利用CRISPR/Cas9技术敲除GS3和GS9基因改良水稻粒型性状
2022-05-12韩政宏段宇轩徐善斌王敬国刘化龙杨洛淼郑洪亮邹德堂
韩政宏,段宇轩,徐善斌,王敬国,刘化龙,杨洛淼,贾 琰,辛 威,郑洪亮,邹德堂
(东北农业大学 寒地粮食作物种质创新与生理生态教育部重点实验室,黑龙江 哈尔滨 150030)
水稻(Oryzasativa)是世界范围内最主要的粮食作物之一,为全球1/2以上的人口提供主要的食物来源[1]。近年来,人们对优质稻米的需求量逐渐增加,优质稻米除了有良好的食味品质外,还应该具有良好的外观品质。水稻粒型是稻米外观品质的重要构成因素之一,对稻米的外观品质有直接影响。因此,通过改良粒型提高水稻外观品质,是近年来水稻育种的一个重要目标。迄今为止,水稻中已有多个粒型相关基因被发现并克隆,如GSA1[2]、GW2[3]、GS2/GL2[4]、TGW2[5]、GS3[6]、OsLG3[7]、GL3.1/qGL3[8]、qLGY3/OsLG3b[9]、TGW3/GL3.3[10]、GL4[11]、qSW5/GW5[12]、GS5[13]、TGW6[14]、GW6a[15]、GL6[16]、GLW7[17]、GL7/GW7[18]、GW8[19]和GS9[20]等。其中,GS3、GL3.1/qGL3、TGW3/GL3.3和TGW6是水稻粒长性状的主效基因,并对该性状起负调控作用;而OsLG3、qLGY3/OsLG3b和GL4对水稻粒长性状起正调控作用。GW2、TGW2和qSW5/GW5是对水稻粒宽性状起负调控作用的主效基因,而GS5、GW6a和GW8对水稻粒宽性状起正调控作用。GSA1、GS2、GL6和GLW7对水稻粒长和粒宽性状都有正向调控作用。GL7/GW7正向调控粒长,但对粒宽起负调控作用;而GS9负调控粒长,但正向调控粒宽。对于一些负调控水稻粒型性状的QTL,如GS3、GL3.1/qGL3、GW2、TGW2等,可通过弱等位基因或功能缺失等位基因替换的方法,快速、精确改良水稻粒型相关性状[21]。这种方法可以靠2种生物技术来实现,一种是通过分子标记辅助育种从供体材料中引入弱等位基因或功能缺失等位基因,另一种是通过CRISPR/Cas9等基因编辑技术对负调控基因进行敲除。在水稻中,通过分子标记辅助育种构建近等基因系和准确引入多个等位基因需要多年时间。此外,打破基因之间的连锁也是一大难题[22],难以广泛应用。然而,利用CRISPR/Cas9介导的基因编辑系统,对负调控基因进行突变或敲除,可以方便、准确、高效地实现对水稻粒型相关性状的改良,相对于传统育种还能缩短育种年限,加快优良种质资源的创制。
CRISPR/Cas9技术是继ZFNs[23]技术和TALENs[24]技术之后的新型基因编辑技术[25],CRISPR/Cas9系统作为一种快速且高效的基因编辑工具,通过sgRNA(Single guide RNA)和Cas9(CRISPR associated protein 9)核酸酶对目的基因进行定点编辑,sgRNA是一种具有颈环结构的单链RNA,可以识别PAM(Protospacer adjacent motif)序列并引导Cas9(CRISPR associated protein 9)核酸酶对其进行切割,在PAM序列上游的第3或第4个碱基处造成DNA双链断裂(DSB)[26]。随后,生物体通过同源重组(Homologous direct recombination,HDR)修复和非同源末端连接2种方式进行修复(Nonhomologous end joining,NHEJ),在DNA双链的修复过程中形成碱基的插入、缺失和替换等突变类型,从而对目的基因进行编辑和改造。
