COI-R:5′-TAGACTTCTGGGTGGCCAAAGAATCA-3′。PCR 总反应体系为25 μL,各试剂剂量和引物剂量如表1 所示。

表1 PCR 反应体系Tab.1 The PCR reaction system
PCR 反应程序为:95 ℃预变性5 min;95 ℃变性0.5 min,54 ℃退火0.5 min,72 ℃延伸1.25 min 循环35 次;72 ℃延伸5 min。用浓度1.5%琼脂糖凝胶电泳检测PCR 扩增产物,凝胶成像系统拍照后,挑选条带亮度较高的PCR 产物送至上海桑尼生物有限公司进行双向测序。
1.3 数据分析
运用DNAstar Lasergene 7.1 软件包[14]中的MegAlign 和Seqman 程序对测序结果进行比对和编辑;采用DnaSP 6.0 软件[15]计算2 种鱼类的单倍型数(H)、核苷酸多态位点数(S)、单倍型多样度(Hd)和核苷酸多样性指数(Pi);DAMBE 软件[16]进行碱基替换饱和性分析;使用MEGA 6.0 软件[17]分析序列碱基组成和变异位点情况,基于Kimura 2-Parameter(K2P)模型计算群体间遗传距离并构建邻接系统树,系统树各分支的置信度采用自展法重复1 000 次检验。
2 结果
2.1 COI 基因序列特征
经比对和校正去除序列两端测序不准确部分,最终得到52 条长度为550 bp 的带鱼和沙带鱼COI基因同源序列。利用MEGA 软件计算序列碱基组成如表2 所示。从碱基组成来看,沙带鱼的T 碱基含量最高,而带鱼C 含量最高,但2 种带鱼科鱼类G 含量都最低,且A+T 含量大于C+G 含量,这与脊椎动物线粒体DNA 的特征相符[18]。碱基转换明显大于颠换,转换/颠换(R)平均值为2.72,变化范围在2.65~10.43 之间。4 种碱基在密码子第1、2、3 位点的平均含量存在一定差别,其中第1 位点各碱基含量差别较小,第2 位点T 碱基含量偏高而A、G 碱基含量大致相当,G 含量偏低这一现象在密码子第3 位点尤为突出(图2)。在183 个三联体密码子中,第1 位点发生碱基突变的有37 个(27 个碱基转换、10 个碱基颠换),第2 位点发生碱基突变的有3 个(3 个碱基转换),第3 位点发生碱基突变的有1 个(1 个碱基颠换),由此可以看出密码子第1 位点碱基突变数目明显高于其它位点。

图2 带鱼和沙带鱼COI 基因片段密码子碱基组成Fig.2 Codon composition of COI gene fragment in T.lepturus and L.savala

