2020~2021年广州市售牡蛎中GII型诺如病毒污染情况调查
2022-05-14杨家乐蔡伟程李贻静高珺珊陈谋通叶青华张菊梅别小妹吴清平
杨家乐,薛 亮 ,蔡伟程,李贻静,高珺珊,陈谋通,叶青华,吴 诗,张菊梅,别小妹 ,吴清平
(1.广东省科学院微生物研究所,广东省科学院,华南应用微生物国家重点实验室,广东省微生物安全与健康重点实验室,广东广州 510070;2.南京农业大学食品科技学院,江苏南京 210095)
诺如病毒(Norovirus,NoV)是一种常见的食源性病毒,人感染后会出现腹泻、呕吐等急性肠胃炎症状,通常在医院、学校、养老院等人员密集场所易发生小规模的聚集性暴发[1−2],不但对人体健康产生危害,同时也造成了巨大的经济损失。根据衣壳蛋白(VP1)的序列,诺如病毒可以分为GI-GX十种基因组,包括48个基因型[3−4]。诺如病毒传播主要通过粪口途径,患者的呕吐物及粪便中一般都有较高浓度的病毒,有较强的传染性,并通过气溶胶传播[5]。近些年国内各个省市因感染诺如病毒而出现急性肠胃炎症状的患者数量居高不下[6−9],其中2013~2017年广东省报告的散发诺如病毒感染167例,超过60%的病例是由GII.2、GII.4、GII.17型病毒引起[10]。
诺如病毒能够在自然环境中持续存在并保持感染性,主要通过将未经处理或未经充分处理的污水排放或泄漏到泉水、河流、水坝或入海口等水源中而最终进入环境中。该病毒常在食品及水中被检出[11−13],新鲜的水果、蔬菜表面因接触污水也存在污染风险,双壳贝类(如牡蛎、贻贝等)一般在近海附近养殖,作为滤食性生物能够过滤大量的水,对水中的病原微生物起到富集的作用[14];尤其贝类的消化组织中存在类组织血型抗原[15],在病毒与贝类组织结合过程中发挥重要作用,因此消化组织是病毒富集的主要部位,富集含量最高[16]。由于贝类中的病毒主要来自于环境,因此当新的病毒流行株在环境中出现时,贝类中也会检测到同样的病毒基因型。由于GII型诺如病毒结合的组织类型比其他基因型丰富,它可以存在于消化腺、肠道、腮部、外套膜等多个部位[17],因此需要重点关注该病毒基因型在牡蛎中的存在情况。
广东作为贝类养殖大省,但同时市售牡蛎中存在诺如病毒污染问题[18],食用前如不高温加热将其中的病毒灭活,有被感染的风险,监测相关贝类产品中诺如病毒的污染水平对于预防此类事件的发生有重要意义。因此,为了进一步掌握广州地区市售牡蛎中GII型诺如病毒污染情况,本研究开展了为期一年的监测调查,以期明确该地区牡蛎中诺如病毒的污染水平及季节分布规律。
1 材料与方法
1.1 材料与仪器
牡蛎样品于2020年6月至2021年5月,在广州市黄沙水产市场购买牡蛎,挑选体积大小近似(挑选的样品体长在10~14 cm之间,体宽在6~9 cm之间,牡蛎肉重量(去壳)在14~20 g之间),新鲜且外观无明显损伤的牡蛎,每个牡蛎经过无菌解剖后取消化腺组织进行下一步检测或保存在−80 ℃冰箱待检;诺如病毒阳性样本 来自广州市急性肠胃炎患者的粪便样本[19],经过处理后保存在−80 ℃冰箱;鼠诺如病毒(Murine norovirus,MNV) 由广东省科学院微生物研究所食源性病毒组提供;蛋白酶K 30 U/mg,Merck公司;聚乙二醇(PEG)6000 