CRISPR/Cas9技术被广泛应用于各种作物的遗传育种研究中[27-33],水稻作为单子叶模式作物,可利用CRISPR/Cas9技术在基因组水平上进行定向编辑和改造。Zhang等[34]通过CRISPR/Cas9技术,在水稻Waxy基因引入功能缺失突变,破坏Waxy正常表达,使稻米支链淀粉含量增加,水稻品种由非糯性变为糯性。Zheng等[35]通过CRISPR/Cas9基因编辑技术,对水稻己糖激酶基因OsHXK1进行敲除,上调水稻光合作用相关基因的表达,且增加了水稻的光合产物和产量。Zeng等[36]利用CRISPR/Cas9技术对水稻基因OsPIN5b、GS3和OsNYB30同时进行敲除,得到了高产且高耐寒性的长粒水稻品种。
本研究以圆粒型粳稻品种东富139、龙粳31和长粒型粳稻品种东农427为研究材料,利用CRISPR/Cas9技术同时对水稻粒型基因GS3和GS9进行敲除,得到了多个无转基因成分且纯合突变的长粒型突变株系,创制了多个长粒型粳稻品种,并且分析GS3和GS9功能缺失突变对粒型及其他性状的影响,加快了长粒型粳稻品种的选育进程,旨在为粳稻品种改良提供理论基础。
1 材料和方法
1.1 试验材料
受体材料:本研究以东富139(DF139)、龙粳31(LJ31)和东农427(DN427)3个粳稻品种为试验材料。其中,东富139和龙粳31粒型为圆粒型,稻谷长宽比1.7,东农427粒型为中长粒型,长宽比2.0。
载体:试验所用载体pYL-U3-gRNA、pYL-U6a-gRNA和pYLCRISPR/Cas9Pubi-H双元载体由华南农业大学刘耀光院士惠赠。
1.2 敲除靶点设计及载体构建
通过 NCBI(https://www.ncbi.nlm.nih.gov/)获得GS3(Os03g0407400)、GS9(Os09g0448500)的基因序列,利用 CRISPR-GE 在线网站[37](http://skl.scau.edu.cn/)的“CRISPR Primer Designer”模块设计GS3和GS9基因靶序列,在GS3第1外显子设计2对靶点及接头引物,在GS9第1外显子设计1对靶点接头引物,T1-F/R、T2-F/R及T3-F/R(表1)。并利用NCBI网站上Blast程序进行筛选,未找到脱靶序列。
参照Ma等[38]的试验方法分别构建GS3敲除载体和GS3、GS9双基因敲除载体。

表1 本研究中使用的引物Tab.1 Primers used in this study
1.3 质粒扩繁及阳性克隆筛选
将构建好的载体通过热激法转入Mach1-T1感受态大肠杆菌中,涂板培养并挑取单菌落进行PCR检测,选取阳性菌落摇菌后提取质粒,随后用AscⅠ内切酶对质粒进行酶切检测,将酶切检测无误的质粒通过热激法转入到EHA105感受态农杆菌中。
1.4 T0植株的获得