表2 带鱼和沙带鱼COI 基因片段碱基组成Tab.2 Base composition of COI gene fragments in T.lepturus and L.savala
2.2 遗传多样性
2 种鱼类COI 基因序列共检测到25 种单倍型,其中24 尾带鱼样品中检测到13 种单倍型(DYhap1-DYhap13),28 尾沙带鱼样品中存在12 种单倍型(SDYhap14-SDYhap25)。52 条同源序列存在变异位点总计105 个,约占序列总长度的19.1%,其中13 个带鱼COI 基因序列中有13 个变异位点、12 个沙带鱼COI 基因序列中有28 个变异位点(图3)。沙带鱼平均单倍型多样度(Hd)及核苷酸多样性指数(Pi)分别为0.7169 和0.0050,保守位点数目为522,多态位点数目为28,占位点总数的5.09%,在28 个多态位点中,包括单一变异位点(singleton variable sites)21 个,简约信息位点(parsimony informative sites)7 个;带鱼平均单倍型多样度(Hd)及核苷酸多样性指数(Pi)分别为0.884 1 和0.004 6,保守位点数目为537,多态位点数目为13,占位点总数的2.36%,在13 个多态位点中,包括单一变异位点4 个,简约信息位点9 个(表3)。
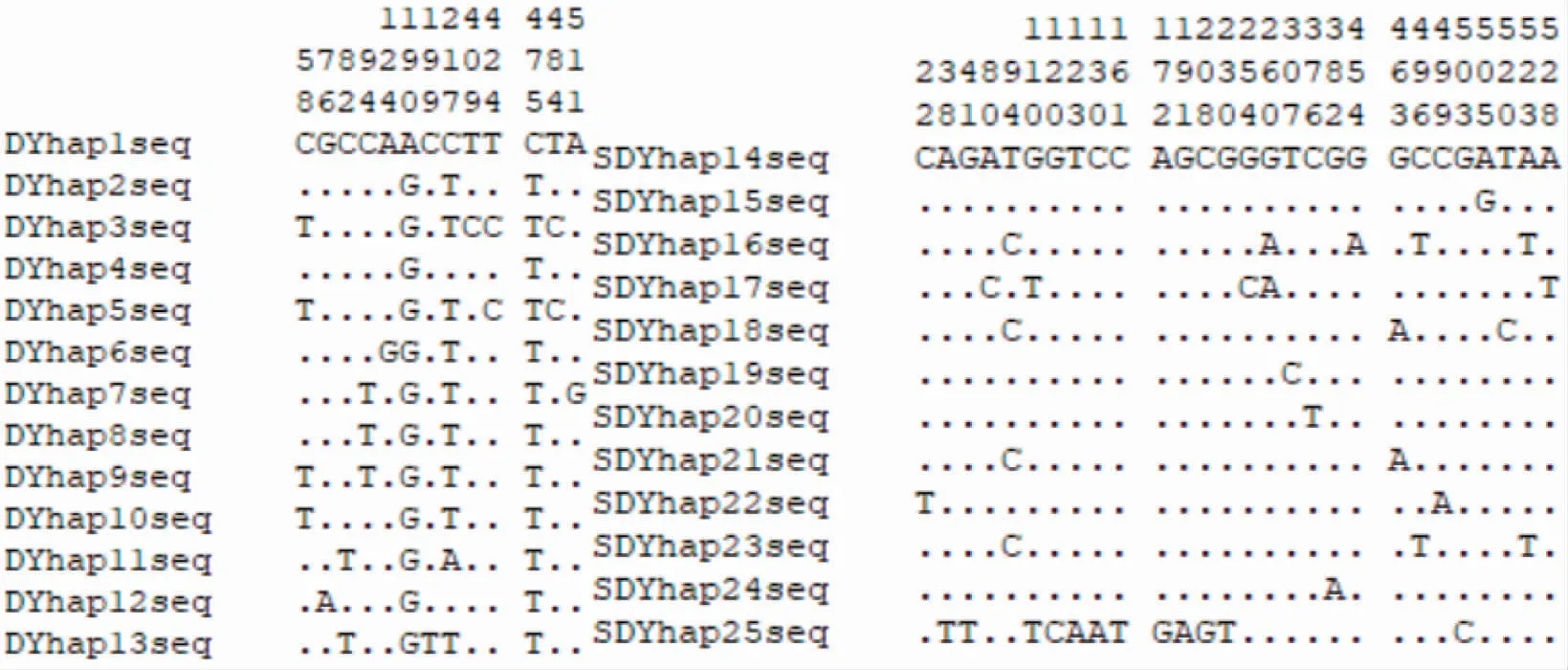
图3 COI 基因序列片段单倍型变异位点分布(左图为带鱼、右图为沙带鱼)Fig.3 Variable sites of COI gene sequence from different haplotypes (T.lepturus is on the left and L.savala is on the right)