kingbio公司;10×磷酸盐缓冲盐溶液 PH7.2,上海生工生物工程股份有限公司;Evo M-MLV One Step RT-qPCR Kit(Probe)艾科瑞公司;Premix Taq™ (Ex Taq™ Version 2.0 plus dye)、One Step Prime Script™ RT-PCR Kit(Perfect Real Time )上海百赛生物技术有限公司;High Pure Viral RNA Mini Kit试剂盒 广州美基生物科技有限公司;引物及探针 由北京六合华大基因科技有限公司合成;100 bp DNA Ladder 北京擎科生物科技有限公司。
Microfuge 20R台式冷冻离心机 美国Beckman公司;Quant Studio 6 and 7 Flex Real-Time 荧光定量PCR仪 美国Thermo Fisher Scientific公司;罗氏LightCycler96实时荧光定量PCR仪 美国Roche公司;Biometra Tone 96 PCR仪 德国耶拿公司;振荡培养箱 上海知楚仪器有限公司;DK-S22型电热恒温水浴锅 广东环凯微生物科技有限公司;MY-10/20手持式电动组织研磨器 上海净信有限公司;SW-CJ-IFD型超净工作台 中国苏净安泰公司;T500电子天平 美国G&G公司。
1.2 实验方法
1.2.1 样品前处理 根据本实验室已建立方法进行牡蛎样本的处理[20],具体如下:每个牡蛎样品取其消化腺组织0.15 g,分装在1.5 mL离心管中打碎匀浆,加入10 μL鼠诺如病毒(作为过程控制对照),并加入1.2 mL蛋白酶K溶液(100 μg/mL)涡旋混匀,置于37 ℃恒温培养箱320 r/min振荡60 min;60 ℃水浴加热15 min,使蛋白酶K灭活,待冷却至室温,10000 r/min离心3 min;吸取上清液,加入600 μL PEG6000(36% PEG6000,0.9 mol/L NaCl)并将混合液放入4 ℃冰箱静置1 h,然后10000 r/min离心10 min;弃去上清液取沉淀;用140 μL 磷酸盐缓冲盐溶液(稀释为1×)重悬沉淀,用于核酸抽提。
此外,取鼠诺如病毒以及诺如病毒样本与样品一起提取RNA,用于计算病毒回收率以及PCR扩增效率。
1.2.2 提取病毒RNA 用High Pure Viral RNA Mini Kit试剂盒提取病毒RNA,按照说明书中的操作步骤提取病毒RNA,直接用于核酸检测,或置于−80 ℃冰箱保存代用。
提取病毒RNA后采用荧光定量RT-PCR和巢式RT-PCR进行检测,两种技术的检测灵敏度存在差异。其中荧光定量RT-PCR是目前国家标准和ISO标准推荐的方法,灵敏度较高,而巢式RT-PCR灵敏度相对较低,但其结合桑格测序能够获得目的病毒序列。