通过农杆菌介导法将CRISPR/Cas9表达载体转入粳稻品种东富139、龙粳31和东农427的愈伤组织,用潮霉素筛选出阳性愈伤组织并分化成T0植株。采用CTAB法[39]提取T0植株全基因组DNA,以潮霉素基因特异性引物Hyg-F/R进行PCR扩增,检测阳性植株。并设计引物GS3-seq-F/R和GS9-seq-F/R(表1),分别扩增GS3和GS9基因靶点及其附近的序列,对所有转基因阳性植株进行PCR扩增,PCR产物送至擎科生物技术有限公司(北京)进行测序,测序结果使用兼并序列解码(DSDecode)方法[40]进行分析。
1.5 T1纯合突变植株及无T-DNA原件植株的筛选
在T1植株分蘖期,取叶片提取全基因组DNA,利用GS3-seq-F/R和GS9-seq-F/R引物进行PCR测序,筛选出纯合突变的植株,再利用Hyg-F和Hyg-R引物在纯合植株中筛选无T-DNA元件插入的植株,并将这些植株自交繁殖至T2。
1.6 T2植株农艺性状考察
种植东富139、龙粳31、东农427野生型和T2无T-DNA元件插入的纯合突变体植株,成熟后分别测量粒长、粒宽、千粒质量、结实率及穗粒数。利用SPSS 18.0软件对农艺性状数据进行分析。
2 结果与分析
2.1 水稻GS3和GS9敲除靶点设计
本研究在GS3的第1外显子设计了2对gRNA靶点接头引物,在GS9第1外显子处设计了1对gRNA靶点接头引物,具体靶点信息见图1。

图1 GS3、GS9基因结构及靶点位置Fig.1 Gene structure and target site of GS3,GS9
2.2 GS3和GS3、GS9基因表达载体的构建
利用“金门”克隆法将带有靶点的gRNA连接到Cas9的载体骨架上,构建好的载体即为pYLCRISPR/Cas9-GS3-RNA载体和pYLCRISPR/Cas9-GS3-GS9-RNA载体(图2)。
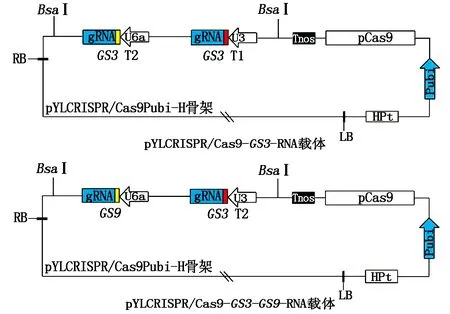
图2 pYLCRISPR/Cas9-GS3-RNA载体和pYLCRISPR/Cas9-GS3-GS9-RNA载体Fig.2 pYLCRISPR/Cas9-GS3-RNA vector and pYLCRISPR/Cas9-GS3-GS9-RNA vector
2.3 T0转基因阳性苗的获得
分别用转入pYLCRISPR/Cas9-GS3-RNA和pYLCRISPR/Cas9-GS3-GS9-RNA载体的农杆菌侵染3个粳稻品种的愈伤组织,在T0突变体植株成熟时,提取基因组DNA,用载体特异性引物Hyg-F/R进行PCR检测,筛选阳性植株。结果表明,共获得转入pYLCRISPR/Cas9-GS3-RNA载体的阳性植株48株,其中东富139、龙粳31和东农427分别获得20,10,18株;转入pYLCRISPR/Cas9-GS3-GS9-RNA载体的阳性植株100株,其中东富139、龙粳31和东农427分别获得35,32,33株。
2.4 T0突变体GS3和GS9基因靶点突变类型分析
对已筛选的阳性植株,利用引物GS3-seq-F/R和GS9-seq-F/R进行PCR扩增并测序分析,筛选得到的148株阳性植株中,检测到GS3靶点突变植株125株,GS3靶点突变率为84%;对GS9基因进行编辑,得到100株阳性植株,其中检测到GS9靶点突变的植株共28株,GS9靶点的突变率为28%。在转入pYLCRISPR/Cas9-GS3-RNA载体的48株阳性植株中获得45株突变植株,突变率高达93.75%。在转入pYLCRISPR/Cas9-GS3-GS9-RNA载体的100株阳性植株中获得85株阳性突变植株,突变率为85%,其中东富139、龙粳31和东农427分别筛选出GS3单基因突变植株19,15,23株,GS9单基因突变植株2,2,1株,GS3、GS9双基因突变植株7,10,6株(表2)。
