人参UGT基因家族功能研究进展
2022-04-15王海娇李宇兴佟爱仔胡勇军
王海娇,李宇兴,佟爱仔,刘 伟,胡勇军*
(1.长春师范大学,长春 130032;2.通化师范学院,通化 134002)
人参(Panax ginseng)是五加科人参属的多年生早本植物,是一种被广泛应用的传统中药材,其主要药理活性成分是人参体内的三萜类化合物人参皂苷[1-3]。目前已经证实人参皂苷的生物合成分为上、下游合成途径,且上游合成途径已经确定,包括MVA途径和MEP途径,其中以MVA途径为主[2,4,5]。下游合成途径一般包括环化、羟基化和糖基化过程[2,5-7]。糖基化是可产生天然化合物的植物体内存在的一种重要修饰反应,糖基转移酶是一类可以催化糖基化反应的酶家族,即将糖基从活化的糖供体分子转移到糖受体分子上[8-12]。在人参体内人参皂苷生物合成最后一步就是通过糖基化来完成的。
截止到2020年,糖基转移酶已被发现有110个家族,其中GT1包含最多[11,13],因此对GT1的研究最为广泛。在人参中,因其都以活化尿苷二磷酸糖类作为糖基供体,因此被称为UDP-糖基转移酶(UDPglycosyltransferase),即UGT[2,12]。
人参皂苷作为一种具有良好药理活性的化合物,搞清楚每种皂苷单体的合成机制,就可以使体外合成皂苷单体并提高皂苷单体合成量成为可能。作为催化人参皂苷合成最后一步糖基化反应的UGT,其具体催化机制大多尚不明确,因此对人参UGT的研究意义重大。本文结合前人的大量研究对糖基转移酶包括UGT及UGT的功能进行概述,同时介绍了人参UGT基因家族功能到目前为止的研究进展。可为后续对UGT基因家族的研究提供借鉴。
1 糖基转移酶的概述
通过阅读文献发现,在2003年时,被发现的糖基转移酶一共有65个家族[14],到2018年时,被发现的糖基转移酶共有99个家族[2],而截止到2020年,被发现的糖基转移酶已经有110个家族[13]。由此可知,人们对糖基转移酶的研究从未停止,正因其家族的庞大,研究过程也就变得十分困难。
1.1 糖基转移酶的分类
据研究表明,糖基转移酶有4种分类形式。根据催化反应机理可分为催化型和保留型;根据糖基转移酶蛋白质的立体空间结构不同可分为GT-A、GT-B、GT-C以及GT-D四种类型;根据糖基化位点可分为O-、N-、S-、C-等连接型,但大多是O-连接型;根据氨基酸的相似性来看,植物中糖基转移酶家族1的氨基酸序列大部分在C端有一个比较保守的、由44个氨基酸残基组成的PSPG盒,其功能可能是糖基转移酶与UDP-糖供体识别和结合的位点[2]。
1.2 糖基转移酶的生物学功能
目前,已知糖基转移酶的生物学功能主要包括:a.调节植物激素平衡;b.参与植物体内的次级代谢反应;c.参与植物的生长发育;d.参与植物的脱毒反应;e.参与植物体内的信号转导;f.参与植物的生物与非生物的胁迫反应等[2,9,15]。