1.2.3 实时荧光定量逆转录聚合酶链反应检测 如表1和表2,反应体系:2 × One Step RT-qPCR Buffer 10 μL,Pro Taq HS DNA Polymerase 0.4 μL,Evo MMLV RTase Enzyme Mix 0.4 μL,上下游引物各0.4 μL(10 μmol/L),探针0.8 μL(10 μmol/L),RNA模板2 μL,加无RNA酶水至反应体系为20 μL。具体反应条件:42 ℃逆转录300 s,1个循环;95 ℃预变性10 s,1个循环;95 ℃变性5 s,60 ℃退火,20 s,45个循环。将诺如病毒标准品进行梯度稀释(浓度为2×108~2×100copies),根据拷贝数与PCR反应的Ct值做标准曲线。结果判断:若样品有S型扩增且Ct值≤40,判为阳性,有S型扩增且Ct值>40或无明显S型扩增曲线但报告有Ct值,仍判定为阴性。检测结果为阳性样本的Ct值,依据标准曲线(y=−3.428lg(x)+43.15)计算病毒拷贝数,其中y为荧光定量Ct值,x为病毒拷贝数。

表1 实时荧光定量逆转录聚合酶链反应(Real-time RT-PCR)的引物与探针Table 1 Primers and probes of Real-time reverse transcription polymerase chain reaction
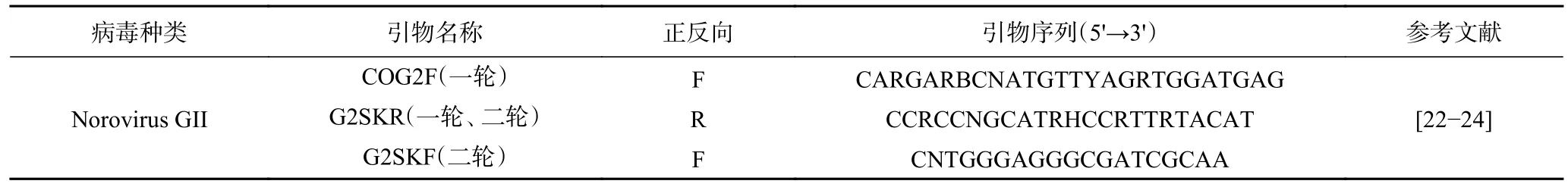
表2 巢式RT-PCR引物Table 2 Nested RT-PCR primers
1.2.4 巢式RT-PCR定性检测 荧光定量检测为阳性结果的样品,经过巢式RT-PCR扩增后进行琼脂糖凝胶电泳,符合目标条带的扩增产物送去测序。第一轮巢式PCR反应体系:2×1 Step Buffer (Dye Plus) 10 μL,PrimeScript 1 Step Enzyme Mix 1 μL,上下游引物各1 μL,RNA模板2 μL,加无RNA酶水至反应体系为 20 μL。第一轮巢式PCR反应条件:50 ℃反转录30 min;94 ℃预变性4 min,55 ℃退火30 s,72 ℃延伸30 s,35个循环;72 ℃延伸7 min。第二轮巢式PCR反应体系:Premix Taq™ (Ex Taq™ Version 2.0 plus dye) 10 μL,上下游引物各1 μL,RNA模板2 μL,加无RNA酶水至反应体系为20 μL。第二轮巢式PCR反应条件:94 ℃预变性3 min;94 ℃变性30 s,50 ℃退火30 s,77 ℃延伸30 s,30个循环;72 ℃延伸5 min。
1.2.5 基因序列分析 扩增产物经琼脂糖凝胶电泳分析后,将阳性扩增产物送于北京六合华大基因科技有限公司直接测序进行测序,将测序结果提交至在线基因分型工具(https://www.rivm.nl/mpf/typingtool/norovirus/),并在NCBI数据库进行BLAST在线比对(https://blast.ncbi.nlm.nih.gov/Blast.cgi)。为进一步分析测序毒株,选取诺如病毒的各基因型毒株作为代表毒株进行比对,参考株基因序列均来源于Gen-Bank。采用ClustalX软件进行序列比对,MEGA 7.0.26软件构建进化树(Neighbor Joining 法),bootstrap 检验设置1000次。
1.3 数据处理