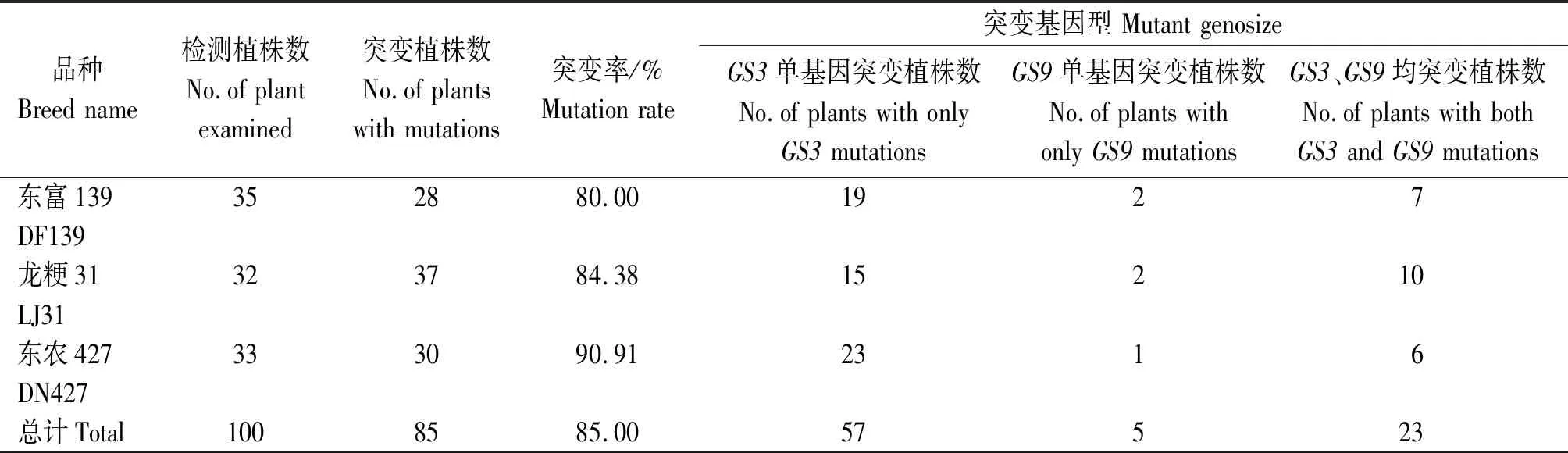
表2 转入pYLCRISPR/Cas9-GS3-GS9-RNA载体的T0植株测序Tab.2 Sequencing results of T0 plants transformed into pYLCRISPR/Cas9-GS3-GS9-RNA vector
2.5 T1纯合无T-DNA元件植株筛选
为获得东富139、龙粳31和东农427 3个品种的GS3单基因、GS9单基因和GS3、GS9双基因纯合无T-DNA元件突变植株,在T1利用引物GS3-seq-F/R、GS9-seq-F/R和Hyg-F/R进行PCR扩增并测序。最终获得东富139背景下GS3单基因、GS9单基因和GS3、GS9双基因纯合无T-DNA元件突变植株3,2,2株;龙粳31背景下GS3单基因、GS9单基因和GS3、GS9双基因纯合无T-DNA元件突变植株各2株;东农427背景下GS3单基因、GS9单基因和GS3、GS9双基因纯合无T-DNA元件突变植株4,2,1株。
在3个品种T1的gs3、gs9和gs3gs9纯合无T-DNA元件突变体中各选1株繁殖至T2,并进行农艺性状考察。详细突变情况见图3。

红色碱基为插入;红色虚线为碱基缺失;PAM序列用下划线及蓝色标注。Base insertion was marked with red letters;Base deletion was marked with red dotted line;PAM sequence was marked with underscore and blue.
2.6 T2植株农艺性状分析
将T2突变株系分别与其对应的野生型品种进行粒长、粒宽、千粒质量、结实率和穗粒数等农艺性状的比较分析(图4)。
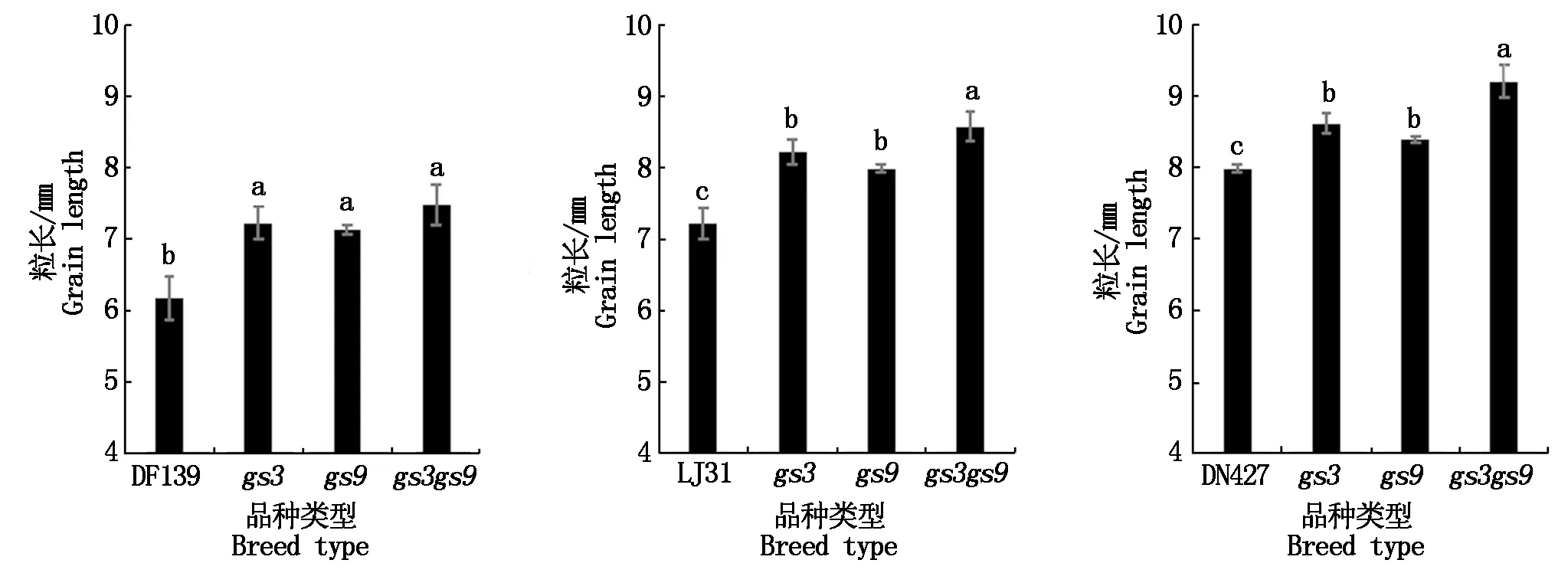
在3个粳稻品种中,gs3、gs9和gs3gs9突变体的粒长与野生型相比均显著增加(图5)。其中gs3突变体粒长较野生型增加了8%~17%,gs9突变体粒长较野生型增加了5%~15%,gs3gs9突变体粒长较野生型增加15%~21%,且每种突变体的粒长与其野生型相比都达到了显著水平。东富139突变体的粒长较野生型增加16%~21%,龙粳31突变体的粒长较野生型增加11%~19%,东农427突变体的粒长较野生型增加5%~15%,其中东富139的gs3gs9突变体粒长增加幅度最大,为21%;而东农427的gs9突变体增加幅度最小,为5%。GS3、GS9单基因突变都能显著增加水稻粒长,而GS3、GS9双基因突变对水稻粒长的影响,表现为2个基因的加性效应。
在粒宽方面,同一品种中,gs3突变体的粒宽与野生型相比,都没有显著变化;而gs9和gs3gs9突变体的粒宽都减少,且达到显著水平(P<0.05),其中gs9突变体的粒宽较野生型相比减少了7%~13%,gs3gs9突变体粒宽较野生型相比减少了8%~12%(图6)。不同品种的gs9、gs3gs9突变体的粒宽较野生型粒宽显著减少。说明GS3单基因突变对粒宽没有显著影响,而GS9单基因突变可显著减少水稻粒宽,在gs3gs9突变体中,粒宽减少主要是GS9的基因突变造成的影响。