UGT作为主要以UDP-糖为糖供体的糖基转移酶同样也具备上述生物学功能,但由于所催化化合物种类的复杂性及每种UGT对糖基供体分子和糖基受体分子严格专一性,导致不同植物中不同种类UGT的催化作用机制不同。有研究表明,有的UGT在以一种糖基供体为主的同时,能利用多种糖基供体分子,有的UGT也能催化形成多种糖基受体分子,这可能与受体分子结构有关[2,8]。本文通过查阅文献总结了以下几种植物中UGT的具体功能。
在模式生物拟南芥中一共发现138个UGT基因,其中107个具有编码酶功能[7],拟南芥UGT73C4基因参与拟南芥的抗病作用[9],UGT76F1基因通过IPyA糖基化修饰参与光和温度介导的植物下胚轴的生长[13]。
蒺藜苜蓿中发现UGT71G1可参与苜蓿酸常春藤皂苷元的糖基化修饰,UGT73K1可参与常春藤皂苷元和大豆皂醇B、E的糖基化修饰,UGT73F3可参与常春藤皂苷元和其它齐墩果烷型皂苷元C-28位羧基的糖基化修饰[16]。
王不留行中发现UGT74M1可参与齐墩果烷型皂苷元的C-28位糖基化修饰[16]。
大豆中发现UGT73P2可催化半乳糖基连接到葡糖醛酸化的大豆皂醇B上,UGT91H4可催化鼠李糖基连接到二糖基化的大豆皂醇Ⅲ的半乳糖基上,UGT73F4可催化木糖基连接到大豆A组皂苷C-22位的阿拉伯糖残基上,UGT73F2可催化葡萄糖基连接到大豆A组皂苷C-22位的阿拉伯糖残基上[16]。
2 人参UGT基因家族研究进展
2.1 人参皂苷合成途径研究
人参皂苷与其他三萜皂苷的生物合成方式类似,首先经过胞质中的甲羟戊酸途径(MVA)和位于质体的2-C-甲基-D-赤藓糖醇-4-磷酸途径(MEP)形成萜类前体异戊烯二磷酸(IPP)及其异构体二甲基烯丙基二磷酸(DMAPP)。IPP和DMAPP缩合生成牻牛儿基焦磷酸(GPP),随后与IPP生成法尼基焦磷酸(FPP)。2个FPP分子在鲨烯合酶(SQS)作用下产生内环化反应生成鲨烯,而鲨烯是三萜类化合物和相关衍生物的重要前体物质[17]。随后,鲨烯经鲨烯环氧酶(SQE)[18]的氧化反应,达玛烯二醇合酶(DS)[19]的环化反应和细胞色素P450单加氧酶[20,21]的羟化反应,分别形成四环三萜皂苷的重要中间产物2,3-氧化鲨烯、达玛烯二醇(DM)、PPD及PPT[20-22]。PPD与PPT是形成人参皂苷的核心糖苷配基结构,在多种UGT糖基化作用下与葡萄糖、木糖、鼠李糖、阿拉伯糖基等单一或者混杂连接形成多种活性功能各异的人参皂苷[23]。
整个过程包括20多个连续酶促反应步骤,其中涉及一系列关键酶,如3-羟基-3-甲基戊二酰辅酶A还原酶(HMGR)、法尼基焦磷酸合成酶(FPS)、SQS、SQE、DS、原人参二醇合酶(PPDS)、(CYP716A47)、原人参三醇合酶(PPTS)、(CYP716A53)和UGT,具体过程见图1,图片来自[24]。
2.2 人参UGT基因功能研究现状