本研究对每份样品的回收率和PCR扩增效率均进行3次平行实验以确保数据的稳定性,若样品回收率>1%则判定为有效提取,否则不纳入数据分析;采用EXCEL 2019汇总全部的原始资料和检测结果;应用SPSS20.0软件对诺如病毒检出情况进行统计学分析,每月的平均病毒拷贝数用(平均值±标准差)表示,采用单因素方差分析对不同季节的病毒检出率进行比较,检验水准α=0.05。
2 结果与分析
2.1 市售牡蛎中诺如病毒检出情况
检测结果显示(表3和表4),广州市市售牡蛎中存在诺如病毒污染情况,在2020年6月~2021年5月共计110份的样品中,实时荧光定量RT-PCR检测结果阳性样本数为58份,全部为诺如病毒GII型,总体阳性率为52.7%。病毒污染含量范围为1.56×103~1.09×106copies/g(消化腺)。
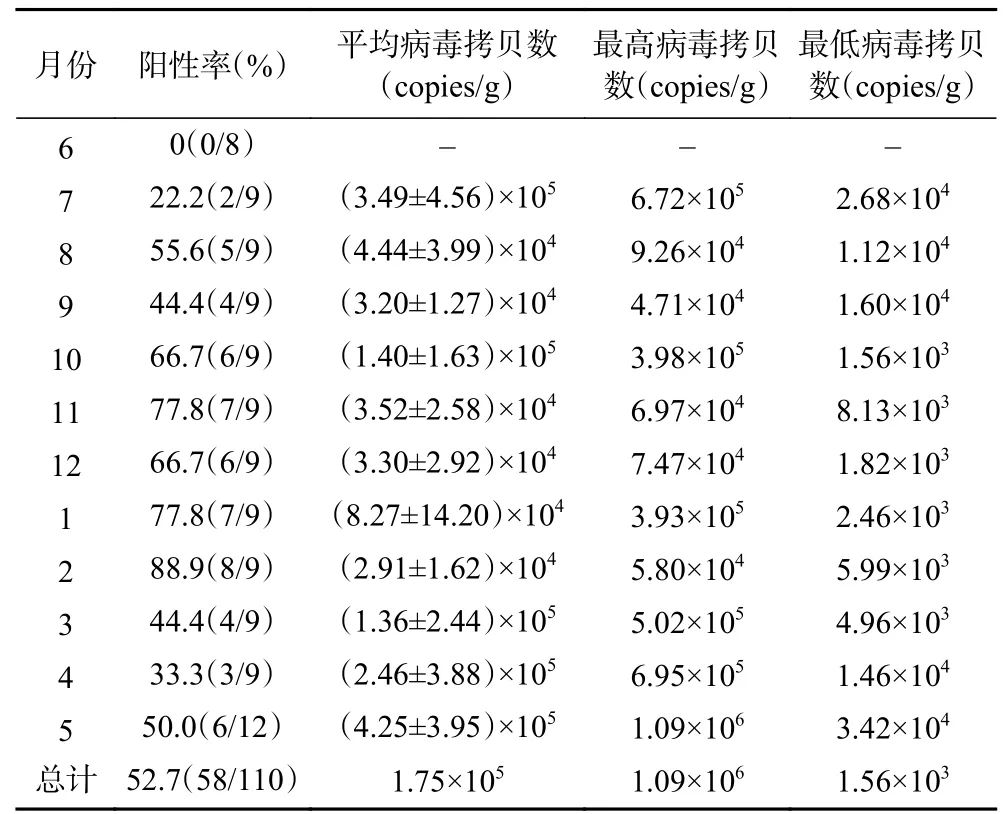
表3 2020年6月~2021年5月广州市售牡蛎诺如病毒(GII)检测情况Table 3 Detection results of norovirus GII in commercial oysters in Guangzhou from June 2020 to May 2021
2.2 市售牡蛎中诺如病毒检出的季节分布规律
如表3所示,在2020年6月~2021年5月期间,病毒检出阳性率随季节变化而变化,冬季阳性率高于其他季节。如表3所示,在2020年6月~2021年5月期间,春季(3~5月)采集牡蛎样品30个,其中阳性样本数为13;夏季(6~8月)采集牡蛎样品26个,其中阳性样本数为7;秋季(9~11月)采集牡蛎样品27个,其中阳性样本数为17;冬季(12~2月)采集牡蛎样品27个,其中阳性样本数为21;秋季及冬季时检测到诺如病毒阳性牡蛎样本数要高于春季及夏季,所有的阳性样本中,病毒含量均高于1000 copies/g(消化腺)(图1)。
从季节分布来看,春季(3~5月)采集的样本中诺如病毒平均污染量为2.69×105copies/g(消化腺),夏季(6~8月)采集的样本中诺如病毒平均污染量为1.97×105copies/g(消化腺),秋季(9~11月)采集的样本中诺如病毒平均污染量为6.91×104copies/g(消化腺),冬季(12~2月)采集的样本中诺如病毒平均污染量为4.83×104copies/g(消化腺),不同季节中检出的阳性样本中病毒含量无明显差异。从不同数量级病毒中的阳性样品数量分布(表5)来看,病毒含量在104~105copies/g之间的样品最多。

表5 不同数量级病毒中的阳性样品数量分布Table 5 Distribution of positive samples in norovirus of different orders of magnitude
广东省属于亚热带季风气候,秋冬季节温度较低(平均温度在20 ℃以下),降雨较少,而春夏季温度较高,降水较多,对环境中病毒含量可能会造成一定影响。本文中的结果在一定程度上反应出牡蛎污染病毒的季节性差异,在下一步持续工作中,需要提高不同季节样本检测数量,更加系统的表征季节对于污染牡蛎的影响。
2.3 市售牡蛎中诺如病毒毒株系统发育分析