表3 基于线粒体COI 基因的带鱼和沙带鱼遗传多样性参数Tab.3 Genetic diversity indexesbetween T.lepturus and L.savala based on mtDNA COI gene
2.3 遗传距离及系统发育
采用Kimura 2-Parameter 模型分别计算沙带鱼和带鱼种内个体间平均遗传距离分别为0.004 9 和0.004 5,前者略大于后者。基于COI 基因序列得到的沙带鱼和带鱼种间的平均遗传距离为0.165 5,种间的遗传距离远大于种内的遗传距离。
使用DAMBE 软件进行替换饱和性分析,并以碱基转换数和颠换数为纵轴,以P-distance 模型校正的遗传距离为横轴做散点图(图4),显示碱基转换和碱基颠换均呈直线上升趋势,COI 基因序列碱基替换数与遗传距离呈显著的线性关系且未出现平台期,表明用于本研究的基因序列替换未饱和,适用于构建系统发育树。图5 为基于25 个单倍型构建的邻接(neighbor-joining,NJ)关系树,可以看出2 种带鱼科鱼类分别聚类成独立的两支。从NCBI(National Center for Biotechnology Information)数据库下载19 种带鱼科鱼类COI 基因序列(表4),选择本研究中2 种鱼类单倍型数量最多的各1 条序列作为代表(DYhap2 和SDYhap14),以花鲈Lateolabrax japonicus(GenBank 登录号:MF122427)作为外群,比对截取长度为504 bp 的同源序列,采用NJ 法构建系统发育关系树如图6 所示。
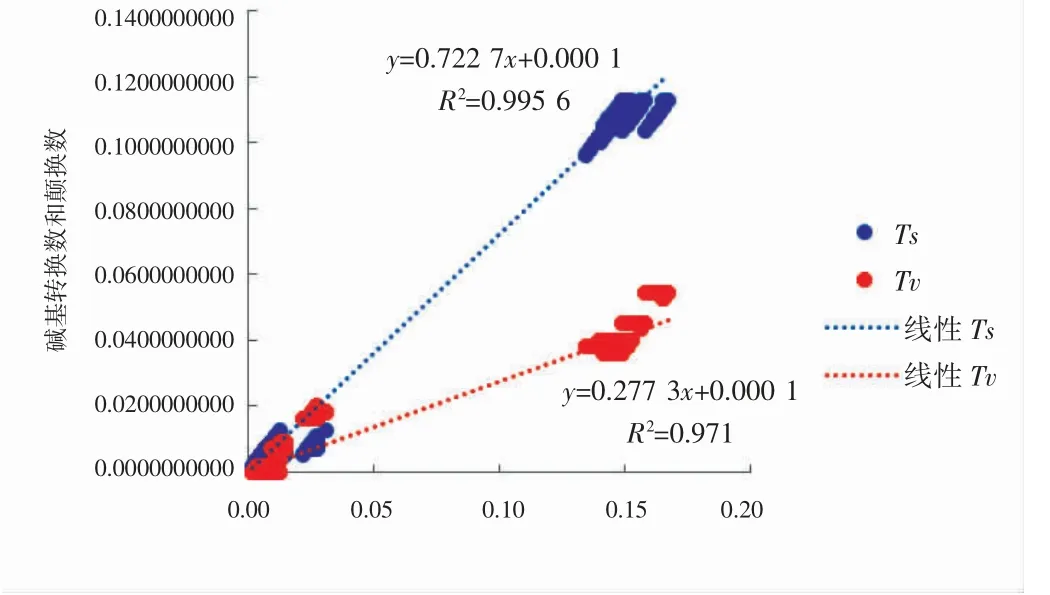
图4 线粒体COI 基因碱基替代饱和度分析Fig.4 The substitution saturation analysis of mitochondrial COI gene

表4 用于构建系统进化树的带鱼科不同物种序列信息Tab.4 Sequences of Trichiuridae fishes used for phylogenetic tree construction

图5 基于单倍型构建的带鱼和沙带鱼NJ 系统树Fig.5 NJ phylogenetic tree of T.lepturus and L.savala based on haplotypes