目前,通过454EST分析检测到人参中共含有235个UGT基因[25]。但其中大部分UGT基因的具体功能尚不明确,经查阅相关文献了解到目前已鉴定出一部分基因功能。如下:人参PgUGT71A27可以催化PPD型人参皂苷糖基化生成化合物K[26];人参PgUGT1具有较松弛的底物特异性,可修饰PPD型人参皂苷的合成,即催化人参皂苷Rg3的C-20位糖基化合成人参皂苷Rd[7,27];人参PgUGTPg100和人参PgUGTPg101均能特异性催化PPT型人参皂苷的生物合成[27],其中PgUGTPg100可同时催化PPT型人参皂苷的C-6位糖基化合成人参皂苷Rh1和人参皂苷F1到人参皂苷Rg1的生物合成,而PgUGTPg101可同时作用于PPT型人参皂苷的C-6和C-20位,产生人参皂苷F1,并通过顺序糖基化作用产生人参皂苷Rg1[7,26];人参PgUGT94Q2可催化葡萄糖部分从UDP-葡萄糖转移到人参皂苷Rh2和F2的糖上,分别合成人参皂苷Rg3和Rd[7,16,26-28];人参PgUGT74AE2可催化PPD型人参皂苷和化合物K C-3羟基位的糖基化,从而分别合成人参皂苷Rh2和F2[7,16,26-28];人参UGTPg71A29可以催化人参皂苷Rh1的C-20位的糖基化作用形成人参皂苷Rg1,同时也可以催化人参皂苷Rd糖基化形成人参皂苷Rb1[27]。这些在人参中发挥重要催化作用的UGT有一个共同特点,即在C-末端有一段由44个氨基酸残基组成的保守序列[7,12],因此推测UGT基因发挥功能的片段很有可能是这个保守序列。已知功能人参UGT及其相关信息具体见表1。
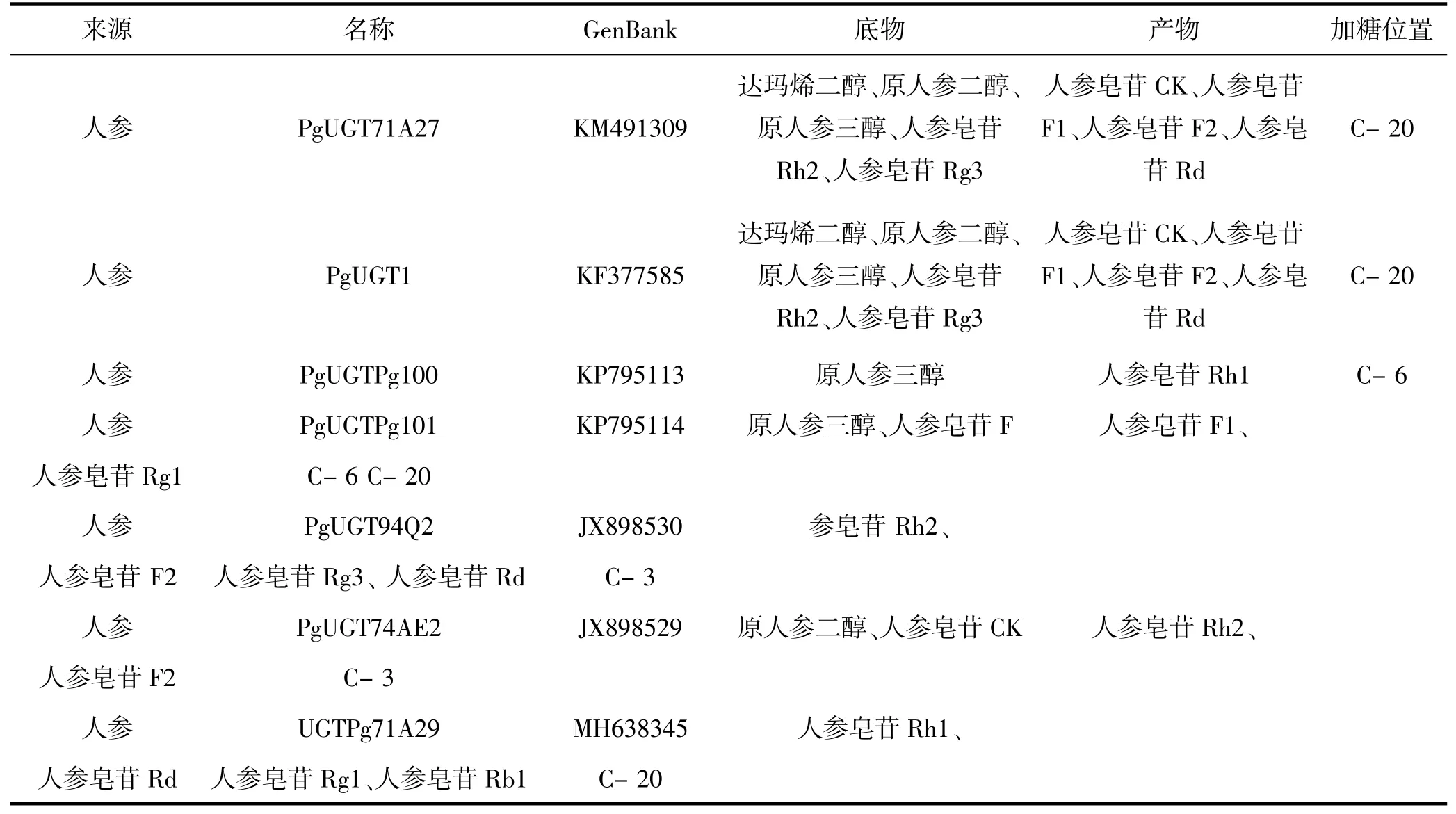
表1 人参中已验证功能的UGTTable 1 Ginseng to verify the function of UGT
综上,在人参中,UGT对人参皂苷生物合成的催化作用对于PPD型人参皂苷主要作用在C-3和C-20位,对于PPT型人参皂苷主要作用在C-6和C-20位。这一结论可帮助我们在后续对候选基因功能的研究提供参考和思路,可根据对候选基因同源序列比对来预测其可能具有的功能。
3 讨论与展望
糖基转移酶修饰的糖基化过程普遍存在于能够合成天然化合物的植物中,是植物中一种重要的修饰反应。对于人参来说,通过科研工作者多年的努力,人参皂苷生物合成途径的解析已基本完成,但对于人参皂苷苷元的糖基化修饰研究还处于初始阶段。而糖基转移酶作为一种多基因家族酶,其庞大的种类数量、特异的序列及专一的选择性都给我们研究人参皂苷苷元的糖基化修饰带来了挑战。不过,近年来,通过发展迅速的转录组测序技术及生物信息学分析,已经筛选出一些与人参皂苷合成相关的糖基转移酶基因序列,但完成功能鉴定的糖基转移酶数量却极其有限。因此,我们可以预见,未来人们对人参糖基转移酶基因及功能的研究将会不断深入且意义重大。这一研究成果可以使未来在人参体外合成各种具有生物活性的人参皂苷成为可能。但此项工作的艰难程度可想而知,这需要科研工作者们共同努力完成。