图4 3个粳稻品种野生型及突变体粒型比较Fig.4 Comparison of grain size of wild type and mutant in three japonica rice varieties
不同小写字母表示在0.05水平差异显著。图6—9同。Different small letters show significantly different at 0.05.The same as Fig.6—9.

图6 3个粳稻品种野生型及突变体粒宽比较Fig.6 Comparison of grain width of wild type and mutant in three japonica rice varieties
在千粒质量方面,3个品种的gs9突变体千粒质量与野生型相比,没有显著差异;而gs3和gs3gs9突变体的千粒质量与野生型相比都显著增加,且增加幅度相近。其中gs3突变体千粒质量较野生型增加8%~15%,gs3gs9突变体较野生型增加10%~13%(图7)。这一结果表明,GS9基因突变对千粒质量性状无显著影响,GS3基因突变可增加水稻千粒质量,gs3gs9突变体千粒质量增加,主要是GS3单基因的效应。在结实率方面,3个品种突变体的结实率与野生型植株相比均没有显著差异(图8)。在穗粒数方面,3个品种的突变体中的穗粒数都略微减少,但都未达到显著水平(图9)。

图7 3个粳稻品种野生型及突变体千粒质量比较Fig.7 Comparison of 1000-grain weight of wild type and mutant in three japonica rice varieties

图8 3个粳稻品种野生型及突变体结实率比较Fig.8 Comparison of seed-setting rate of wild type and mutant in three japonica rice varieties
图9 3个粳稻品种野生型及突变体穗粒数比较Fig.9 Comparison of grain number per panicle of wild type and mutant in three japonica rice varieties
3 结论与讨论
将CRISPR/Cas9技术应用于作物种质资源的改良,不仅具备便捷、高效等优点,还能缩短育种年限。因此,这一技术正被广泛应用于水稻育种中[41-45]。本研究选择圆粒型粳稻品种东富139、龙粳31和中长粒型粳稻品种东农427为受体材料,利用CRISPR/Cas9基因编辑技术对GS3、GS9基因进行编辑,在3个品种中都得到了GS3单基因纯合突变、GS9单基因纯合突变和GS3、GS9双基因纯合突变的突变体材料。
本研究构建了pYLCRISPR/Cas9-GS3-RNA载体和pYLCRISPR/Cas9-GS3-GS9-RNA载体,用分别转入这2个载体的农杆菌侵染3个品种的水稻愈伤组织,成苗后筛选阳性植株,其中,转入pYLCRISPR/Cas9-GS3-RNA载体的48株阳性植株中,有45株为突变植株,突变率高达93.75%;转入pYLCRISPR/Cas9-GS3-GS9-RNA载体的100株阳性植株中,有85株为突变植株,突变率为85%,并且这85株突变植株中,有80株在GS3位点都发生突变。