在对市售牡蛎的检测中,为了确定阳性样本中的病毒基因型,常利用巢式RT-PCR方法并结合桑格测序。由于巢式RT-PCR灵敏度相对较低,因此一些阳性样品无法通过巢式RT-PCR得到病毒序列,在58份阳性样品中最终得到14份阳性样本的测序结果,将序列提交至GenBank数据库,获得序列号为OK090974-OK090987。在GenBank上进行BLAST比对,构建进化树进行基因型鉴定(见图1)。其中仅一条序列为GII.17基因型,来自12月份的牡蛎样品,它与2015年美国检测到的毒株(MT344182)、2016年的巴西检测到的毒株(MN045198)、2018年在韩国检测的毒株(MN461158)相似率均超过99%;此外13条序列均为GII.4基因型,分别来自2、3、5月份的牡蛎样本,它们与2012年广东省的毒株(MH 469205)、2013年美国检测到的毒株(KY486271)、2017年澳大利亚检测的毒株(MK280937)相似率超过90%,与本团队前期研究中检测到人群中流行的病毒基因型基本一致[9,19]。

图1 广州市售牡蛎中诺如病毒序列的系统发育树Fig.1 Phylogenetic trees for norovirus sequences detected in commercial oysters in Guangzhou
3 讨论
近些年世界范围内与食用贝类相关的诺如病毒胃肠炎疫情层出不穷[25−28],多数是因为生食或者食用轻度加热的贝类所导致。大多数国家或地区规定了贝类组织或贝类养殖水中肠道致病菌的可接受水平,例如在欧盟标准中,对贝类生产区划分了A、B、C三个等级,其中规定A区中80%以上的产品中大肠杆菌含量必须少于230 个/100 g,B区中90%以上的产品中大肠杆菌含量少于4600 个/100 g,C区中全部产品中大肠杆菌含量少于46000 个/100 g[29],这些标准很好的控制细菌作为贝类相关疾病暴发原因,但未能阻止许多病毒源性疾病的暴发[30],因食用贝类而引起的病毒性胃肠炎暴发的调查表明,在患者的体内或使用过的餐具中检出了诺如病毒[31−32]。虽然在牡蛎中存在的诺如病毒基因型较丰富,但本研究中主要检测牡蛎中GII型的诺如病毒,因为大部分患者是以感染GII型诺如病毒为主[33],广州地区市场中的牡蛎普遍存在污染问题,我们通过市售牡蛎监测调查发现该地区牡蛎中GII型诺如病毒的阳性率达到了52.7%,病毒含量最高达到了1.09×106copies/g,最低为1.56×103copies/g,经测序确定病毒基因型包括GII.4及GII.17型。由于国内消费者更习惯高温烹饪而不是直接生食牡蛎,即使病毒检出率偏高但也不会引起较高的人群感染率,但不能排除在高温加工后仍有小部分病毒未被灭活,对于免疫力低下的人群如老人及儿童,需要注意此类风险。
沿海的国家和地区因地理和饮食特点等原因经常发生贝类相关的诺如病毒感染,例如韩国[34],日本[35],中国山东[36],广东[37]等地都有报道。因此对贝类中诺如病毒的持续监测具有重要意义,很多国家一直在持续检测国内相关产品中的诺如病毒污染状况。意大利的研究人员在2005~2008年间采集的116个双壳贝类样本中检出阳性率10.3%,其中GII.4-2004和GII.b为主要基因型[38]。在意大利的另一个监测调查中,贝类样本中诺如病毒的检出率为13.2%(31/235)[39]。荷兰为期两年的监测中,在牡蛎和贻贝样本中诺如病毒的检出率为16.7%(7/42份),包括GII.4等[40]。2017年7月到2018年8月在对中国山东省两个牡蛎养殖区为期一年的采样检测中,诺如病毒GII基因型阳性率为31.7%(19/60)[41]。越来越多的国家意识到监测牡蛎中食源性病毒的重要性,不仅可以控制病毒引发的疾病风险,为贝类产品中病毒净化方法提供参考标准,还为预测可能的病毒流行株提供了相关数据。