图6 基于邻接法构建的带鱼科鱼类系统发育关系树Fig.6 The phylogenetic tree of Trichiuridae fishes based on neighbor-joining method
3 讨论
全球带鱼科鱼类的主要捕捞海域包括西北太平洋、北印度洋和西非,其中西北太平洋是主要产区。我国近海5 种主要带鱼科经济捕捞种类有沙带鱼、小带鱼Eupleurogrammus muticus 以及带鱼属的带鱼、短带鱼T.brevis 和南海带鱼T.nanhaiensis。因前两者产量较低且缺乏数据资料,在渔业统计时常被统计为带鱼。作为中国海洋鱼类中捕捞产量最大的品种,20 世纪90 年代,我国带鱼科鱼类捕捞量就已占全球总产量的60%[4,19],近年来平均年捕捞产量基本维持在100 万t 左右[3]。
目前国内关于带鱼的研究主要集中在渔业生物学、种群动态、资源评估模型应用以及遗传多样性等方面。舟山带鱼口感鲜嫩、肉质细腻,是我国首批获得海鲜类地理标志证明的产品,舟山带鱼有别于其他带鱼的优越品质是否与种质因素有关尚未得知。目前专门针对舟山近海带鱼种群遗传学的研究不多,仅见李鹏飞等[20]曾对浙江近海带鱼的线粒体COI 基因进行比较分析,发现带鱼该基因变异程度较小;郑文娟等[21]基于线粒体控制区对舟山海域3 个带鱼地理群体遗传多样性分析,发现群体间分化程度小、基因交流频繁,遗传多样性处于中等偏下水平。本研究对采自舟山近海的24 尾带鱼线粒体COI 基因片段序列分析发现其平均单倍型多样度(Hd=0.884 1)及核苷酸多样性指数(Pi=0.004 6)均低于卞光明等[22]的研究结果(Hd=0.998 3,Pi=0.006 3)。该结果显示近年来舟山近海带鱼遗传多样性水平呈现某种程度的下降趋势。造成这一差异的原因一方面与采样地点和取样数量有关。另一方面,由于遗传多样性是生物多样性的基础,也是物种和种群长期续存的进化潜力,遗传多样性降低会引起生物对环境适应力的减弱并最终导致灭绝[23-24]。尽管我国实行的伏期休渔期制度对保护海洋资源起到了积极作用,但日益增长的市场需求对带鱼野生资源造成的捕捞压力以及带鱼这种深海鱼类难以突破人工繁殖的世界性难题,使得舟山渔场带鱼资源量恢复缓慢,种群数量的减少加之栖息地环境的改变使得舟山带鱼种质资源恢复面临一定压力。从2 种带鱼科鱼类的遗传差异来看,沙带鱼多态位点数目、平均核苷酸差异数和核苷酸多样性均高于带鱼,表明较之带鱼而言,沙带鱼具有较丰富的遗传资源和较好的开发潜力。
线粒体COI 基因作为DNA 条形码(DNA barcoding),已广泛应用于昆虫、鸟类、甲壳类及鱼类等物种的快速识别和鉴定,成为当今生态学研究的重要工具[25]。HEBERT,et al[26]分析比较了13 320 种生物的线粒体COI 基因序列,发现绝大多数生物的种内遗传距离差异在0%~2%之间,大于2%非常少,种间遗传距离则高达11.3%[26]。本研究发现沙带鱼和带鱼种间平均遗传距离为0.165 5,约为种内平均遗传距离(0.004 9和0.004 5)的33~36 倍,表明各物种的COI 序列间已经形成明显“条形码间隙”(Barcoding gap),即种间遗传距离为种内遗传距离的10 倍及以上[27]。基于K2P 遗传距离构建的NJ 系统树明显分为2 支,带鱼和沙带鱼单倍型各自聚在一起,也表明两种鱼类可以得到很好地区分。结合NCBI 数据库中下载的其他19 种带鱼科鱼类COI 基因序列进行聚类分析,发现这些鱼类按照尾鳍形态可分为鞭状和叉状2 大类群,前者包含了绝大多数常见的种类,其中带鱼属和沙带鱼属亲缘关系最近,两者聚类后再与小带鱼属鱼类相聚,小带鱼属是最早分化出来的种类,其次为沙带鱼属,而带鱼属则是最新演化的种类;后者主要包含了叉尾带鱼属和等鳍叉尾带鱼属鱼类,与蔡传庚对10 种带鱼科鱼类COI 基因聚类的结果相似[28]。有学者曾将带鱼科划分为3 个亚科,其中等鳍叉尾带鱼亚科Aphanopodinae 包含了等鳍叉尾带鱼属Aphanopus 和深海带鱼属Benthodesmus[29]。但从本研究的聚类情况来看,两者并未聚类到一起,彼此间的演化关系也不甚明确,还有待于今后增加样品种类和数量并结合其他分子标记进行深入探讨。