在之前的研究中,Chen等[46]利用CRISPR/Cas9技术构建GS3、GL3.1双敲除载体对水稻品种日本晴进行基因编辑,得到的所有阳性突变植株均在GS3靶点发生纯合突变。Shen等[47]研究结果表明,GS3靶点的诱变效率极高,本试验结果也证实了这一结论。总体上看,试验中所有阳性植株中GS3靶点的突变率为84%,该频率高于前人的研究结果;而对GS9基因进行编辑的受体材料中,GS9靶点的突变率为28%,显著低于前人的研究结果。有研究表明,靶序列的CG含量可能影响突变效率[38]。因此,推测不同靶序列中CG含量的差异,可能是造成基因突变率有显著差异的因素之一。
GS3是水稻粒型的主效QTL。沈兰等[48]研究结果表明,在4个粳稻品种中,gs3突变体的粒长较野生型增加了8%~18%,千粒质量较野生型增加2%~21%,结实率较野生型略微下降,但未达到显著水平,而粒宽和穗粒数较野生型没有显著差异。本试验中,3个粳稻品种的gs3突变体较野生型的粒长增幅(8%~17%)、千粒质量增幅(8%~15%)均与前人试验结果相近,结实率、粒宽和穗粒数的分析结果,也与前人试验结果基本一致。以上结果表明,GS3单基因突变可显著增加水稻粒长和千粒质量,并且在粒宽、结实率和穗粒数等性状较野生型没有显著差异,通过对GS3进行基因编辑,理论上可以增加水稻粒长,并通过增加千粒质量提高增产潜力。
GS9负调控水稻粒长,但对水稻粒宽有正调控作用[20]。张晨[49]研究结果表明,NIL(gs9)近等基因系的粒长较野生型增加9%,粒宽减少8%,千粒质量、株高、穗长、分蘖数等农艺性状较野生型没有显著差异。本试验中,3个品种的gs9突变体粒长、粒宽的变化幅度,与前人NIL(gs9)近等基因系的粒长、粒宽较野生型的变化幅度大致相近。前人试验结果表明,构建近等基因系也是精确改良水稻性状的有效方法。而本试验利用CRISPR/Cas9技术得到3个品种的gs9突变株系,相比前人构建近等基因系的方法,更加省时省力,并且对3个品种的突变株系进行农艺性状分析的结果,理论上来说更具说服力。综合本试验与前人试验结果,发现GS9作为一个控制粒长粒宽的主效QTL,可以在不影响其他农艺性状的基础上通过增加粒长并减少粒宽改良水稻粒型。
粒型是一个相对复杂的性状,受多个QTL影响,而GS3、GS9都是控制粒型的主效QTL,理论上结合这2个主效QTL,可以在水稻粒型的改良中,达到更理想的效果。徐善斌等[50]研究结果表明,通过对GS3和GS9进行基因编辑,粒长比野生型增加26.43%~27.01%,增幅大于GS3、GS9单基因突变体,粒宽降低13.4%~24.6%,千粒质量增加18.34%~41.36%,并且增产10.82%~12.11%。本试验中,3个品种的gs3gs9突变体的粒长、粒宽和千粒质量较野生型的变化幅度均低于前人研究结果,但粒长和千粒质量增幅明显大于GS3和GS9单基因突变体,与前人结果一致。以上结果表明,相较于单独敲除GS3或GS9基因,同时对GS3和GS9这2个主效QTL进行基因编辑,可以在改良水稻粒型中取得更好的效果,并且在一定程度上提高水稻的增产潜力。
综上所述,本试验以3个粳稻品种东富139、龙粳31和东农427为受体材料,利用CRISPR/Cas9技术对GS3、GS9基因进行定向敲除,3个品种均获得了gs3、gs9和gs3gs9且无T-DNA元件的纯合突变体,创制了多个粒型改良的粳稻新种质,加快了长粒型粳稻新品种的选育进程。