本研究的样品主要来自广东省阳江、茂名、台山、湛江、顺德等地,在采样时选择体积大小相近,新鲜有活力的牡蛎样品,对牡蛎进行无菌解剖取其消化腺组织。根据文献报道,在大多数的病毒污染牡蛎的调查中,检测的组织是消化腺,因为该组织中病毒含量较其他部位高,相对容易检出[16]。本研究中市售牡蛎诺如病毒总检出率为52.7%(58/110),结果表明广州市售牡蛎中诺如病毒污染水平较高,检出率高于北京市(37.5%,21/56)[42]和连云港市(26.67%,48/180)[43],这可能和样本前处理方法或结果判断条件有关,本研究使用实验室前期建立的检测方法,病毒回收率是ISO/TS 15216-2:2013方法的7倍[20],对病毒含量偏低的牡蛎样本有较好的检测效果。同时,在对同一地区中牡蛎污染率增加的调查表明,阳性率的增加(2.7%上升至20.7%)可能与牡蛎产区的污染增加有关[44]。
秋冬季检测的牡蛎中病毒的阳性率普遍比春夏季要高,而病毒含量水平却没有较大差异,这一情况与其他研究中的结果相似[45]。由于冬季环境温度有利于诺如病毒持续在环境中存在[46],且冬季是诺如病毒流行的高峰时间段,因此在一定程度上会造成环境中如污水中病毒含量升高,滤食性双壳贝类如贻贝、蛤和牡蛎在近海口地区生长时,会受到污水排放影响而富集水中的诺如病毒[47],构成了人-环-食传播路径。
测序结果显示检测的样本中大多数为GII.4型毒株,GII.4型毒株是全球范围内感染人体和在环境中检测到的主要毒株,在70.0%~80.0%的急性胃肠炎患者的排泄物中都有检测出该基因型的诺如病毒,并且GII.4基因型的变异株也陆续出现[48−49],另有一株GII.17型,近几年GⅡ.17型逐渐成为世界范围内的优势流行株,Pu等[50]发现GII.17型毒株在污水、牡蛎和肠胃炎病例中的同时存在和占主导地位。在对北京市售牡蛎中诺如病毒检测结果中同样检出该基因型,与人感染的GⅡ.17 毒株相似性高达 99.2%~100.0%[51]。
诺如病毒感染人体所需的剂量极低(18~2800个拷贝数)[52],根据本文的研究结果,广州市售牡蛎中诺如病毒含量达到1.56×103~1.09×106copies/g(消化腺),因此食用时加工不彻底或者直接生食牡蛎,存在较大的诺如病毒感染风险。
4 结论
贝类是全球重要的经济水产品,同时也是食源性诺如病毒传播的重要载体。通过对市售牡蛎中诺如病毒污染状况的监测研究,发现GII型诺如病毒阳性检出率达52.7%,且GII.4型为优势基因型,与病毒的临床流行存在紧密联系。因此,直接食用未加工或加工不彻底的市售贝类仍存在较大食品安全风险。本研究建议持续加强贝类中食源性病毒污染的长期监测工作,将有助于进一步推动我国贝类产业的良性发展,保障民众健康。
